m6A RNA甲基化修飾在昆蟲生長發育、免疫和抗藥性中的作用
2023-10-16 03:35:10張欣怡朱流紅張友軍郭兆將
植物保護 2023年5期
張欣怡, 白 楊, 郭 樂, 朱流紅,蘇 奇, 張友軍, 郭兆將*
(1. 長江大學農學院,荊州 434025;2. 中國農業科學院蔬菜花卉研究所,蔬菜生物育種全國重點實驗室,北京 100081)
RNA修飾是最普遍存在的表觀遺傳修飾之一,迄今為止已經鑒定出170余種修飾方式[1-2]。目前研究最為廣泛的RNA修飾主要包括N6-腺苷酸甲基化(N6-methyladenosine, m6A)、N1-腺苷酸甲基化(N1-methyladenosine, m1A)和5-胞嘧啶甲基化(5-methylcytidine, m5C)等。其中,RNA甲基化m6A修飾是真核生物中含量最為豐富存在最為普遍的RNA修飾[3]。m6A修飾存在于幾乎所有形式的RNA中,包括轉運RNA (transfer RNA, tRNA)、信使RNA (messenger RNA, mRNA)、核糖體RNA (ribosomal RNA, rRNA)以及小RNA (microRNA, miRNA)、長鏈非編碼RNA (long non-coding RNA, lncRNA)和環狀RNA (circular RNA, circRNA)等[4-6]。
m6A修飾最早發現于1974年[7],隨后被證明通過影響RNA的選擇性剪接、穩定性、出核、翻譯和降解等分子功能,從而參與調控多項重要生物學功能[6]。在哺乳動物中,m6A修飾參與調控胚胎干細胞分化[8]、神經系統發育[9-10]和疾病生成[7, 11]等生理過程。在植物中,m6A修飾參與調控植物的生長發育[12-13],植物病毒感染免疫應答[14]等生理過程。在微生物中,m6A修飾影響酵母菌株減數分裂[15]。目前,m6A修飾在昆蟲中的研究主要集中在模式昆蟲果蠅Drosophila中,近年來在經濟昆蟲家蠶Bombyxmori和意大利蜜蜂Apismellifera中也有所研究,在為害水稻和小麥的灰飛虱Laodelphaxstriatellus和寄主十分廣泛的煙粉虱Bemisiatabaci等農業害蟲中也有報道。本文總結了目前m6A修飾在昆蟲生長發育、免疫和抗藥性中的作用,為進一步研究m6A修飾在昆蟲中的生理功能奠定基礎。
1 m6A修飾的分子機制
研究發現,m6A修飾是一種動態可逆的過程[16-17]。在生物體內,甲基轉移酶(writer)可將S-腺苷甲硫氨酸(SAM)上的甲基(-CH3)轉移到RNA分子上m6A基序(RRACH, R=A/G; H=A/U/C)中腺嘌呤(A)的第6位氮原子上,而此甲基又可被去甲基化酶(eraser)移除,起到去甲基化的作用。帶有m6A修飾的RNA分子可被細胞核和細胞質中的識別蛋白(reader)識別,進而影響RNA分子的命運,調控基因的生物學功能[18](圖1)。

圖1 黑腹果蠅中m6A修飾的分子機制Fig.1 Molecular mechanisms of m6A modification in Drosophila melanogaster
1.1 甲基轉移酶
