miR-145-5p靶向FGF5對神經細胞缺血/再灌注損傷誘導的神經元細胞凋亡和氧化應激的影響
2023-09-22 03:21:02賈燕燕張小林
中西醫結合心腦血管病雜志 2023年17期
關鍵詞:氧化應激
賈燕燕,張小林,潘 燕,趙 迪
缺血性腦血管病(ischemic cerebrovascular diseases,ICVD)是具有高發病率、高致殘率及高死亡率的急性腦血管疾病[1-2],其中缺血性腦卒中的發病率占所有腦卒中的87%,臨床通常采用及時溶栓、介入等方式恢復缺血區域的血流再灌注,但血流再灌注可能引發缺血性再灌注損傷(ischemia/reperfusion injury,I/R)。神經元細胞是對缺血缺氧最敏感的細胞,I/R對神經元細胞的損傷非常大。研究表明,缺血性腦卒中的病理生理機制包括興奮神經毒性神經遞質釋放、細胞內Ca2+積聚、自由基損傷、細胞凋亡、神經炎癥及脂質分解等[3],但I/R的發病機制仍不完全清楚,因此,進一步研究缺血性腦卒中的發病機制以尋找潛在的治療靶點,從而提高缺血性腦卒中療效,降低病人死亡率和致殘率。miRNA是真核生物中具有調控功能的短鏈非編碼RNA[4],包含21~23個核苷酸序列[5],可與目標基因mRNA的3′-非編碼區(3-UTR)堿基配對結合,對目標基因發揮調控作用。近年來的研究表明,miRNA對缺血再灌注損傷具有調控作用,如miR-429的表達可緩解氧糖剝奪/復氧(OGD/R)對神經細胞的損傷[6],而microRNA-320可通過抑制胰島素生長因子-1調控腦缺血/再灌注損傷[7];miR-145-5p受長鏈非編碼RNA SNHG14調控,參與腦梗死的發生發展[8]。最近的研究表明,缺血時miR-145-5p表達水平的改變可影響神經元的活性[9]。成纖維細胞生長因子5(FGF5)是小鼠擬胚體外胚層標志物,在胚胎發育和神經發育中發揮重要作用。本實驗以體外培養的小鼠海馬區神經元細胞為研究對象,構建OGD/R模型,研究miR-145-5p與FGF5的靶向關系及其對氧化應激反應的影響,為腦缺血再灌注提供潛在靶點。
1 材料與方法
1.1 實驗材料
小鼠海馬神經元HT22細胞購自中國科學院上海細胞庫;miR-145-5p inhibitor由廣州銳博生物有限公司合成;小干擾RNA(siRNA)靶向FGF5購自上海Genechem公司;胎牛血清、DMEM培養基和胰蛋白酶購自美國Gibco公司;活性氧(ROS)檢測試劑盒、放射免疫沉淀試驗(RIPA)裂解液、二喹啉甲酸(BCA)試劑盒均購自上海碧云天生物技術研究所;丙二醛(MDA)試劑盒、超氧化物歧化酶(SOD)試劑盒、乳酸脫氫酶(lactate dehydrogenase,LDH)試劑盒均購自北京康為世紀生物科技有限公司。
1.2 細胞培養與OGD/R模型建立
細胞培養:采用含10%胎牛血清、1%青霉素/鏈霉素的DMEM培養基培養HT22細胞,培養條件為37 ℃、5%CO2。OGD/R模型建立:HT22細胞置于無糖平衡鹽溶液中,在三氣缺氧細胞培養箱(37 ℃、1%O2、94%N2、5%CO2)中培養4 h后,更換為含4.5 g/L葡萄糖的培養基,常規復氧培養(95%空氣、5%CO2)12、24 h制備OGD/R模型,對照組不進行OGD/R處理,常規條件下培養。
1.3 流式細胞術檢測神經元細胞凋亡
將HT22細胞(密度為2×105個/mL)接種于六孔板,按上述OGD/R模型建立方法處理HT22細胞6、12、24 h,收集細胞并離心,用預冷磷酸緩沖鹽溶液(PBS)洗滌兩次,調整細胞密度為1×106個/mL。將5 μL的V-異硫氰酸熒光素(Annexin V-FITC)和10 μL的碘化丙啶(PI)加入100 μL的1×Binding Buffer重懸細胞懸液中混勻,室溫避光孵育5 min,最后加入400 μL的1×Binding Buffer混勻,在流式細胞儀上進行檢測,并采用BD FAC-Suite軟件作圖。
1.4 蛋白免疫印跡法(Western Blot)檢測蛋白表達
復氧24 h后收集各組細胞,加入RIPA蛋白裂解液冰上裂解30 min,在低溫冷凍離心機上以12 500 r/min離心10 min,取上清,采用BCA蛋白定量試劑盒測定蛋白總濃度。取50 μg總蛋白進行聚丙烯酰胺凝膠(SDS-PAGE)電泳,電轉至聚偏二氟乙烯膜(PVDF),Tris 緩沖鹽溶液(TBS)洗膜10 min,5%牛血清白蛋白封閉2 h,加一抗4 ℃孵育過夜,次日加入相應二抗孵育1~2 h,增強型化學發光劑(ECL)顯色,采用Image J2x圖像分析系統分析蛋白條帶灰度值,以甘油醛-3-磷酸脫氫酶(GAPDH)為內參。
1.5 氧化應激標志物檢測
分別采用MDA、SOD、ROS及LDH試劑盒檢測細胞MDA、SOD、ROS及LDH含量。
1.6 熒光素酶報告實驗驗證靶向關系
采用Targetscan 和MiRDB生物信息學預測軟件預測miR-145-5p和FGF5的結合位點,合成攜帶miR-145-5p結合位點的野生型(WT)FGF5 3′-UTR,并將其克隆至pMIR-control熒光素酶報告質粒中,293T細胞接種到95孔板中,使用Lipofectamine3000將pcDNA3.0-FGF5 WT和突變型(MUT)質粒分別與miR-145-5p mimic共轉染,48 h后,通過雙熒光素酶報告試驗系統測定熒光素酶活性。
1.7 細胞轉染
將siRNA靶向FGF5和對照siRNA分別插入pcDNA3.0質粒,取對數生長期的HT22細胞,調整細胞密度為1×104個/mL,接種于24孔板,常規培養24 h后,采用Lipofectamine 3000將上述pcDNA3.0質粒轉染至HT22細胞。
1.8 實時定量聚合酶鏈式反應(RT-PCR)
采用TRIzol試劑提取細胞總RNA,然后將RNA逆轉錄為cDNA,然后以cDNA為模板采用SYBR Gene RealtimePCRMaster Mix進行RT-PCR,使用Fast Real-time PCR 7300系統分析miR-145-5p的mRNA水平,反應均以U6為內參計算蛋白的相對表達量,采用2-ΔΔCt計算mRNA相對表達量。詳見表1。

