LncRNA TUC338通過激活EGFR/PI3K/AKT 信號通路促進肺癌細胞的增殖、遷移和侵襲*
2023-09-25 11:29:12夏航彪趙界王述紅
西部醫學 2023年9期
夏航彪 趙界 王述紅
(遂寧市中心醫院呼吸與危重癥醫學科,四川 遂寧 629000)
肺癌是發病率與死亡率最高的惡性腫瘤,其中主要病理類型為非小細胞肺癌(Non-small cell lung cancer,NSCLC),約占肺癌患者85%以上,盡管肺癌的治療手段取得一定進步,仍有部分患者發生耐藥、復發轉移等問題,因此研究肺癌的發生發展與轉移的分子機制具有重要意義[1-2]。長鏈非編碼RNA(long non-coding RNA,lncRNA)為內源性非編碼RNA,研究顯示,多種lncRNA在肺癌中異常表達,與肺癌患者預后及臨床病理特征、侵襲和轉移有關[3-5]。LncRNA TUC338為新發現的lncRNA,研究顯示,它在前列腺癌、肺癌、宮頸癌等多種腫瘤中表達上調[6-8],發揮促癌基因的作用,但LncRNA TUC338在NSCLC中的作用機制尚不清楚。表皮生長因子受體(Epidermal growth factor receptor,EGFR)可調控腫瘤細胞增殖,在肺癌中高表達,與腫瘤的惡性轉化有關[9],且EGFR高表達可激活磷酸肌醇3激酶/絲氨酸/蘇氨酸蛋白激酶(Phosphoinositide 3-kinase/serine/threonine protein kinase,PI3K/AKT)通路,在腫瘤細胞的增殖、侵襲、遷移等過程中發揮重要作用[10-11]。本研究將探討LncRNA TUC338對肺癌細胞增殖、遷移及侵襲的影響及其作用機制,現報告如下。
1 資料與方法
1.1 一般資料
1.1.1 病例 收集2019年7月—2021年6月我院胸外科手術切除的37例NSCLC患者肺癌組織與癌旁組織標本,均為肺腺癌,患者手術前未經任何抗腫瘤治療,其中男性15例,女性22例,年齡32~75歲。取得標本在液氮罐中長期保存,本研究得到本院倫理委員會的批準(批準號:1905-04),患者知情同意并簽署受試者簽署的知情同意書。