m6A甲基轉移酶是一個保守的多蛋白復合體,可分為2個亞復合體: m6A-METTL復合體(m6A-METTL complex, MAC),即甲基轉移酶3 (methyltransferase-like 3, METTL3)和甲基轉移酶14 (methyltransferase-like 14, METTL14)形成的異源二聚體復合物,以及m6A-METTL相關復合體(m6A-METTL associated complex, MACOM),包括Wilms腫瘤1相關蛋白(WT1 associated protein, WTAP)、KIAA1429(也稱為VIRMA)、RNA結合基序蛋白15 (RNA binding motif protein 15, RBM15)、CCCH型鋅指蛋白13 (zinc finger CCCH-type containing 13, Zc3h13)和E3泛素連接酶(E3 ubiquitin ligases, Hakai)。
METTL3在甲基轉移酶中作為催化亞基與SAM結合,同時將甲基從SAM供體轉移至腺苷的第6位氮原子上。在甲基轉移酶中與METTL3形成異源二聚體的METTL14被證明支持甲基轉移酶復合物與RNA靶點之間的相互作用,并且對METTL3發揮活性至關重要,但是單獨的METTL14并不具有催化作用[19]。WTAP作為調節亞基同樣不具備甲基轉移酶的催化活性,但它與MAC異源二聚體相結合并調控其活性、穩定性以及核定位[20]。除此之外,Schwartz等[21]發現KIAA1429也作為m6A甲基轉移酶復合物的核心組分,其功能與WTAP類似,且對甲基轉移酶復合體起到支架作用。進一步研究發現,RBM15/RBM15B結合m6A甲基化復合體并將其募集到RNA的特定位點,促進甲基化[22]。Zc3h13亞基對于Zc3h13-WTAP-Virilizer-Hakai復合物的細胞核定位至關重要,并可促進m6A甲基化[23]。此外,Hakai也是m6A-METTL相關復合體的組成部分,對于維持MACOM成分的穩定性至關重要[24]。
哺乳動物m6A甲基轉移酶因子METTL3、METTL14、WTAP、Hakai、Zc3h13、RBM15/RBM15B和KIAA1429在果蠅中的同源基因分別為Ime4、METTL14、Fl(2)d、Hakai、Xio(Flacc)、Spenito(Nito)和Virilizer(Vir)[24-33]。
1.2 去甲基化酶
在哺乳動物中,目前發現2個m6A的去甲基化酶基因,分別為脂肪量與肥胖相關蛋白(fat mass and obesity-associated, FTO)和AlkB同系物5 (AlkB homolog 5, ALKBH5)。
在最初的研究中,FTO一直被認為是一種肥胖相關基因,與2型糖尿病和肥胖癥等疾病相關。直到2011年,Jia等在人類肝腫瘤細胞中首次發現FTO可作為m6A修飾的去甲基化酶[16]。FTO屬于非血紅素鐵(Ⅱ)/α-酮戊二酸依賴的雙加氧酶AlkB基因家族,繼FTO之后,Zheng等又在AlkB家族中報道了ALKBH5可作為小鼠的去甲基化酶[34],且發現ALKBH5的去甲基化活性影響mRNA輸出和代謝,進而影響小鼠生育能力等生理過程。除此之外,在擬南芥中,發現ALKBH10B也可發揮去甲基化酶的作用,影響植物的開花性狀、營養生長[35]以及調節擬南芥種子萌發過程中對脫落酸(abscisic acid, ABA)的反應[36]。目前,在昆蟲中發現ALKBH8可能作為其m6A去甲基化酶,但其功能仍處于探索階段[37]。
1.3 識別蛋白
