鹽穗木HcALDH7B4基因的原核表達增強耐鹽抗旱性
2023-09-12 09:40:48毛曉菲黃世平銀芳柳曾幼玲
新疆農業科學 2023年8期
關鍵詞:植物
毛曉菲,黃世平,銀芳柳,曾幼玲
(新疆大學生命科學與技術學院/新疆生物資源基因工程重點實驗室,烏魯木齊 830000)
0 引 言
【研究意義】干旱和高鹽脅迫是抑制植物生長和作物產量的兩個主要環境因素。當植物遭受鹽旱脅迫時會導致細胞中活性氧(ROS)的過度累積[1],ROS會氧化膜磷脂以及游離氨基酸等物質而產生有毒物質,如醛類[2]。解決植物在逆境環境下的生存問題,有必要去除和解毒過量的醛類化合物[3]。【前人研究進展】乙醛脫氫酶基因(ALDH)超家族成員是一大類催化酶,在NAD(P)+的協助下,乙醛脫氫酶可以將有毒醛氧化形成相對應的羧酸,使植物不被醛類毒害而緩解逆境脅迫[4]。基于其蛋白序列,共鑒定出24個家族,其中ALDH10、ALDH12、ALDH19、ALDH21、ALDH22、ALDH23和ALDH24家族是植物所特有的[5]。ALDH7家族的酶被稱為Δ1-piperideine-6-carboxylate脫氫酶(P6CDH, E.C.1.2.1.31),或α-氨基己二酸半醛脫氫酶(AASADH),是一個典型的跨物種高度保守的蛋白群[5],參與賴氨酸的分解代謝,并催化α-氨基己二酸半醛轉化為無毒的α-氨基己二酸,從而緩解醛類對植物的毒害[6]。植物ALDH基因已經從多種植物中克隆,如擬南芥(Arabidopsisthaliana)[7]、水稻(OryzasativaL.)[8]、葡萄(VitisviniferaL.)[9]、玉米(ZeamaysL.)[10]等,且基因的過表達在不同程度上可提高轉基因植物對鹽旱脅迫和氧化脅迫的耐受性。【本研究切入點】鹽穗木(Halostachyscaspica, Bieb.)是一種耐鹽性極強的莧科矮灌木,自然生長在高鹽堿半荒漠地區。