擬干酪乳桿菌全合成培養基氨基酸及維生素組分優化研究
2023-10-09 06:43:50施盛超劉志豪王永紅
食品與發酵工業 2023年18期
施盛超,劉志豪,王永紅
(華東理工大學,生物反應器工程國家重點實驗室,上海,200237)
乳酸分子結構相對簡單,是一種極其重要的生物基化學品,在食品行業,化妝品,醫藥以及化工等傳統行業具有廣泛的應用[1],由其合成的聚乳酸在新材料行業也具有重要的應用價值[2]。目前乳酸的生產以微生物發酵為主[3],擬干酪乳桿菌是具有良好乳酸生產能力的工業乳酸菌,能夠在高濃度葡萄糖條件下高效生產L-乳酸[4-5],具有廣闊的應用前景。
然而,目前對于擬干酪乳桿菌的培養主要使用復合培養基或成分比較復雜的合成培養基[6-7],培養基的復雜性對擬干酪乳桿菌生理特性及其代謝調控的研究增加了困難。在全合成培養基中氨基酸類和維生素類是種類比較多的營養物質,對菌體的生長和代謝具有重要作用。氨基酸對于微生物的作用除了是蛋白質的基礎組成單位外,還能在微生物生長過程中提供生長所需的能量,促進微生物能量代謝,更是很多生物體內非常重要的含氮化合物合成前體[8-10]。維生素是細胞多種酶的輔酶,參與維持和調節細胞的生理代謝[11]。因此更加合理的添加這些營養物質,對于優化菌體生長,提高乳酸產量具有重要意義。
本研究旨在基于已有的乳酸發酵全合成培養基,通過對培養基中各種氨基酸以及維生素對擬干酪乳桿菌代謝影響的研究,在保持至少不低于原全合成培養基的底物利用率以及乳酸產率的前提下,盡可能減少培養基組分,提高合成乳酸效率,為今后該菌株的代謝通量定量分析和代謝工程研究奠定基礎。
1 材料與方法
1.1 菌種
擬干酪乳桿菌(Lactobacillusparacasei)由本實驗室保藏。
1.2 培養基與培養條件
擬干酪乳桿菌經一級與二級斜面培養基活化[12],用50 mL超純水重懸二級斜面菌體,將15 mL菌懸液轉接至裝有85 mL MRS培養基的250 mL搖瓶中作為種子瓶[13]。以20%的接種量接種于全合成培養基[14]。搖瓶培養的條件為溫度37 ℃,轉速130 r/min,培養48 h。2.5 L發酵罐的培養條件為發酵體積1.5 L,攪拌轉速150 r/min,溫度37 ℃,pH值維持在6.0左右,發酵周期為48 h。