1.1.2 主要材料 人肺腺癌細胞系NCI-H157、A549、NCI-H1299、HCC827及正常肺上皮細胞系BEAS-2B(中國科學院細胞庫);RPMI 1640培養基(Gibco公司);FBS、DMSO(北京Solarbio公司);Lipofectamine 3000轉染試劑(Sigma-Aldrich公司);抑制lncRNA TUC338表達(si-TUC338)及陰性對照(si-NC)質粒,lncRNA TUC338過表達(pcDNA-TUC338)及陰性對照(pcDNA-NC)質粒、以及RT-qPCR引物(GenePharma公司);EGFR與PI3K雙重抑制劑MTX-211(百奧萊博);CCK-8檢測試劑盒、ECL試劑、RIPA裂解液(江蘇凱基生物技術股份有限公司);兔抗人cyclin D1(ab16663)、MMP-2(ab92536)、β-actin抗體(ab8227)、MMP-9(ab76003)、p-EGFR(ab40815)、EGFR(ab52894)、p-PI3K(ab278545)、PI3K(ab32089)、p-AKT(ab38449)、AKT(ab8805)單克隆抗體(英國Abcam公司)。
1.2 方法
1.2.1 細胞的分組及處理 取對數增殖期的NCI-H157細胞,以每孔2×105個細胞接種于6孔板中,細胞匯合約85%時,將NCI-H157細胞分為對照(control)組、si-TUC338組、si-NC組、pcDNA-TUC338、pcDNA-NC組、MTX-211組及pcDNA-TUC338+MTX-211組,進行質粒轉染,轉染后將細胞在37°C、5% CO2培養箱中培養48 h后,RT-qPCR檢測各組NCI-H157細胞中lncRNA TUC338表達,鑒定是否轉染成功。
1.2.2 RT-qPCR檢測lncRNA TUC338水平 RNA提取試劑提取總RNA,紫外分光光度計測量RNA純度及含量,以RNA作為模板,合成cDNA后,采用RT-qPCR檢測lncRNA TUC338相對表達量,以GAPDH為內參,采用2-△△Ct法計算lncRNA TUC338相對表達量。LncRNA TUC338上游引物(5′-3′):GGTGAGAGGGGATGTTCAGT,下游引物(5′-3′):TGGGTGAAATGAGGTTG;GAPDH上游引物(5′-3′):CTGGGCTACACTGAGCACC,下游引物(5′-3′):AAGTGGTCGTTGAGGGCAATG。