課題組前期以pMD18-T Simple-HcALDH7B4為模板,通過PCR擴增、連接轉化以及酶切,成功構建重組的原核表達載體pET28a-HcALDH7B4[11]。需繼續基于已構建的pET-28a-HcALDH7B4重組載體,原核誘導表達,分析鹽旱脅迫下重組大腸桿菌耐鹽抗旱性。【擬解決的關鍵問題】研究HcALDH7B4基因在原核細胞里的耐鹽抗旱性,分析鹽旱脅迫下重組菌株的菌落大小和生長活力,為HcALDH7B4基因的功能提供依據。
1 材料與方法
1.1 材 料
將實驗保存的鹽穗木種子接種到培養基質(1∶1∶2蛭石∶珍珠巖∶土壤)中,培養條件:溫度26℃,相對濕度20%~30%,黑暗/光照周期為8/16 h。生長2.5個月后給予鹽旱脅迫處理。設鹽穗木不同濃度的NaCl處理(200、400、600 mmol/L)并分別于0、6、12、24、48、72 h采樣;于旱脅迫,設自然干旱處理并分別于15、30 d采樣。
E.coliDH5α、Transetta (DE3)購自北京全式金生物技術有限公司;植物總RNA提取試劑盒購自北京天根生化科技有限公司;Transcriptase M-MLV(RNase H)、ITPG、蛋白Marker購自大連寶生物技術有限公司;SYBR?Select master Mix購自賽默飛世爾科技(中國)有限公司;引物由上海生工工程股份有限公司合成;其余常用試劑為國產分析純試劑。
1.2 方 法
1.2.1 HcALDH7B4的生物信息學分析
ExPASy 軟件用于在線預測鹽穗木HcALDH7B4蛋白質的基本理化性質、親疏水性和三級結構;Conserved Domain Search(https://www.ncbi.nlm.nih.gov/structure/cdd/wrpsb.cgi)用于其結構域分析。通過NCBI中的BLAST(https://blast.ncbi.nlm.nih.gov)檢索HcALDH7B4同源序列,利用MEGA 7.0軟件中ClustalW程序進行多序列比對,通過Neighbor-joining鄰接法構建系統進化樹。