1.3 檢測方法
1.3.1 菌體光密度
在Spectrumlab 22 PC分光光度計620 nm下測定吸光值(OD值)。
1.3.2 葡萄糖濃度
采用葡萄糖氧化酶法試劑盒進行測定。
1.3.3 乳酸濃度
采用Agilent高效液相色譜檢測,色譜柱為Hi-Plex H (300 mm×7.7 mm);檢測波長為210 nm;流動相為0.01 mol/L硫酸;流速為0.4 mL/min;進樣量為10 μL;柱溫為50 ℃。
1.4 合成培養基優化方法
1.4.1 單一缺失實驗
對全合成培養基中氨基酸和維生素進行單一缺失搖瓶實驗,確定對菌體生長和乳酸產量有顯著影響的組分。
1.4.2 單組分濃度梯度實驗
對該組分的添加量設置為初始濃度的倍數,0、0.5、1、2、4倍,其余組分濃度保持不變。
1.4.3 Plackett-Burman實驗
該方法在單組分濃度梯度實驗的基礎上,對每個因素取高(1)、低(-1)2種水平,以考察每種因素的影響水平及影響的顯著性,以期從眾多影響因素中篩選出顯著影響乳酸生產的因素。
1.4.4 最陡爬坡實驗
根據Plackett-Burman實驗篩選出顯著影響合成乳酸的因素,結合各因素效應的正負,設置合適的步長和爬坡方向,進而有效地確定各因素具有最大響應值的濃度范圍。
1.4.5 響應面分析實驗
采用Box-Behnken實驗設計方法,每個因素取3個水平:-1,0,1。建立多元回歸方程并計算出各因素的最佳濃度值。Plackett-Burman實驗和Box-Behnken實驗均通過Design Expert 8.0軟件進行實驗設計和結果分析。
2 結果與分析
2.1 產L-乳酸的合成培養基中氨基酸的優化