表1 基因名稱及引物序列
1.9 統計學處理
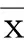
2 結 果
2.1 OGD/R處理對神經細胞氧化應激反應的影響
神經細胞經氧糖剝奪后復氧,細胞ROS、MDA及細胞上清LDH含量隨復氧時間的增加而增加,而SOD活性則隨復氧時間的增加而降低,OGD/R 6、12、24 h時ROS、MDA、LDH、SOD水平與0 h比較,差異均有統計學意義(P<0.01),提示OGD/R誘導神經細胞出現氧化應激反應。詳見圖1、圖2。

圖1 OGD/R處理對神經細胞LDH、SOD水平的影響

圖2 OGD/R處理對神經細胞ROS、MDA水平的影響
2.2 OGD/R處理對miR-145-5p表達的影響
神經細胞經氧糖剝奪后復氧不同時間,miR-145-5p表達均高于對照組(P<0.01),并且隨著復氧時間的增加,miR-145-5p表達量增加,提示OGD/R能誘導神經細胞中miR-145-5p的表達,詳見圖3。神經細胞轉染miR-145-5p mimic后氧糖剝奪,miR-145-5p表達水平明顯高于OGD/R 24 h;而miR-145-5p inhibitor組miR-145-5p表達水平明顯低于OGD/R 24 h組,差異均有統計學意義(P<0.01),詳見圖4。提示miR-145-5p的高表達與I/R損傷相關。