1.2.2 RNA的提取和反轉錄
取1.5 g鹽穗木同化枝,提取總RNA。使用M-MLV(Takara)反轉錄成cDNA。
1.2.3HcALDH7B4在不同處理下的表達模式
基于鹽穗木HcALDH7B4的序列,設計特異性引物HcALDH7B4-p1和HcALDH7B4-p2 。使用SYBR?Select master Mix和ABI 7500實時熒光定量PCR儀進行檢測。反應條件:94℃ 5 min,94℃ 30 s,60℃ 30 s,40個循環。每個樣本有4個生物學重復。β-actin作為內參基因。相對表達水平用2-ΔΔct定量。相對定量的最終值為基因表達與試驗中對照相比的倍數變化。表1
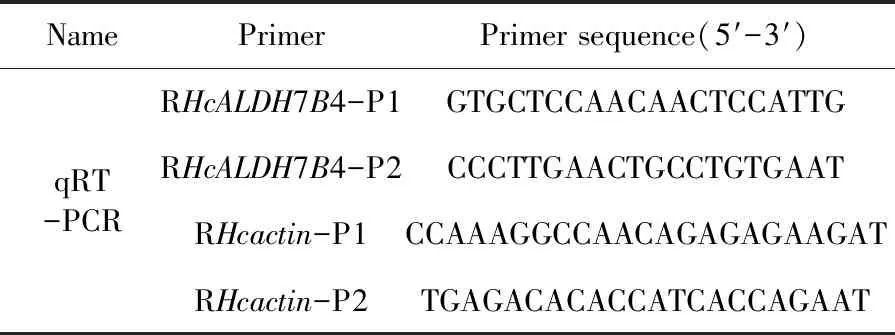
表1 引物
1.2.4 原核表達的重組菌(pET-28a-HcALDH7B4)的鑒定和誘導表達
通過PCR擴增以及雙酶切對前期構建的pET28a-HcALDH7B4重組載體進行鑒定。將鑒定正確的含有重組質粒pET28a-HcALDH7B4的Transetta菌種擴大培養至菌液OD600在0.6~0.8時,加入ITPG至終濃度0.6 mmol/L,30℃誘導表達4h,SDS-PAGE檢測目的蛋白的表達并用Western blot鑒定[11]。
1.2.5 重組菌株的鹽旱脅迫耐受性測定
滴板實驗:將重組菌株(pET-28a-HcALDH7B4)和對照菌株(pET-28a)擴大培養至OD6000.5后加入0.6 mmol/L ITPG,誘導培養4 h至OD6000.8,之后以10-1、10-2、10-3和10-4為梯度將菌液進行稀釋[12]。將每個稀釋的樣品(5 μL)分別點于含有500 mmol/L NaCl、500 mmol/L KCl、500、800 mmol/L Manntiol的固體LB培養基上培養12 h后拍照。