本實驗在全合成培養基的基礎上對20種氨基酸進行單一缺失實驗,發現半胱氨酸、谷氨酸、谷氨酰胺、天冬氨酸、天冬酰胺、酪氨酸、賴氨酸、丙氨酸單一缺失后的菌體生長和乳酸產量降低最多(圖1)。對這8種氨基酸進行單組分濃度梯度實驗,結果顯示(圖2)隨著氨基酸濃度的增加,其對菌體的生長和產酸存在不同程度的影響,不同氨基酸的結果也存在顯著差異。半胱氨酸、谷氨酸、谷氨酰胺的濃度變化對菌體生長和產酸有明顯的促進作用,但高濃度的氨基酸導致菌體生長過盛,從而使乳酸生成受到影響。天冬氨酸、天冬酰胺的濃度變化對菌體生長有一定影響,當濃度大于1倍時,菌體生物量無變化,但與乳酸產量呈正相關。酪氨酸、賴氨酸濃度變化對菌體生長影響較小,濃度在1倍以下時,對乳酸生成有促進作用。丙氨酸濃度變化對菌體生長影響較小,低濃度的丙氨酸促進產酸,高濃度的丙氨酸抑制產酸。另外,從氨基酸濃度梯度實驗的數據可以看出天冬氨酸和天冬酰胺對于菌體生長和乳酸產量的影響作用相類似,考慮到微生物中廣泛存在天冬酰胺酶,可將天冬酰胺轉換成天冬氨酸[15],因此將天冬酰胺替換為相同濃度的天冬氨酸,其搖瓶實驗的菌體生長、乳酸生成以及糖耗情況與替換前基本一致,谷氨酸與谷氨酰胺之間也同樣存在轉換作用[16],實驗結果也相似,故選擇將谷氨酰胺替換成谷氨酸。因此最終確定半胱氨酸、酪氨酸、賴氨酸、丙氨酸、天冬氨酸、谷氨酸作為合成培養基中對生長和產酸有重要作用的氨基酸。
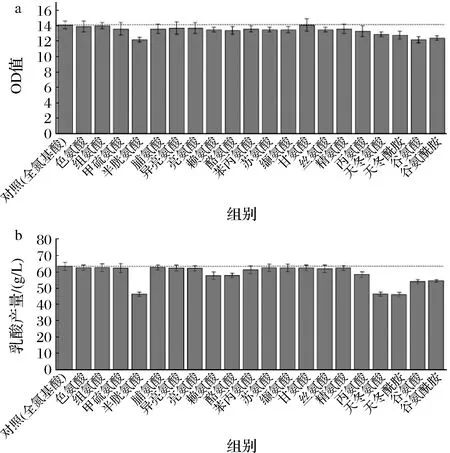
a-單一氨基酸缺失對生長的影響;b-單一氨基酸缺失對乳酸產量的影響圖1 單一氨基酸缺失對菌體生長和乳酸產量的影響Fig.1 Effects of single omission of amino acid on growth and lactic acid production
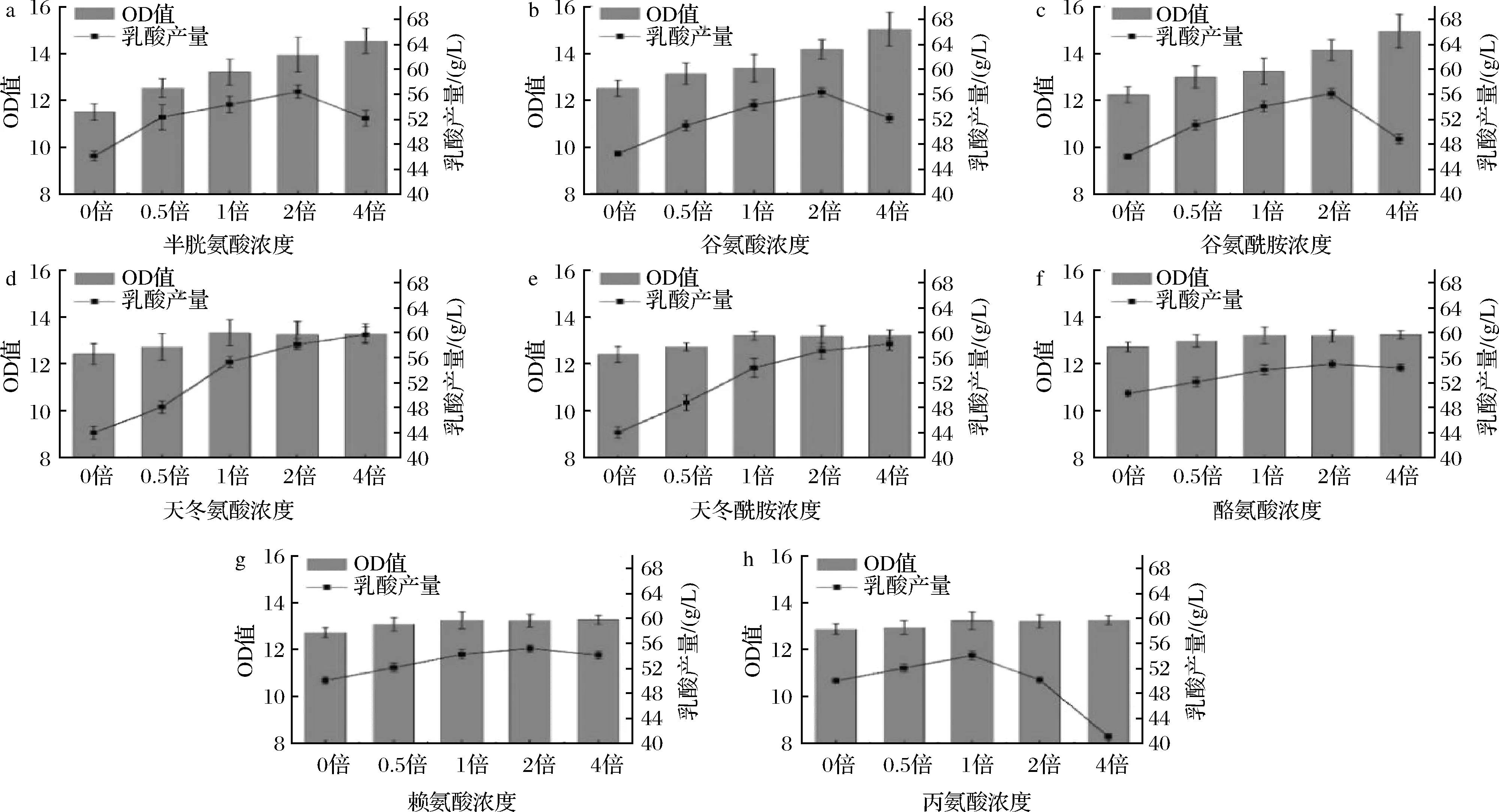
a-半胱氨酸;b-谷氨酸;c-谷氨酰胺;d-天冬氨酸;e-天冬酰胺;f-酪氨酸;g-賴氨酸;h-丙氨酸圖2 不同濃度梯度氨基酸對菌體生長和乳酸產量的影響Fig.2 Effects of different concentrations of amino acid on growth and lactic acid production