1.2.3 CCK-8法檢測細胞增殖 將轉染后的細胞按照每孔1×105個細胞接種于96孔板中,分別培養24、48和72 h后,每孔加入適量的CCK-8溶液避光孵育2h,于酶標儀450 nm處檢測吸光度(A)值。
1.2.4 Transwell小室法檢測細胞遷移與侵襲能力 使用無血清培養基制備NCI-H157細胞懸液,以每孔2×104個細胞接種于Transwell上室中,遷移實驗使用常規Transwell小室的上室,侵襲實驗使用Matrigel基質膠包被Transwell小室的上室,下室為含10% FBS的RPMI 1640培養基,培養24 h后,使用4%多聚甲醛固定,0.1%結晶紫染色,顯微鏡下隨機選取6個視野觀察穿膜細胞數。
1.2.5 Western blot檢測cyclin D1、MMP-2、MMP-9、p-EGFR、p-PI3K、p-AKT蛋白表達 提取各組細胞總蛋白,BCA法進行蛋白濃度定量后,取30 μg蛋白煮沸后進行SDS-PAGE并轉膜,5%脫脂奶粉封閉2 h,加入cyclin D1、MMP-2、MMP-9、p-EGFR、EGFR、p-PI3K、PI3K、p-AKT、AKT一抗,在4 ℃孵育過夜,加入HRP標記的羊抗兔二抗室溫孵育2 h,ECL試劑顯影,以β-actin為內參,用Quantity One 4.6軟件分析圖像,分析目的蛋白相對表達水平。
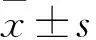
2 結果
2.1 LncRNA TUC338在肺癌組織與細胞中表達上調 LncRNA TUC338在肺癌組織表達水平顯著高于癌旁組織(t=41.799,P<0.05)。LncRNA TUC338在人NSCLC細胞系NCI-H157、A549、NCI-H1299、HCC827中表達水平顯著高于BEAS-2B細胞(t=10.431、11.023、20.762、12.928,P<0.05),其中在NCI-H157細胞中表達水平最高,選擇NCI-H157細胞進行后續實驗,見圖1。
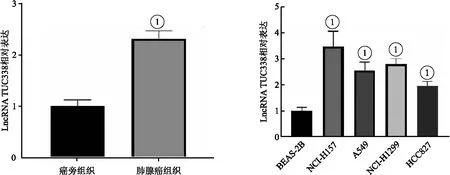
圖1 LncRNA TUC338在肺癌組織與細胞中表達
2.2 各組NCI-H157細胞中 lncRNA TUC338的表達 與si-NC組相比,si-TUC338組lncRNA TUC338表達水平顯著降低;與pcDNA-NC組相比,pcDNA-TUC338組lncRNA TUC338表達水平顯著升高(均P<0.05);control組與MTX-211組、pcDNA-TUC338與pcDNA-TUC338+MTX-211組之間lncRNA TUC338表達水平差異無統計學意義(P>0.05)。見圖2。