帶有m6A修飾的基序可以被細胞內的多種蛋白質識別并結合,從而調控RNA的加工、翻譯及穩定性等。目前已被發現的識別蛋白,一類是YTH結構域家族蛋白,包括位于細胞質中的YTHDC2、YTHDF1~ YTHDF3以及位于細胞核中的YTHDC1,它們可以通過YTH結構域選擇性識別并結合m6A修飾基序(RRACH, R=A/G; H=A/U/C)。其中,YTHDC1有調節mRNA剪接[38]和加快mRNA核輸出[39]的作用。YTHDC2可以影響靶標mRNA的穩定性及其翻譯效率[40]。YTHDF1能夠增強mRNA的翻譯[41]。YTHDF2能夠通過直接招募CCR4-NOT脫腺苷酸化酶復合體而促使靶標RNA衰減[42]。YTHDF3可以與YTHDF1協同作用進而促進mRNA翻譯;另一方面,YTHDF3可通過YTHDF2促進mRNA的降解[43]。異質性核糖核蛋白(heterogeneous nuclear ribonucleoproteins, HNRNPs)家族也可作為m6A的識別蛋白,包括HNRNPC、HNRNPG和HNRNPA2B1,其中HNRNPA2B1可調控靶標RNA的加工和選擇性剪接[44]。近期研究發現,胰島素樣生長因子2 mRNA結合蛋白(insulin-like growth factor 2 mRNA-binding proteins, IGF2BPs)也被證明可作為m6A識別蛋白。IGF2BPs在正常和應激條件下以m6A依賴的方式促進靶標mRNA的存儲以及其穩定性,從而影響其翻譯效率[45]。
目前在昆蟲中,發現了2種m6A修飾識別蛋白,即在果蠅中發現的YT521-B (CG12076)和YTHDF (CG6422),它們分別是哺乳動物中YTHDC1和YTHDF2的同源蛋白[28]。
2 m6A修飾在昆蟲生長發育、免疫和抗藥性中的作用
2.1 m6A修飾調控果蠅性別決定和劑量補償
研究發現,果蠅性別決定的主調控基因sex-lethal (Sxl)的選擇性剪接調控果蠅的性別,實現雄性果蠅的劑量補償[46-47]。Sxl有兩種不同的剪接形式。雄性果蠅中,外顯子3的終止密碼子處被剪接導致Sxl蛋白提前終止。但雌性果蠅則跳過外顯子3,從而產生功能性Sxl蛋白,保持雌性分化[46-47]。除此之外,在雌性果蠅中,有功能的Sxl蛋白可抑制雄性特異性致死基因(msl-2)的翻譯,使其在雌性個體中沉默,而在雄性個體中,msl-2基因正常表達,促進X染色體上基因轉錄,從而實現雄性個體的劑量補償,因此Sxl基因的選擇性剪接在果蠅中至關重要[47]。而近期研究發現,m6A修飾通過參與調控Sxl基因的選擇性剪接影響果蠅的性別決定和劑量補償[24-33]。
早在20世紀80年代,Fl(2)d[25]和Virilizer[26]就被證明通過調控果蠅Sxl基因的選擇性剪接參與其性別決定。在2015年,研究者通過RNA干擾(RNAi)技術篩選又發現Spenito是性別決定通路中新的組成部分[27]。隨著研究的不斷深入,發現Fl(2)d、Virilizer和Spenito都是m6A甲基轉移酶復合體的一部分,且共同調控果蠅Sxl基因的選擇性剪接,這表明m6A修飾可能參與調控性別決定。進一步研究發現,識別蛋白YT521-B可識別Sxl上的m6A修飾,介導Sxl基因的選擇性剪接從而調控果蠅性別決定和劑量補償[28-30],但Sxl上具體的m6A位點仍不清楚。隨后,Xio被證明是m6A甲基轉移酶復合體中的成員并且通過調控Sxl基因的選擇性剪接參與果蠅的性別決定[31-32]。