2.2 產L-乳酸的合成培養基中維生素的優化
維生素的單一缺失實驗同氨基酸,發現生物素、硫胺素、核黃素、煙酸、吡哆醇單一缺失后的菌體生長和乳酸產量降低最多(圖3)。對這5種維生素進行單組分濃度梯度實驗,結果顯示(圖4)不同的維生素隨著濃度的增加對菌體生長和產酸存在不同的影響。其中菌體生物量與生物素和硫胺素的濃度呈正相關,生物素的效果更加明顯,乳酸濃度也隨著與生物素、硫胺素的濃度增加而增加,但增加到原2倍濃度之后,乳酸濃度增加趨勢漸緩,生物素和硫胺素是微生物體內多種酶系統的輔酶,對糖酵解途徑中多種酶的活性有促進作用,能夠調節糖代謝,促進菌體生長,加快丙酮酸和乳酸的合成,在生物體的碳代謝循環中起到了至關重要的作用[17-18]。核黃素、煙酸、吡哆醇的濃度變化對菌體生長沒有影響,但是對產酸合成有一定促進作用,吡哆醇的濃度在0.5倍時,產酸量達到最大,煙酸及核黃素濃度達到1倍后,乳酸增長乏力,尤其是高濃度煙酸抑制了產酸,產酸量出現了較大的下降。核黃素是細胞黃酶類輔基的組成成分,在細胞的氧化還原反應中具有重要作用[19],煙酸是NAD+和NADH的關鍵前體物質,NADH作為乳酸脫氫酶的輔酶參與反應[20],因此核黃素和煙酸對于乳酸的生成具有重要的調節作用[21]。吡哆醇在生物體內可轉變為吡哆醛,與氨基酸的代謝密切相關[22],間接參與乳酸的代謝調控。因此最終確定生物素、硫胺素、煙酸對生長和產酸有重要作用。
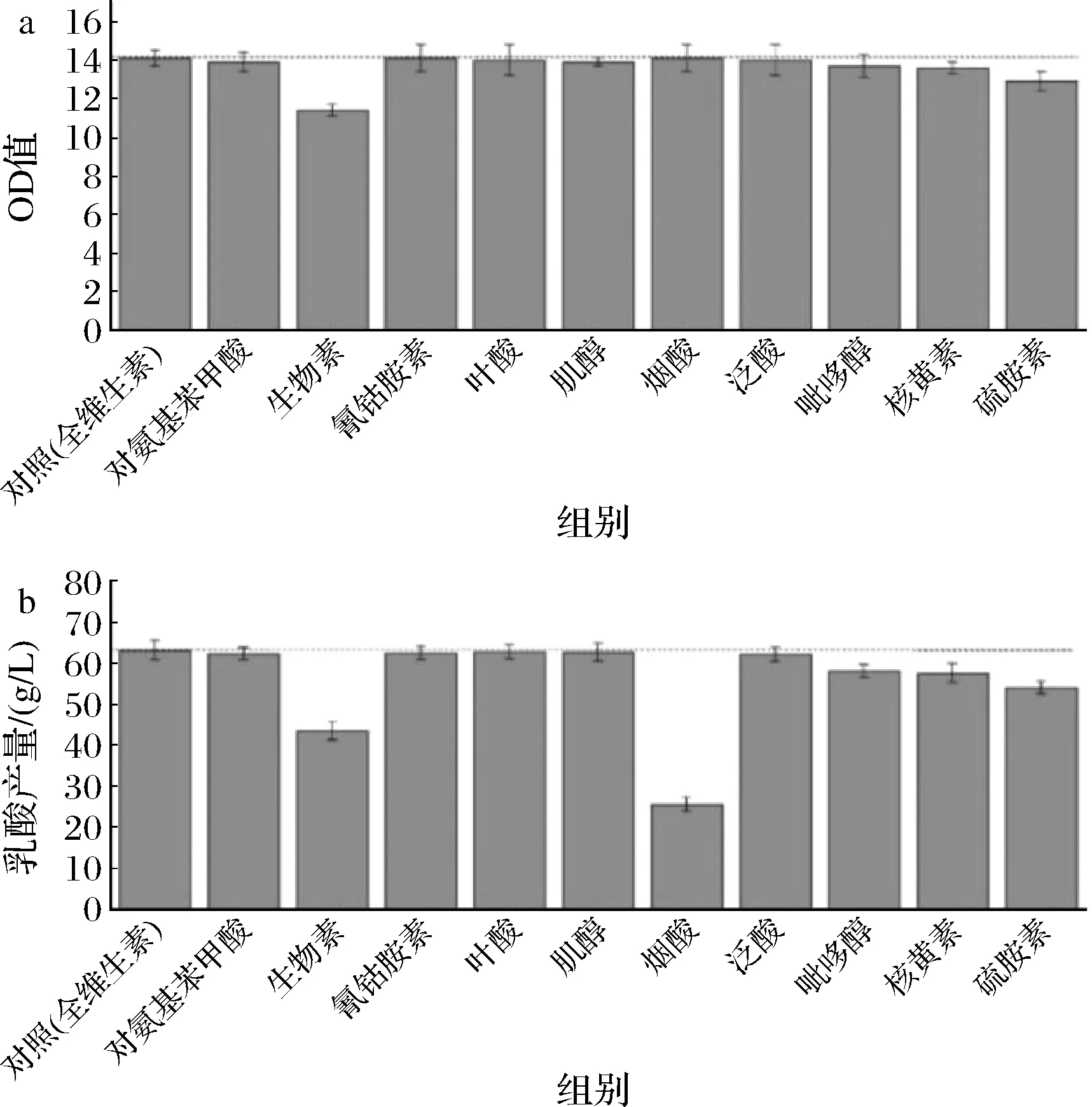
a-單一維生素缺失對生長的影響;b-單一維生素缺失對乳酸產量的影響圖3 單一維生素缺失對菌體生長和乳酸產量的影響Fig.3 Effects of single omission of vitamin on growth and lactic acid production
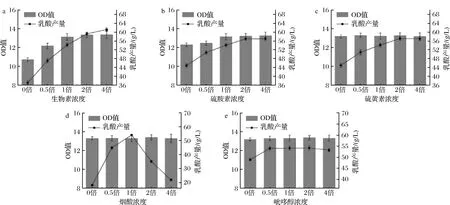