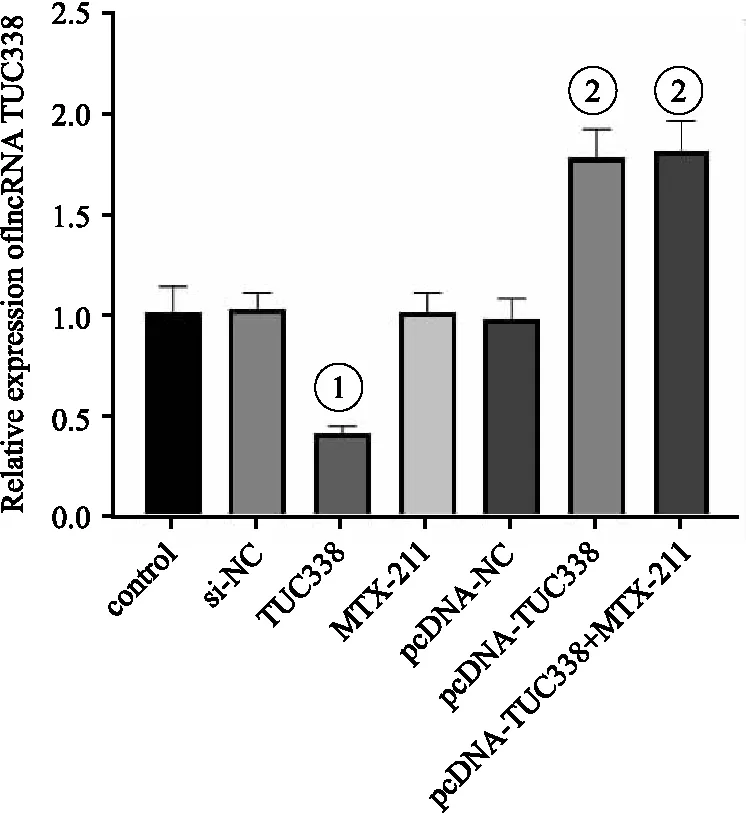
圖2 各組NCI-H157細胞中lncRNA TUC338的表達情況
2.3 各組NCI-H157細胞增殖活性 與對照組相比,MTX-211組細胞增殖活性顯著降低;與si-NC組相比,si-TUC338組細胞增殖活性顯著降低(均P<0.05);與pcDNA-NC組相比,pcDNA-TUC338組細胞增殖活性顯著升高;與pcDNA-TUC338組相比,pcDNA-TUC338+MTX-211組細胞增殖活性顯著降低(均P<0.05)。見圖3。
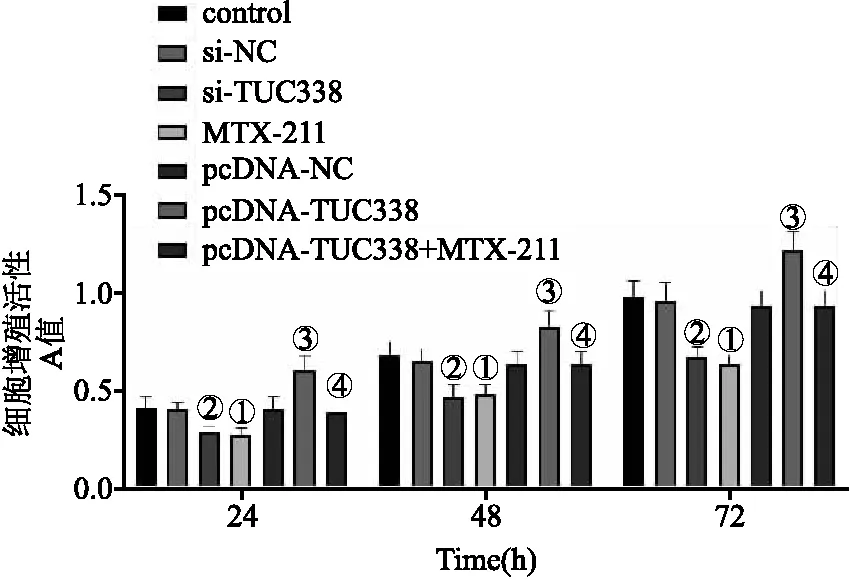
圖3 各組NCI-H157細胞增殖活性比較
2.4 各組NCI-H157細胞遷移與侵襲能力 與對照組相比,MTX-211組細胞遷移與侵襲數顯著降低;與si-NC組相比,si-TUC338組細胞遷移與侵襲數顯著降低(均P<0.05);與pcDNA-NC組相比,pcDNA-TUC338組細胞遷移與侵襲數顯著升高;與pcDNA-TUC338組相比,pcDNA-TUC338+MTX-211組細胞遷移與侵襲數顯著降低(均P<0.05)。見圖4。
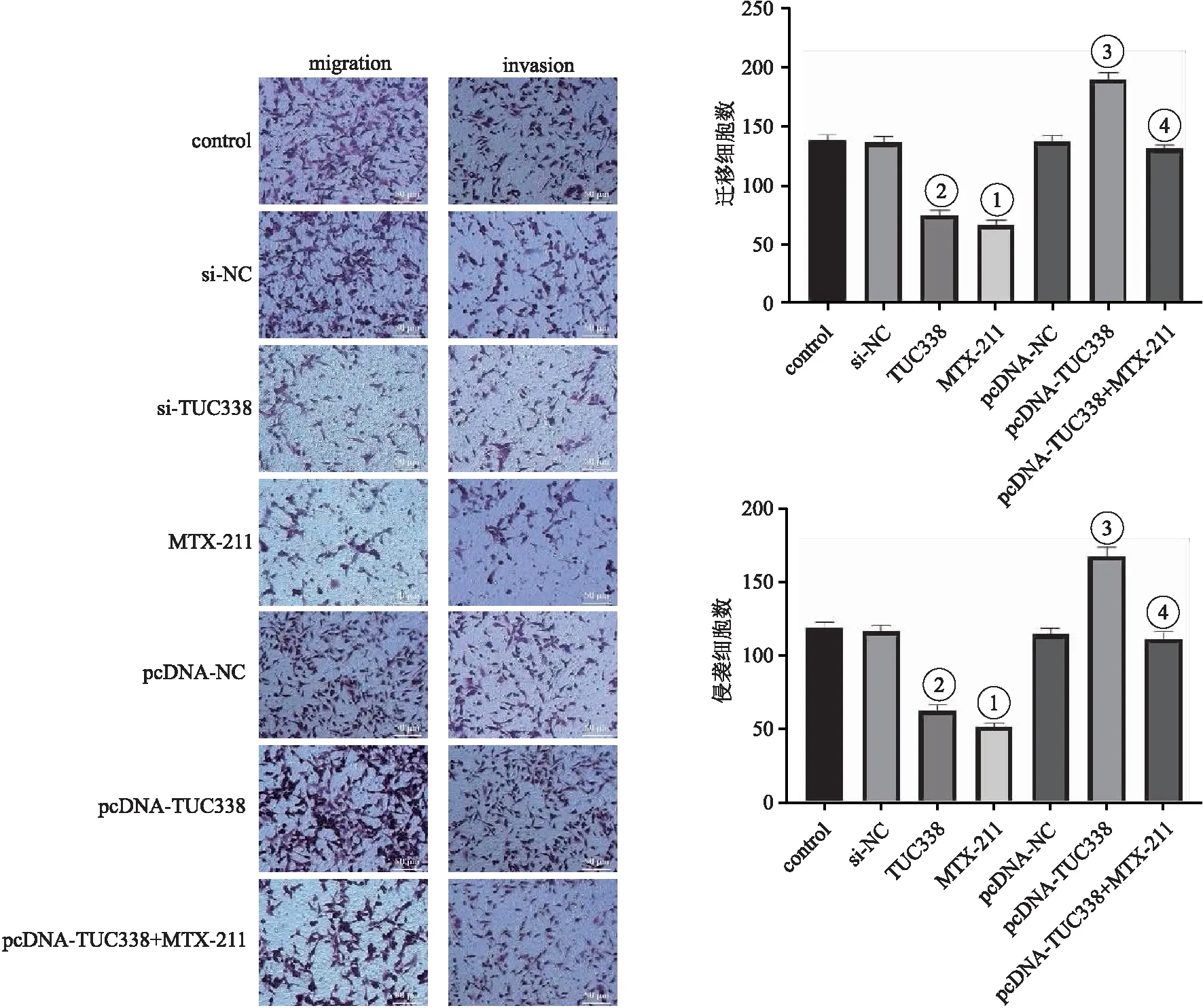
圖4 各組NCI-H157細胞遷移、侵襲(結晶紫染色,200×)
2.5 各組NCI-H157細胞cyclin D1、MMP-2、MMP-9蛋白表達 與對照組相比,MTX-211組細胞cyclin D1、MMP-2、MMP-9蛋白表達水平顯著降低;與si-NC組相比,si-TUC338組細胞cyclin D1、MMP-2、MMP-9蛋白表達水平顯著降低(均P<0.05);與pcDNA-NC組相比,pcDNA-TUC338組細胞cyclin D1、MMP-2、MMP-9蛋白表達水平顯著升高;與pcDNA-TUC338組相比,pcDNA-TUC338+MTX-211組細胞cyclin D1、MMP-2、MMP-9蛋白表達水平顯著降低(均P<0.05)。見圖5。