液體培養:將達到OD6000.8的2種菌株按1%(V/V)的接種量,分別接種到添加有500 mmol/L NaCl、500 mmol/L KCl、500、800 mmol/L Manntiol的新鮮LB液體培養基中,37℃、220 r/min振蕩培養。每隔2 h測菌液在不同處理下OD600的值。
1.3 數據處理
利用GraphPad Prism軟件繪圖和數據分析。用Duncan法進行差異顯著性檢驗。不同小寫字母代表組間有顯著差異(P< 0.05)。表1
2 結果與分析
2.1 HcALDH7B4的生物信息學
研究表明,鹽穗木HcALDH7B4蛋白的理論等電點為5.91,理論分子量為55kD。該蛋白多肽鏈中疏水性氨基酸多于親水性氨基酸,HcALDH7B4屬于疏水性蛋白。HcALDH7B4蛋白具有ALDH家族典型的NAD結合位點、四聚體界面、催化殘基,HcALDH7B4是ALDH基因超家族的一員。圖1
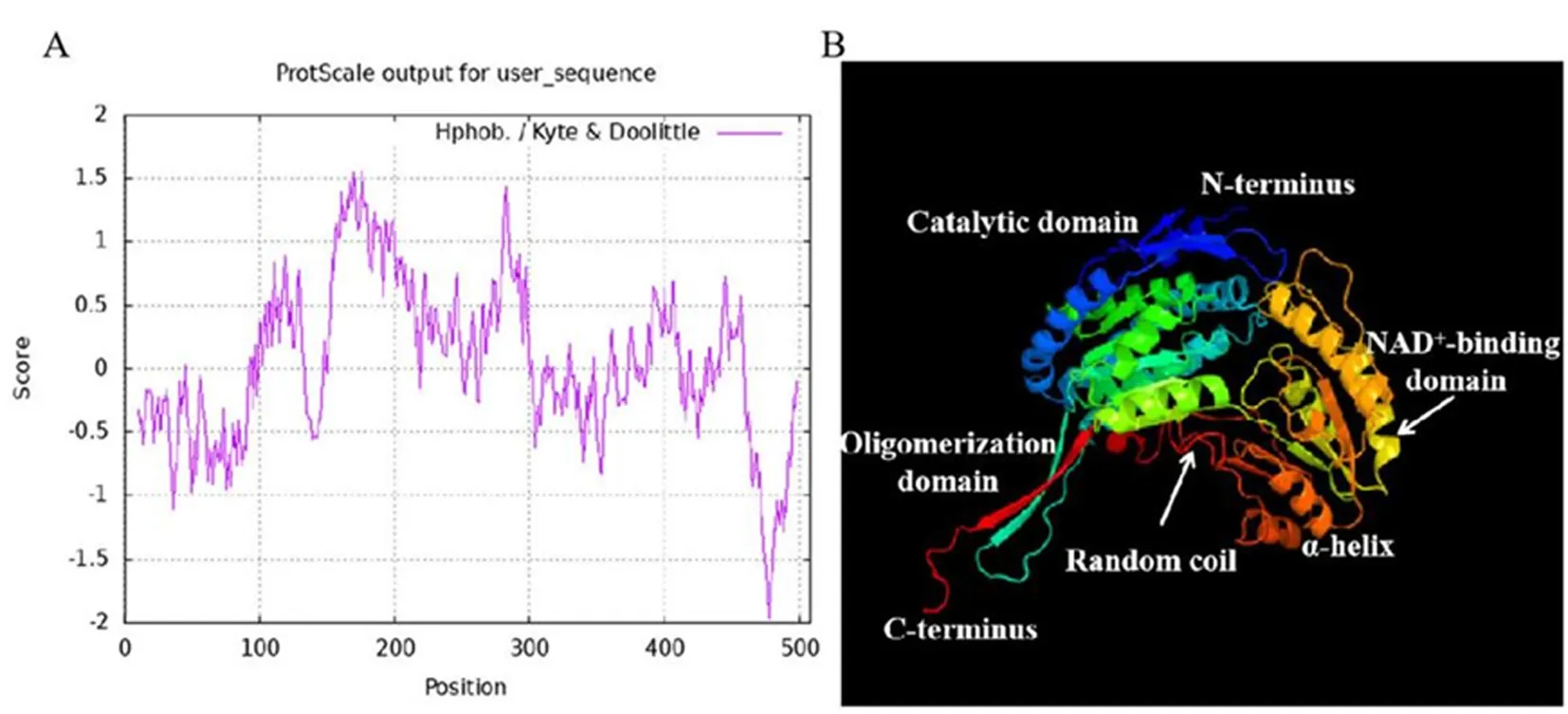
注:A、B分別為HcALDH7B4蛋白的疏水性分析和三級結構預測
HcALDH7B4與擬南芥AtALDH7B4在同一分支且有較高的氨基酸序列相似性,HcALDH7B4與擬南芥AtALDH7B4在耐鹽抗旱方面有相似的功能。圖2
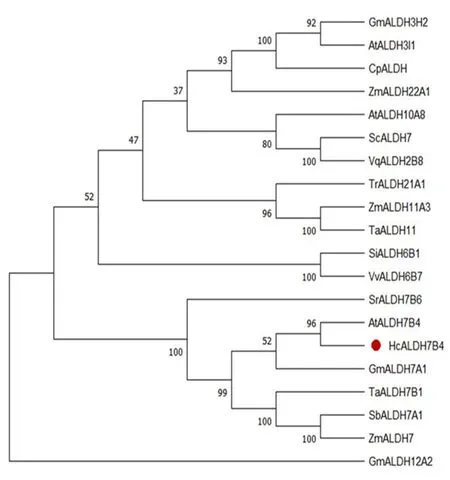
注:HcALDH7B4由紅色顯示
2.2 鹽旱脅迫下鹽穗木HcALDH7B4基因的表達模式
研究表明,HcALDH7B4基因在鹽旱脅迫后被顯著誘導。600 mmol/L NaCl處理48 h時HcALDH7B4的表達量最高。自然干旱脅迫下,隨著脅迫時間的延長,HcALDH7B4基因的表達逐漸增加,30 d時表達量最為顯著。圖3
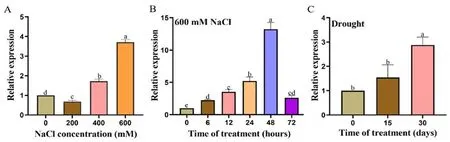
注:不同濃度的NaCl脅迫(A);600 mmol/L NaCl脅迫(B);不同時期的干旱脅迫(C)
2.3 融合蛋白的鑒定與誘導表達