近期研究發現Hakai也是m6A甲基轉移酶復合體中的成員,同樣也參與調控果蠅Sxl基因的選擇性剪接[24, 33]。與此同時,通過RNA甲基化免疫共沉淀測序(MeRIP-seq)發現,Sxl基因外顯子3附近存在3個m6A位點參與調控Sxl的選擇性剪接[33]。基于以上研究,可以總結出m6A修飾參與調控Sxl選擇性剪接進而調控果蠅性別決定和劑量補償的作用模型:在雌性果蠅中,Sxl基因外顯子3附近的m6A位點被m6A甲基轉移酶復合物特異性識別并甲基化,同時由于這些位點非常接近于外顯子與內含子的連接區域,因此m6A識別蛋白YT521-B會特異性識別并結合到這些m6A位點,從而干擾剪接機制,最終導致外顯子3被跳過,使得在雌性果蠅中可以翻譯成完整的Sxl蛋白(圖2)。
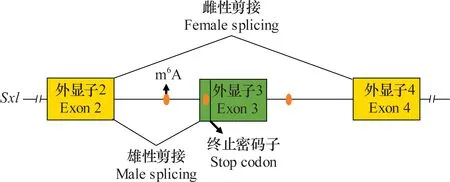
圖2 m6A修飾調控Sxl基因的選擇性剪接Fig.2 The m6A modification regulates alternative splicing of Sxl gene
2.2 m6A修飾調控果蠅神經發育和功能
研究發現,在果蠅胚胎發育過程中,m6A甲基轉移酶亞基Ime4、METTL14、Fl(2)d和Virilizer大量富集于神經外胚層,表明m6A修飾可能參與調控果蠅的神經系統發育。雖然m6A水平在果蠅成蟲階段總體有所下降,但在頭部仍然顯著高表達,且當Ime4和METTL14被敲除后,果蠅的飛行和移動行為受到嚴重影響,但這樣的影響可以通過表達神經元(elav-GAL4)所挽救,表明m6A修飾通過影響神經元功能來調控果蠅的行為[28]。除此之外,研究發現18S核糖體RNA 的甲基轉移酶METTL5可能通過調控核糖體RNA的m6A修飾從而影響果蠅神經系統,進一步影響果蠅的移動和辨別方向等行為[48]。進一步研究表明,果蠅中m6A修飾的識別蛋白YTHDF參與調控神經系統發育,YTHDF可以與脆性X智力遲鈍1(fragile X mental retardation type 1,Fmr1)基因相互作用,且抑制關鍵靶標基因的翻譯,以確保適當的軸突生長和穩態,從而使神經系統正常發育[49]。m6A修飾調控果蠅神經功能不僅僅體現在果蠅的移動和辨別方向等行為,通過科學家們深入研究發現,m6A/YTHDF通路參與調控蘑菇體(mushroom body, MB)(聯想學習中心)的神經元,從而介導果蠅對氣味的識別記憶[50],這與哺乳動物中m6A修飾的功能相似,在哺乳動物中已經發現m6A修飾影響神經功能和行為,其中包括了學習和記憶機制[9-10, 51-52]。
2.3 m6A修飾可能調控家蠶應對核型多角體病毒(BmNPV)感染
家蠶核型多角體病毒(Bombyxmorinucleopolyhedrovirus, BmNPV)病是家蠶養殖過程中最常見且危害最嚴重的病毒。BmNPV侵入后首先感染家蠶中腸組織,使其體節腫脹,體色乳白,體壁易破。近期研究發現m6A修飾參與調控家蠶應對核型多角體病毒的感染[53-54]。Zhang等[54]對感染病毒和未感染病毒的家蠶中腸進行了m6A組學分析,發現感染與未感染病毒的家蠶之間有1 221個差異表達的m6A峰,表明m6A修飾可能參與了家蠶應對病毒感染的過程。除此之外,作者發現當使用siRNA技術沉默家蠶細胞中的2個甲基化酶核心亞基BmMETTL3和BmMETTL14以及家蠶識別蛋白BmYTHDF3基因后,家蠶中病毒結構蛋白VP39表達增加,而當過表達這3個基因后,情況相反[54]。