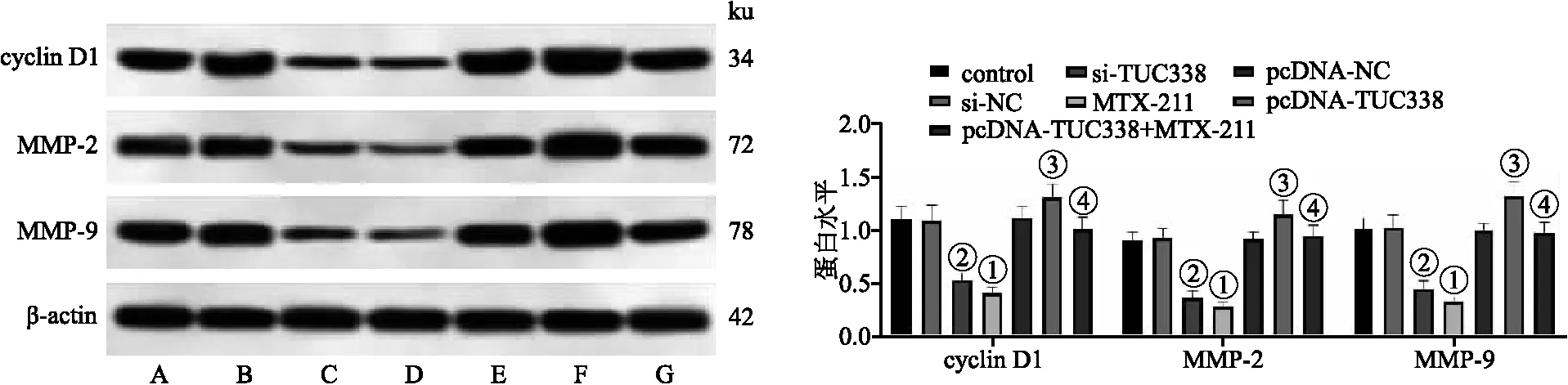
圖5 各組NCI-H157細胞中cyclin D1、MMP-2、MMP-9蛋白表達
2.6 各組NCI-H157細胞EGFR/PI3K/AKT通路相關蛋白表達 與對照組相比,MTX-211組細胞p-EGFR/EGFR、p-PI3K/PI3K、p-AKT/AKT蛋白表達水平顯著降低;與si-NC組相比,si-TUC338組細胞p-EGFR/EGFR、p-PI3K/PI3K、p-AKT/AKT蛋白表達水平顯著降低(均P<0.05);與pcDNA-NC組相比,pcDNA-TUC338組細胞p-EGFR/EGFR、p-PI3K/PI3K、p-AKT/AKT蛋白表達水平顯著升高;與pcDNA-TUC338組相比,pcDNA-TUC338+MTX-211組細胞p-EGFR/EGFR、p-PI3K/PI3K、p-AKT/AKT蛋白表達水平顯著降低(均P<0.05)。見圖6。
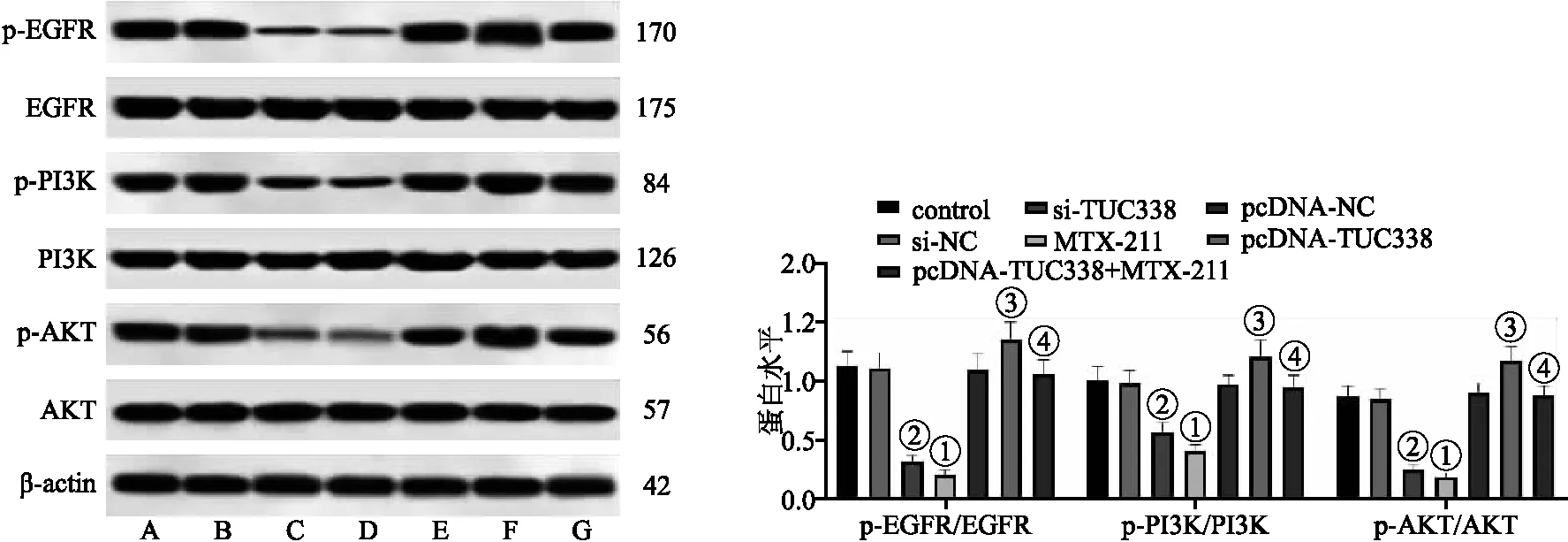
圖6 各組NCI-H157細胞中EGFR/PI3K/AKT通路相關蛋白表達
3 討論
LncRNA在肺癌的發生發展和轉移過程中發揮重要作用,參與調控肺癌細胞增殖、遷移及侵襲、細胞周期、血管生成等過程[12]。有研究顯示,LncRNA TUC338在肝癌、前列腺癌中上調表達并介導腫瘤的惡性進展[13-14]。已有研究顯示,LncRNA TUC338在肺癌組織中高表達,與腫瘤大小、淋巴結轉移和患者總生存率有關,抑制LncRNA TUC338表達可抑制肺癌細胞增殖與侵襲能力,而過表達LncRNA TUC338則可增加肺癌細胞增殖與侵襲能力[7]。為進一步研究LncRNA TUC338在肺癌中的具體作用機制,本研究分別檢測肺癌組織及癌旁組織和NCI-H157、A549、NCI-H1299、HCC827細胞中LncRNA TUC338表達情況,結果顯示LncRNA TUC338在人肺癌組織中上調表達,且在NCI-H157細胞中表達水平最高,選擇NCI-H157細胞進行后續實驗。Qian等[15]研究表明,在食管癌細胞中下調lncRNA TUC338表達可抑制食管癌細胞的遷移和侵襲。本研究在NCI-H157細胞中抑制lncRNA TUC338表達后,NCI-H157細胞增殖活性、遷移與侵襲數、Cyclin D1、MMP-2、MMP-9表達水平顯著降低,表明抑制lncRNA TUC338表達可顯著抑制NCI-H157細胞增殖、遷移與侵襲。