a-生物素;b-硫胺素;c-核黃素;d-煙酸;e-吡哆醇圖4 不同濃度梯度維生素對菌體生長和乳酸產量的影響Fig.4 Effects of different concentrations of vitamin on growth and lactic acid production
2.3 響應面分析優化合成培養基
在對6種氨基酸和3種維生素的單組分濃度梯度進行考察基礎上,進而通過響應面分析,建立乳酸生成與影響因素間的回歸關系,并最終獲得產酸穩定、高效的合成培養基配方。首先通過Plackett-Burman實驗分別以原濃度和原0.5倍濃度作為1和-1水平的值,實驗設計因素水平及結果見表1,通過顯著性分析(表2)發現,因素A(生物素)、H(谷氨酸)、J(天冬氨酸)對乳酸的生成有極顯著的影響,且均為正效應,得到線性回歸方程為:Y=34.69+5.38A+1.82H+1.68J,R2=0.938 1,因此確定生物素、谷氨酸、天冬氨酸作為下一步研究的影響因素。

表1 Plackett-Burman實驗設計與結果Table 1 Plackett-Burman experiment design and results

表2 偏回歸系數及影響因子的顯著性分析Table 2 Significance analysis of partial regression coefficients and factors
根據上述實驗的結果設計生物素、谷氨酸、天冬氨酸的最陡爬坡路徑,由于生物素、谷氨酸、天冬氨酸均為極顯著正效應,所以都要沿濃度增大的方向爬坡。結果如表3所示,隨著這3個因素濃度的增加,乳酸產量呈現出先升高后降低的趨勢,并以第4組實驗濃度值作為最陡爬坡實驗的中心濃度。
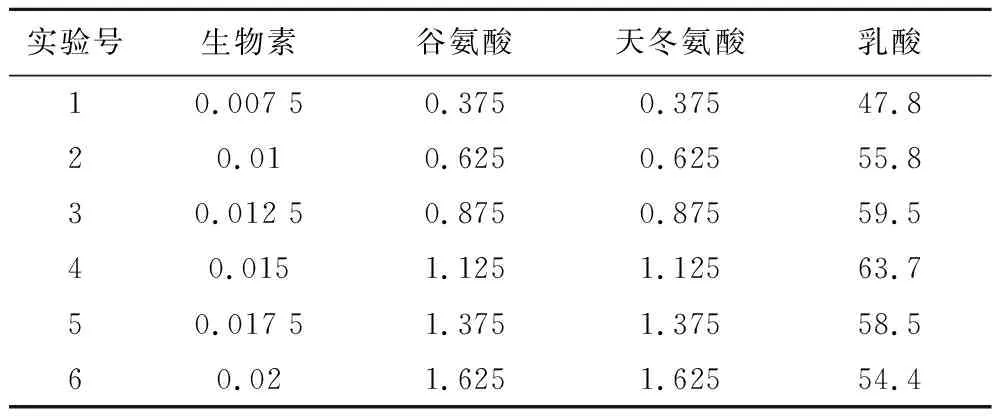
表3 最陡爬坡實驗設計及結果 單位:g/LTable 3 Design and results of the steepest climbing experiment
根據最陡爬坡實驗確定的3個影響因素的中心濃度,對其進行3因素、3水平的響應面實驗,分別以第2、4、6組的條件作為-1、0、1水平的值,實驗設計與結果見表4。