研究表明,將鑒定正確的原核表達載體pET28a和重組質粒pET28a-HcALDH7B4分別轉化Transetta(DE3)后,經IPTG誘導表達,12%SDS-PAGE電泳檢測,在44~66 kDa有一條差異表達條帶,構建原核表達載體正確。在55~70 kDa有一條明顯的特異性條帶,HcALDH7B4蛋白被準確表達。取超聲處理后菌體上清及沉淀進行蛋白可溶性檢測。包涵體是目的蛋白存在的主要形式。圖4
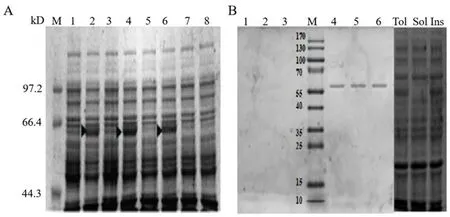
注: A: SDS-PAGE鑒定。M: 蛋白分子量Marker;1,3,5: 重組菌誘導前;2,4,6: 重組菌誘導后;7: 對照菌誘導前;8: 對照菌誘導后。B: Western blot分析。1,2,3: 重組菌誘導前;4,5,6: 重組菌誘導后。Total: 細胞總粗蛋白;Ins: 不溶性蛋白質;Sol: 可溶性蛋白
2.4 重組菌株的鹽旱耐受性
研究表明,重組菌與對照菌在菌落數方面沒有明顯差異;相較對照菌,重組菌株在500 mmol/L NaCl、500 mmol/L KCl、500、800mmol/L Mannitol脅迫下有更多的菌落數。正常情況下,重組菌和對照菌生長沒有差異,20 h后基本達到平穩期,脅迫條件下,兩者菌體生長量均有顯著差異,并且對照菌的生長受到明顯脅迫抑制。HcALDH7B4的表達可正向調節大腸桿菌在鹽旱脅迫下的生長。圖5,圖6
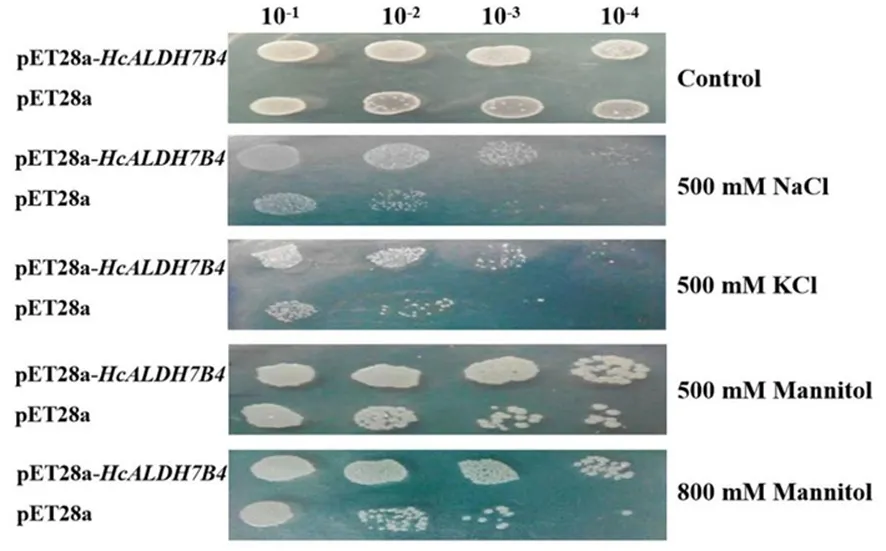
圖5 重組大腸桿菌菌株的鹽旱抗性
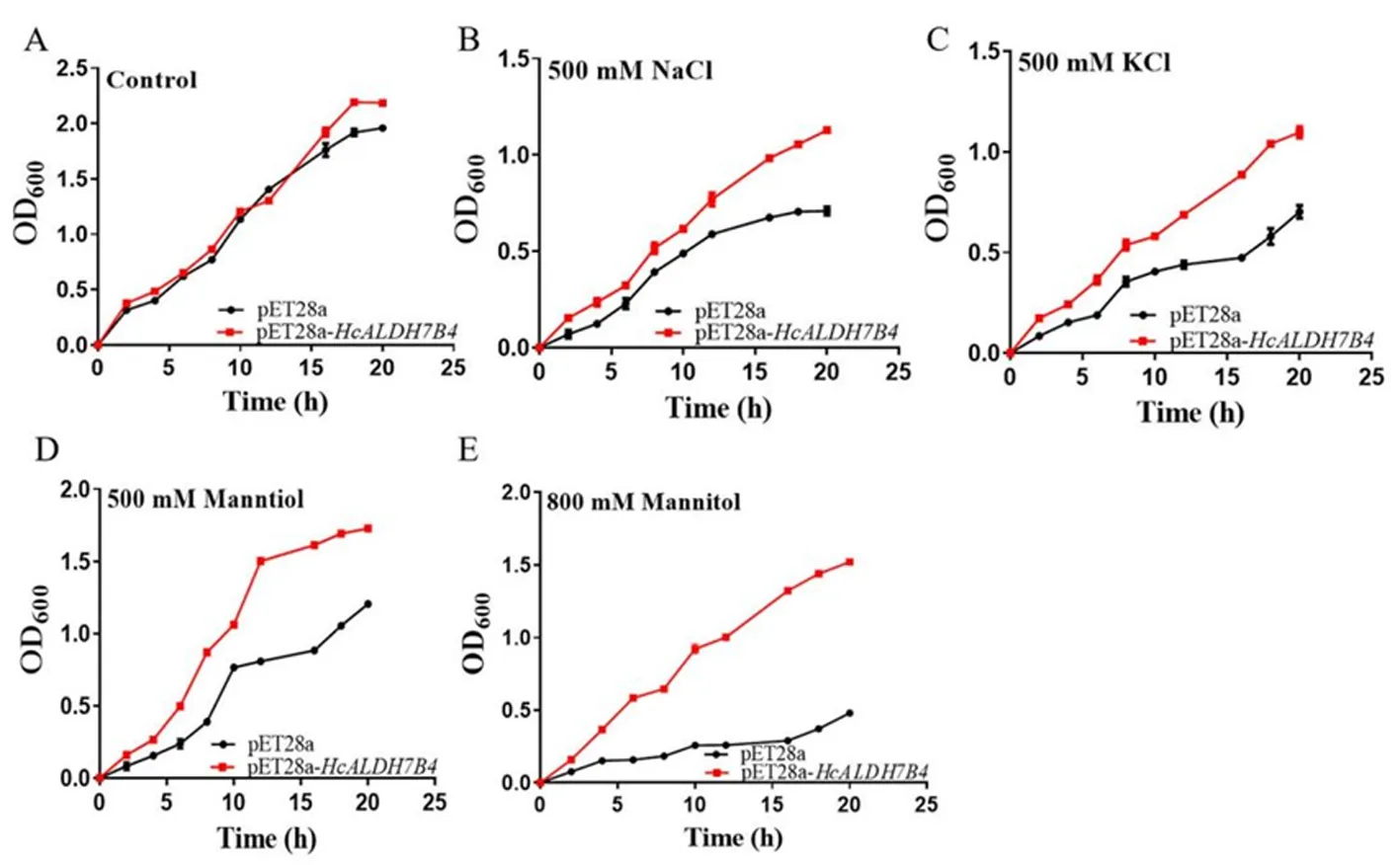
注:(A) 對照;(B) 500 mmol/L NaCl脅迫;(C) 500 mmol/L KCl脅迫;(D) 500 mmol/L Mannitol脅迫;(E) 800 mmmol/L Mannitol脅迫
3 討 論