而在NCI-H157細胞中過表達lncRNA TUC338后,NCI-H157細胞增殖活性、遷移與侵襲數、Cyclin D1、MMP-2、MMP-9表達水平顯著升高,表明lncRNA TUC338在肺癌中發揮促癌基因的作用,這與Zhang等[7]研究結果一致,然而其具體作用機制尚不清楚。
EGFR在多數腫瘤中上調表達,EGFR成為腫瘤治療的重要分子靶標[16-17]。EGFR與相應配體特異性結合后,引起受體酪氨酸激酶的磷酸化,激活PI3K/AKT途徑,PI3K/AKT信號通路可參與調控細胞的分裂和增殖及血管的形成,還與腫瘤的侵襲和轉移有關[18-19]。激活EGFR/PI3K/AKT通路可促進腫瘤的惡性特征[20-22]。研究表明,LINC00520可通過下調EGFR,使PI3K/AKT信號通路失活,從而阻止皮膚鱗狀細胞癌細胞的增殖、遷移[23]。本研究中,NCI-H157細胞中抑制lncRNA TUC338表達后,細胞p-EGFR、p-PI3K、p-AKT蛋白表達水平顯著降低,與加入EGFR/PI3K/AKT通路抑制劑MTX-211結果類似,表明抑制lncRNA TUC338表達可顯著抑制EGFR/PI3K/AKT通路激活。而在NCI-H157細胞中過表達lncRNA TUC338后NCI-H157細胞p-EGFR、p-PI3K、p-AKT蛋白表達水平顯著升高,且加入MTX-211后可逆轉這一現象,提示過表達lncRNA TUC338可激活EGFR/PI3K/AKT通路。Li等[24]研究表明,lncRNA TUC338通過抑制miR-28-5p表達激活EGFR/PI3K/AKT通路促進彌漫大B細胞淋巴瘤細胞增殖,本研究與此結果類似,因此推測lncRNA TUC338通過激活EGFR/PI3K/AKT通路,促進NCI-H157細胞增殖、遷移與侵襲。
4 結論
綜上所述,lncRNA TUC338在肺癌組織中上調表達,lncRNA TUC338可通過激活EGFR/PI3K/AKT信號通路,促進肺癌細胞增殖、遷移與侵襲。但本研究僅選擇一種細胞系進行研究,后續仍需選擇多種細胞系對此通路進行驗證。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