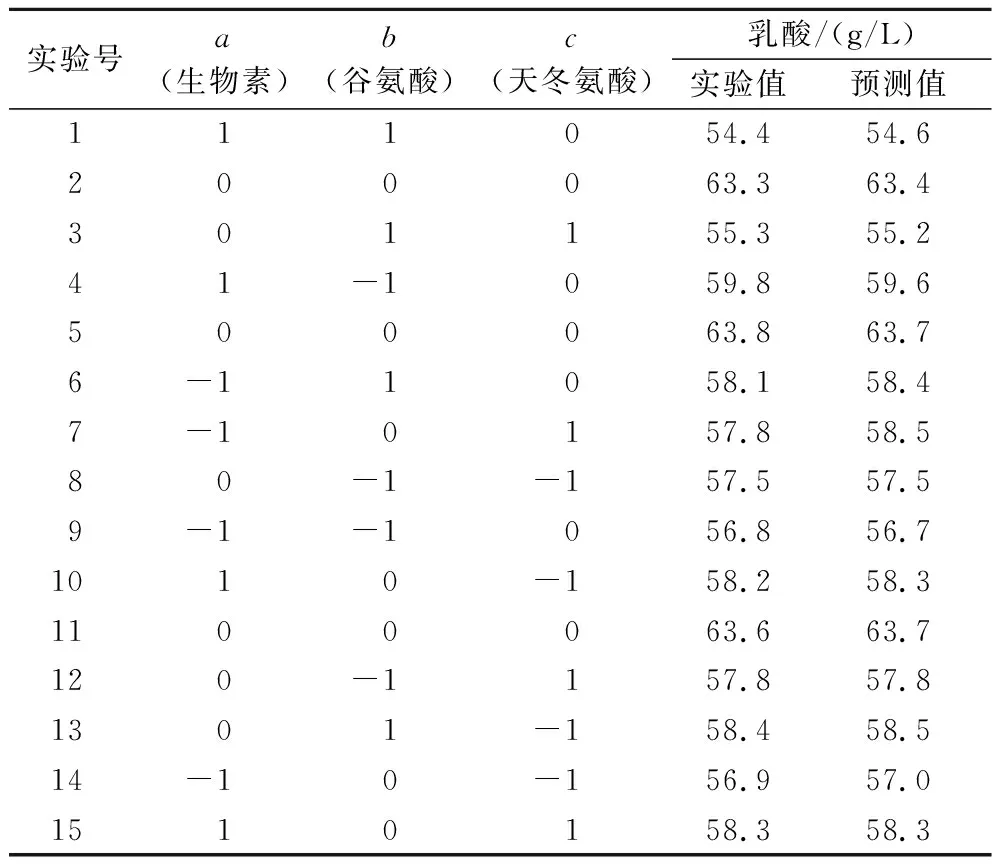
表4 Box-Behnken設計表及實驗結果Table 4 Design and results of Box-Behnken experiment
通過響應面的結果進而獲得乳酸濃度對生物素、谷氨酸和天冬氨酸的多元二次回歸方程為:Y=63.8+1.31a+0.77b-0.40c+0.23ab-0.025ac-0.87bc-2.74a2-3.49b2-3.19c2,R2=0.970 2,如表5所示,模型的P值小于0.05,失擬項大于0.05,說明模型顯著,擬合度較好。由表6回歸系數的顯著性檢驗可知,因素a、b、a2、b2對乳酸生成的影響極顯著(P<0.01),因素c2對乳酸生成的影響顯著(P<0.05),其他因素對乳酸生成的影響不顯著。
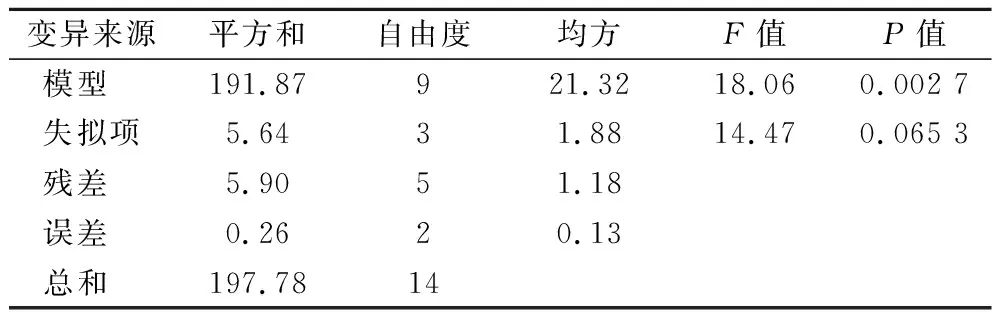
表5 回歸模型的方差分析Table 5 Variance analysis of regression model
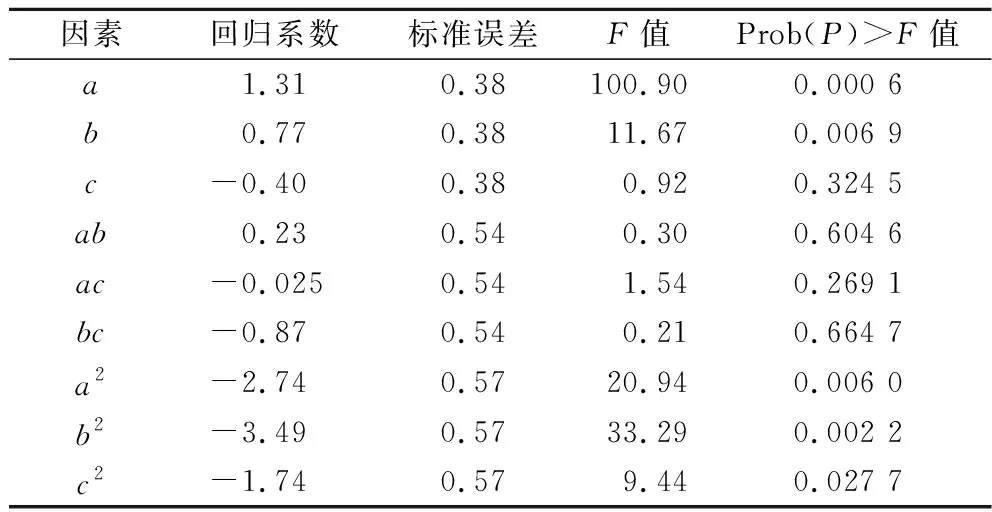
表6 二次模型回歸系數顯著性檢驗Table 6 Significance test for regression coefficients of quadratic model
對二次回歸方程求最值后換算可得生物素0.016 g/L,谷氨酸1.128 g/L,天冬氨酸1.120 g/L,在該濃度條件下模型預測的乳酸質量濃度為64.3 g/L,3次驗證實驗的平均乳酸質量濃度為64.1 g/L,與模型的預測值基本一致,說明模型對乳酸生成擬合效果較好。通過以上單組分濃度梯度實驗以及響應面分析實驗最終確定優化后合成培養基中的氨基酸和維生素質量濃度(g/L)分別為半胱氨酸0.375,酪氨酸0.125,賴氨酸0.3,丙氨酸0.125,天冬氨酸1.120,谷氨酸1.128,硫胺素0.007 5,煙酸0.007 5,核黃素0.007 5,吡哆醇0.007 5,生物素0.016。