他們在進一步的研究中發現與病毒復制和增殖相關的病毒極早期基因ie-1具有高水平m6A修飾,且此基因的m6A修飾負向調控了IE-1蛋白表達。除此之外,作者發現病毒復制在過表達BmYTHDF3的細胞中被顯著抑制,且呈劑量依賴性,而在轉染si-BmYTHDF3的細胞中則發現相反的作用[55]。但在另一項研究中,Xue等發現在家蠶中腸中,低水平的METTL3可以抑制核型多角體病毒的增殖。在家蠶細胞中,當METTL3表達水平降低抑制了BmNPV核衣殼蛋白基因VP39和包膜融合蛋白基因GP64的表達,說明低水平的m6A可能在一定程度上抑制BmNPV的增殖[56]。綜上所述,m6A修飾參與調控家蠶應對家蠶核型多角體病毒的過程,但其作用機制十分復雜,因此還需要更多的研究去探索m6A修飾是如何具體參與這一過程的。
2.4 m6A修飾可能調控家蠶滯育
昆蟲滯育是受環境條件的誘導發生的暫停發育的現象,是為了保障其在不利的環境中生存而進化出的特殊生活習性和生理反應。家蠶通過滯育可以調節生長和繁殖,提高抗應激能力,從而維持整個種群生存[57]。Jiang等發現與非滯育家蠶種群相比,滯育種群中m6A修飾相關基因表達和m6A修飾含量更高,這表明m6A修飾可能在家蠶滯育中起著關鍵作用[58],但其具體功能尚不清楚。
2.5 m6A修飾可能調控蜜蜂級型分化
蜜蜂的群居性表現出嚴格的勞動分工,這很大程度上是基于級型分化。級型分化是指蜜蜂的雌性低齡幼蟲可以通過不同營養攝入控制幼蟲發育成蜂王或者工蜂[59-60]。通過對3齡階段的蜂王和工蜂幼蟲進行m6A甲基化測序分析發現,蜜蜂幼蟲的m6A甲基化水平可因營養攝入不同而發生變化,測序結果顯示,工蜂幼蟲比蜂王幼蟲含有更多的m6A高甲基化峰。除此之外,在工蜂和蜂王之間,許多與級型分化相關轉錄本的m6A水平存在顯著差異。值得注意的是,當使用甲基化抑制劑3-脫氮基腺苷(3-deazaadenosine, 3-DAA)抑制工蜂幼蟲的m6A修飾水平時,工蜂幼蟲表現出蜂王幼蟲特征,進一步證明了m6A修飾在蜜蜂的級型分化中起到了重要作用[61]。
2.6 m6A修飾調控煙粉虱殺蟲劑抗性
煙粉虱是一種全球分布,且破壞性極強的超級農業害蟲,其寄主范圍十分廣泛,嚴重危害蔬菜和糧食作物生產[62-64]。新煙堿類殺蟲劑是防治此害蟲的有效手段,然而隨著田間大量及不合理地使用此類殺蟲劑,煙粉虱對這類殺蟲劑已產生了嚴重的抗藥性[65]。Yang等首次報道了細胞色素P450基因CYP4C64的表達量升高是導致煙粉虱對噻蟲嗪產生抗性的主要原因之一[66]。CYP4C64基因序列的分析結果表明,抗性煙粉虱CYP4C64基因的5′-UTR區有一個T-206A突變,突變后的腺嘌呤位點可被甲基轉移酶識別并進行甲基化修飾,即該突變引入了一個m6A修飾位點,通過進一步研究發現此m6A修飾位點可促進煙粉虱CYP4C64基因的表達,從而導致煙粉虱對噻蟲嗪產生抗藥性[66](圖3)。
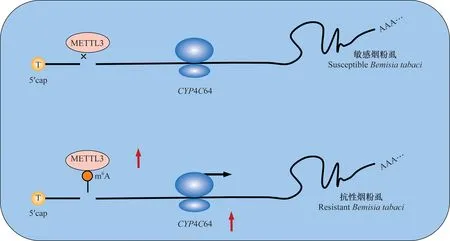
圖3 m6A修飾介導煙粉虱CYP4C64基因的表達Fig.3 m6A-mediated regulation of CYP4C64 expression in Bemisia tabaci
2.7 m6A修飾調控灰飛虱應對水稻黑條紋矮縮病毒(RBSDV)感染