圖3 神經細胞經OGD/R不同時間miR-145-5p表達比較

圖4 神經細胞轉染miR-145-5p mimic后氧糖剝奪對miR-145-5p表達的影響
2.3 OGD/R處理對神經元細胞凋亡的影響
流式細胞術檢測結果顯示,與0 h比較,OGD/R處理6、12、24 h神經元凋亡細胞及凋亡率均增加,差異均有統計學意義(P<0.01)。詳見圖5、圖6。

圖5 流式細胞術檢測HT22神經元細胞凋亡情況

圖6 OGD/R處理對HT22神經元細胞凋亡率的影響
2.4 miR-145-5p與FGF5的靶向關系
miR-145-5p在miRTar base、Target scan、Target miner及miRDB在線軟件中分別存在15 064、890、1 305、495個潛在靶基因,經過繪制Venn圖并取其交集共有100個潛在的靶基因,FGF5為其中之一。詳見圖7。

圖7 Venn圖
采用Targetscan 驗證miR-145-5p與FGF5的靶向關系,結果顯示,miR-145-5p與FGF5的3′-UTR有互補序列。熒光素酶報告結果顯示,WT FGF5 3′-UTR與miR-145-5p mimic共轉染后,熒光素酶的活性明顯降低;MUT FGF5 3′-UTR與miR-145-5p mimic共轉染后,熒光素酶的活性變化不明顯。與OGD/R比較,miR-145-5p inhibitor明顯上調FGF5表達,FGF5 siRNA明顯下調FGF5表達,差異均有統計學意義(P<0.01)。詳見圖8、圖9。

圖8 熒光素酶實驗驗證FGF5與miR-145-5p的靶向關系

圖9 miR-145-5p靶向負向調控FGF5表達
2.5 miR-145-5p inhibitor 抑制OGD/R誘導的HT22細胞凋亡
與OGD/R組比較,OGD/R+miR-145-5p inhibitor組HT22細胞凋亡率明顯降低(P<0.01),OGD/R+FGF5 siRNA組HT22細胞凋亡率明顯增加(P<0.01)。OGD/R+miR-145-5p inhibitor+FGF5 siRNA組與OGD/R+FGF5 siRNA組比較,HT22細胞凋亡率明顯降低(P<0.01)。詳見圖10、圖11。

圖10 流式細胞術檢測HT22神經元細胞凋亡情況

圖11 miR-145-5p inhibitor 抑制OGD/R誘導的HT22細胞凋亡
2.6 miR-145-5p inhibitor 減輕OGD/R誘導的氧化應激損傷
細胞氧化應激損傷檢測結果顯示,與OGD/R組比較,OGD/R+miR-145-5p inhibitor組細胞MDA、ROS和LDH含量均明顯降低,SOD活性明顯升高,差異均有統計學意義(P<0.01);OGD/R+FGF5 siRNA組細胞MDA、ROS和LDH含量均明顯增加,SOD活性明顯降低,差異均有統計學意義(P<0.01)。OGD/R+miR-145-5p inhibitor+FGF5 siRNA組與OGD/R+FGF5 siRNA組比較,細胞MDA、ROS和LDH含量均明顯降低,SOD活性明顯升高,差異均有統計學意義(P<0.01)。詳見圖12、圖13。