2.4 合成培養基優化前后的搖瓶和2.5 L罐發酵水平比較
合成培養基優化前后搖瓶發酵結果如圖5所示,在初始葡萄糖質量濃度為80 g/L的條件下,菌體生長在前期并沒有太大變化,12 h之后,優化后的菌體生長速度明顯加快,最終OD值為14.5,高于對照的OD值13.2。乳酸產率在前期略有增長,發酵至12 h后,乳酸產量和產率出現了較大幅度的增長,發酵至48 h時乳酸產量從54.8 g/L增加到64.2 g/L,產率從1.04 g/(L·h)增加到1.25 g/(L·h),糖酸轉化率從81.9%增加到93.8%。
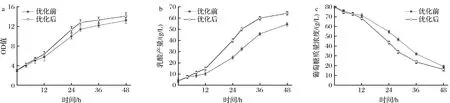
a-OD值隨培養時間的變化;b-乳酸產量隨培養時間的變化;c-葡萄糖濃度隨培養時間的變化圖5 合成培養基優化前后搖瓶發酵過程特性變化Fig.5 Profiles of fermentation process with original and optimized chemical defined medium in shake flasks
在初始葡萄糖質量濃度為200 g/L的條件下,用2.5 L發酵罐比較培養基優化前后的乳酸發酵情況(圖6),與250 mL搖瓶發酵不同的是,2.5 L罐OD值在24~28 h的時候達到最大值,在這之后OD值平穩中略有下降,優化前的OD值在24 h達到最大,為11.5,優化后OD值在28 h達到最大,為12.7,優化后比優化前提高了10.4%。發酵至12 h后,乳酸生產開始明顯增加,發酵至48 h時,乳酸產量從140.2 g/L 增加到165.5 g/L,增長幅度18%,產率從2.83 g/(L·h)增加到3.35 g/(L·h),糖酸轉化率從82.4%增加到97.6%,較搖瓶水平進一步提高。
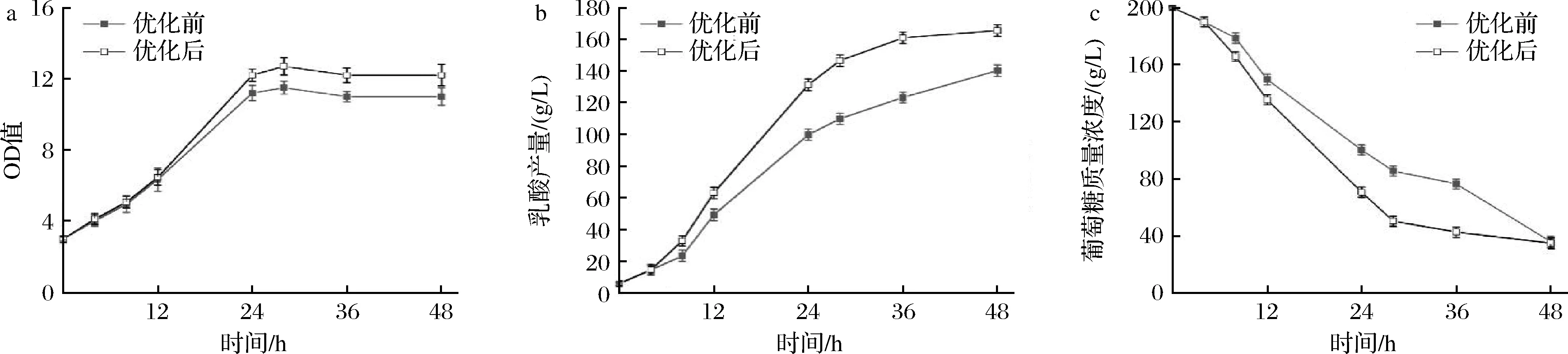