3.1醛脫氫酶(ALDHs)是一個進化保守的氧化還原酶超家族,將大量的醛轉化為羧酸來緩解毒害。前期對ALDH家族在非生物脅迫上的功能研究主要集中在模式植物擬南芥[13]、玉米[14,15]、水稻[16]中,而對鹽生植物中該蛋白家族的研究較少。
3.2前期研究證實ALDH7家族基因在各種非生物脅迫下被顯著誘導,并參與植物對非生物脅迫的響應。在鹽、旱以及ABA等其他非生物脅迫下,大豆GmALDH7B1[17]、水稻OsALDH7[18]和高粱SbALDH7B2[19]基因的表達顯著上調,ALDH7基因參與了植物對多種非生物脅迫的響應。與HcALDH7B4聚為一類的擬南芥AtALDH7B4在脫水、高鹽、H2O2或ABA處理后在葉和根中也被強烈誘導[20],且在轉基因植物中可提高其鹽旱耐性。試驗中HcALDH7B4在鹽旱脅迫下也被顯著誘導。HcALDH7B4可能有助于植物響應鹽旱脅迫。
3.3研究結果表明,鹽穗木HcALDH7B4基因的原核表達增強了大腸桿菌的鹽旱耐性。與Gautam等[21]的結果相一致,油菜BrALDH7B2的過表達也使大腸桿菌和酵母在鹽脅迫和氧化脅迫下有更好的適應和生存能力,且過表達BrALDH7B2可提高轉基因煙草的耐鹽性[22]。鹽穗木HcALDH7B4很有可能也在植物的非生物脅迫中發揮正向的調控作用。HcALDH7B4是一個脅迫應答基因,其原核表達可以提高大腸桿菌鹽旱耐性。進一步通過植物轉基因方式提高作物的非生物抗性后,有望作為候選基因提高作物和林木的耐鹽和抗旱性。
4 結 論
根據鹽穗木HcALDH7B4基因在600 mmol/L NaCl和自然干旱脅迫下不同時間點的表達量,鹽穗木HcALDH7B4基因在鹽旱脅迫下被顯著誘導;相較對照菌,500 mmol/L NaCl、500 mmol/L KCl、500、800 mmol/L Manntiol脅迫下重組菌的菌落大小和生長活力更優。原核表達HcALDH7B4能夠提高大腸桿菌的鹽旱抗性。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