在灰飛虱中,Tian等發現當灰飛虱中腸組織攜帶水稻黑條紋矮縮病毒(rice black-streaked dwarf virus, RBSDV)后會導致灰飛虱的m6A水平下降,而干擾灰飛虱中m6A甲基轉移酶基因(LsMETTL3和LsMETTL14)的表達后,灰飛虱中腸細胞中該病毒的積累水平顯著升高,表明m6A水平與病毒復制呈負相關,即m6A修飾可以限制病毒的復制,同時病毒也可以抑制灰飛虱的m6A修飾達到其持久性傳播的目的[67]。
3 總結與展望
近年來,隨著RNA甲基化免疫沉淀測序等方法的應用,人們對RNA修飾生物學的興趣和認識有了快速的發展[68]。m6A修飾目前已被證明調控哺乳動物、植物以及微生物的多項生理過程。而在昆蟲中,m6A修飾的研究主要集中在模式昆蟲果蠅和家蠶中。近年來在經濟昆蟲家蠶和蜜蜂,以及在為害水稻、小麥的灰飛虱和寄主十分廣泛的煙粉虱中也有報道。
目前,m6A修飾對于果蠅性別決定和劑量補償[24-33]以及神經發育和神經功能[28, 49-50]兩方面的影響機制已經較為明確。但由于m6A修飾是一種動態可逆的過程,當前在昆蟲中潛在的去甲基化酶ALKBH8已有相關研究報道,但針對昆蟲去甲基化酶如何參與m6A調控及對其各項功能的影響還有待進一步明確[37]。
與此同時,m6A修飾也調控了其他昆蟲的部分生理過程。研究發現,m6A修飾在家蠶滯育中發揮作用[57],并且調控家蠶應對家蠶核型多角體病毒(BmNPV)的侵染[54-56]。在蜜蜂中,發現m6A修飾調控了蜜蜂的級型分化[61]。在煙粉虱中,發現m6A修飾調控其對殺蟲劑的抗性[66]。在灰飛虱中,發現其中腸中的m6A水平與中腸細胞中水稻黑條紋矮縮病毒(RBSDV)復制能力呈負相關[67]。但m6A修飾的功能在這些昆蟲中的研究中還處于起始階段,系統的調控作用機制有待進一步研究。此外,m6A除了調控以上昆蟲的各項生理功能,在其他昆蟲中的研究仍處于空白狀態,還有待研究。本文系統總結了目前m6A修飾在昆蟲生長發育、免疫和抗藥性中的作用,為今后研究基于m6A的表觀遺傳修飾對昆蟲各項生理機能的調控作用提供了新的思路。
昆蟲與人類的關系密不可分,對人類而言,昆蟲可以分為害蟲和益蟲。首先是一部分昆蟲作為害蟲對人類農業生產的危害,據聯合國糧農組織的數據顯示,全球每年將近40%的農作物在生產過程中受到害蟲為害,造成的經濟損失達4 466億元。如農業害蟲果蠅為害黑莓、藍莓、櫻桃、覆盆子和草莓等250余種栽培果蔬作物[69]。煙粉虱為害番茄、辣椒、十字花科蔬菜及棉花等600多種植物[62-64]。灰飛虱主要為害水稻、玉米等糧食作物[67]。其次是另一部分昆蟲作為益蟲對人類農業生產的益處。如經濟昆蟲家蠶吐出的蠶絲對人類紡織工業至關重要[70],蜜蜂取食花蜜花粉促進植物傳粉受精而有益于農作物結實[71]。未來需要進一步研究m6A修飾在昆蟲中的作用機制以及功能。如此,一方面可以更深入地了解昆蟲各項生理功能,有助于針對果蠅、煙粉虱等害蟲發現新的害蟲防治靶標、制定新的防治策略,期望通過綠色環保的生物防治手段降低害蟲對農作物的危害。另一方面,有助于更好地改造并培育家蠶、蜜蜂等經濟昆蟲的新品種,從而服務于人類農業生產。
總而言之,隨著研究技術的不斷發展,哺乳動物與植物中對于m6A的調控機制以及功能的研究已經取得了較為豐碩的進展。但在昆蟲中,m6A的相關研究總體而言還處于起始階段,未來需要進一步研究m6A修飾在昆蟲中的作用機制以及各項功能,為更有效的害蟲生物防治以及經濟昆蟲培育等生產實踐提供堅實的研究基礎。