圖12 miR-145-5p inhibitor對OGD/R誘導的LDH、SOD水平的影響

圖13 miR-145-5p inhibitor對OGD/R誘導的ROS、MDA水平的影響
3 討 論
I/R是一個復雜的生理病理過程,涉及多種發病機制,如神經元的死亡、膠質細胞的激活和炎癥反應等。miRNA在大腦功能中扮演重要角色,包括神經發生、神經發育、導致突觸可塑性發生改變的細胞反應等。研究表明,miR-145-5p在血管內皮細胞損傷、心肌細胞存活及H2O2所致神經元細胞損傷等過程中均明顯上調[10-11],小膠質細胞OGD/R處理 2 h后和大腦中動脈栓塞(MCAO)12 h后大腦皮質中miR-145-5p也明顯上調。以上研究表明,miR-145-5p可能在OGD/R過程中損傷神經元細胞。本實驗結果顯示,對HT22細胞進行氧糖剝奪復氧不同時間后,細胞ROS、MDA、LDH含量明顯升高,SOD的活性明顯降低,提示造模成功。I/R發生后機體氧化和抗氧化能力失衡,發生氧化應激反應并產生大量氧自由基,清除自由基、抑制氧化應激反應是治療I/R的重要方式之一。本研究結果顯示,在I/R細胞模型中,miR-145-5p表達增加,抑制miR-145-5p表達后,細胞氧化應激反應得到緩解。研究表明,氧自由基可誘導產生大量細胞因子,從而刺激腦血管內皮細胞黏附因子的表達,引起再灌注的大量炎癥反應,最終導致神經細胞的凋亡和壞死[12]。ROS是高含氧活性小分子,有效清除過剩的ROS可改善缺血性腦卒中轉歸。SOD是清除氧自由基的蛋白酶,可通過磷酸化蛋白激酶B(AKT)降低缺血腦損傷導致的細胞凋亡[13]。本研究中,神經細胞經氧糖剝奪后復氧,細胞ROS、MDA及細胞上清LDH含量隨復氧時間的增加而增加,而SOD活性則隨復氧時間的增加而降低。
成熟miRNA通過與靶mRNA的3′-UTR的互補結合調節基因表達,此種調節作用將導致靶mRNA降解或蛋白質翻譯抑制,從而使細胞功能發生變化[14]。本研究中I/R損傷細胞miR-145-5p表達升高,并且FGF5受到抑制,通過生物信息學分析及熒光素酶實驗證實兩者之間具有較強的靶向關系,HT22細胞轉染miR-497-5p inhibitor后OGD/R,細胞中氧化應激、炎癥反應和細胞凋亡明顯減輕,而FGF5 siRNA轉染HT22細胞后,細胞氧化應激反應及細胞凋亡增加,提示miR-145-5p靶向FGF5,調節I/R損傷誘導的細胞凋亡和氧化應激損傷。
FGF5屬于成纖維細胞生長因子家族(FGFs)成員,在小鼠胚胎和成年小鼠的中樞神經系統及原腸胚、肌肉、運動神經元形成中被檢測到[15]。有研究表明,成年小鼠FGF5的信息水平在海馬、脊髓和大腦皮層比在其他神經節、小腦、中腦和后腦略高,可視為神經元或神經膠質持續釋放的營養因子[16]。FGF5具有多種生物學功能,并發揮重要作用。Walter等[17]研究表明,FGF5及其主要受體FGFR1介導的信號可作為人多形性膠質母細胞瘤的治療靶點。Fang等[18]研究結果表明,miR-188-5p通過靶向FGF5基因抑制肝癌細胞的增殖和轉移,在肝癌中起抑癌作用。此外,FGF5在高血壓疾病中也發揮重要作用,FGF5基因內部或附近區域是原發性高血壓的易感區域,原發性高血壓外周血中FGF5基因T/C突變與舒張壓和收縮壓呈正相關,提示FGF5 mRNA和蛋白質水平異常表達可能與高血壓疾病的發生發展有關[19-20]。本研究結果顯示,FGF5在OGD/R誘導的HT122中表達減少,并通過Targetscan證明FGF5是miR-145-5p的下游靶點,此外,miR-145-5p inhibitor轉染HT22細胞后,FGF5蛋白表達增加,細胞凋亡和氧化應激損傷均得以改善。而FGF5 siRNA轉染HT22細胞后,FGF5蛋白表達減少,氧化應激反應加強。以上結果提示miR-145-5p通過抑制FGF5表達從而增加OGD/R環境下HT22細胞的凋亡和損傷。
綜上所述,miR-145-5p可通過靶向結合FGF5誘導HT22細胞凋亡和氧化應激損傷,miR-145-5p可能是I/R的潛在治療靶點。但本研究僅通過體外細胞培養探討相關機制,關于miR-145-5p對I/R的作用和影響機制還需進一步深入研究。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