a-OD值隨培養時間的變化;b-乳酸產量隨培養時間的變化;c-葡萄糖濃度隨培養時間的變化圖6 合成培養基優化前后2.5 L反應器發酵過程特性變化Fig.6 Profiles of fermentation process with original and optimized chemical defined medium in 2.5 L bioreactor
3 結論與討論
本文優化了擬干酪乳桿菌乳酸發酵合成培養基成分中的氨基酸和維生素組分及濃度。利用單一缺失實驗確定了待優化的組分為8種氨基酸和5種維生素,而部分氨基酸和維生素的缺失并不會對菌體生長和產酸造成影響,說明這些組分并不是生長和代謝限制因素,或者本身可以合成相關組分滿足菌體生長和產酸需求,因此無需從外界環境中獲取。根據濃度梯度實驗并通過Plackett-Burman實驗確定了顯著影響因素為生物素、谷氨酸、天冬氨酸,運用響應面分析方法確定了三者質量濃度分別為0.016、1.128、1.120 g/L。使用優化后的合成培養基進行擬干酪乳桿菌乳酸發酵,其最終乳酸濃度無論是在搖瓶還是在2.5 L發酵罐中都要顯著高于優化前,在80 g/L葡萄糖的搖瓶中乳酸質量濃度達到64.3 g/L,在200 g/L葡萄糖的2.5 L發酵罐中乳酸質量濃度達到了165.5 g/L,糖酸轉化率分別提高至93.8%和97.6%,在大幅減少氨基酸和維生素種類的情況下,取得了更好的發酵效果。
優化后的培養基中生物素、谷氨酸、天冬氨酸的添加量提高,說明這3種物質對乳酸的生成有重要作用。其中谷氨酸和天冬氨酸是三羧酸循環的重要參與者[23],二者添加量的提高使得在減少碳源流向三羧酸循環的情況下仍能保持三羧酸循環的流量,加上生物素促進糖酵解途徑的作用,從而使得更多的碳源流向了乳酸生成。此外,由于代謝流實驗需要對胞內外的氨基酸成分進行定量檢測,而胞外氨基酸組分的減少排除了不必要氨基酸的檢測干擾,也為更有效的計算代謝流提供了極大便利,因此本實驗結果為后續擬干酪乳桿菌代謝流定量研究奠定了良好基礎。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
