鹽脅迫對裸果木幼苗生長和葉片抗逆生理指標的影響
2023-10-17 14:05:56周曉瑾黃海霞齊建偉羅永忠
西北植物學報 2023年9期
關鍵詞:植物
周曉瑾,黃海霞,張 婷,齊建偉,羅永忠
(甘肅農業大學 林學院,蘭州 730070)
鹽漬化是引起土地荒漠化和土地退化的主要原因,由于全球氣候變暖和環境惡化,以及不合理灌溉、耕作活動,鹽漬土地面積呈逐年擴大的趨勢,土壤鹽漬化已成為全球性的生態和資源問題[1]。據聯合國糧農組織(FAO)統計,目前全球鹽漬化土地面積約有10億 hm2,中國現有各類鹽漬土面積約9 913萬 hm2,占全國可耕地面積的25%,特別是西北地區鹽漬土所占比例較大,其中甘肅鹽漬化土壤面積約有141.3萬 hm2,約占鹽漬化耕地面積的22.81%[2]。因此,改良并利用鹽漬土及修復農業生態環境在目前農業生產中顯得尤為重要。推廣和選育耐鹽植物是改良和利用鹽漬土壤有效措施之一[3]。研究發現,當土壤發生鹽漬化后,首先會引起植物遭受滲透脅迫,然后造成離子失衡,繼而導致離子毒害,損害植株的葉綠體色素系統,抑制Rubisco酶活性,使葉綠素含量降低,進而抑制植物的光合作用[4-5]。鹽脅迫還會造成活性氧(ROS)大量積累,導致膜脂過氧化,改變細胞膜透性,引起生理代謝紊亂及有毒物質積累,從而影響植物的正常生長及形態建成[6]。研究表明,鹽脅迫下可溶性小分子物質與植株的滲透調節密切相關,滲透調節是植物適應鹽脅迫的主要生理機制[7,8]。植物還可通過調節抗氧化系統清除ROS積累的方式減輕或抵御鹽脅迫對細胞的傷害[9]。
裸果木(Gymnocarposprzewalskii)為石竹科裸果木屬亞灌木狀植物,主要分布在干旱灰棕色荒漠土或棕色荒漠土的礫石戈壁或低矮的剝蝕殘丘下部[10],為古地中海區旱生植物區系成分,是鹽生環境中最古老的類群,對研究中國西北、內蒙古荒漠的發生、發展、氣候的變化以及旱生植物區系成分的起源有重要的科學價值[11]。由于生存條件惡劣,繁殖困難,又常遭樵采和駱駝啃食,導致其分布區已日益縮小。因此,其日益受到越來越多的學者關注。目前,國內外學者對裸果木種子萌發[12]、種群結構[13-14]、形態解剖學[15]、氣候變化與系統地理結構[16]及抗旱生理特性[17-18]等方面的研究較多,而鹽脅迫對裸果木生理特性影響方面的系統研究報道較少。研究表明,100 mmol/L以下的NaCl處理對裸果木種子萌發率的影響不顯著,萌發率可達到85%以上[12];在低濃度鹽脅迫(NaCl≤0.4%)下,裸果木幼苗葉片凈光合速率下降不明顯,瞬時水分利用效率有所提高[19]。這些研究結果說明裸果木幼苗具有一定的耐鹽性,也存在耐鹽閾值,但在NaCl脅迫下,裸果木幼苗的滲透調節和抗氧化響應能力如何,迄今尚不清楚。因此,本研究以一年生裸果木為研究對象,設置不同NaCl濃度梯度模擬不同程度鹽脅迫,通過控制性實驗分析裸果木幼苗的生長指標及葉片滲透調節物質、抗氧化指標及光合色素含量的變化規律及差異性,探究裸果木對鹽脅迫的生理響應機制及其耐鹽性,以期為進一步研究裸果木耐鹽性及人工種植地選擇提供一定的理論依據。
1 材料和方法
1.1 試驗材料
試驗于2021年4-8月在甘肅農業大學實驗室和甘肅農業大學試驗基地大棚內進行,裸果木種子采集于甘肅安西極旱荒漠國家級自然保護區植物園(E95°44′12″,N40°29′53″)。
1.2 材料培養與處理
于2021年4月采用花盆進行播種育苗,播種前先對種子和基質進行處理,種子先用10%的次氯酸鈉溶液浸泡3 min,再用蒸餾水沖洗干凈,最后用濾紙吸干水分。育苗基質由泥炭土和蛭石按照3∶1比例配制而成,經自然風干后裝入花盆(直徑17.8 cm,高度15 cm),每盆裝入等量的基質(360.0 g),并按照設置的基質鹽分處理和盆土干重,計算每個處理需要加入NaCl的量,分別配制成1 L的溶液,緩慢加入到基質中;每盆播種3穴,每穴4粒,深度為1 cm,待成活穩定后,每盆保留3株,于2021年5月中旬移至甘肅農業大學試驗基地的塑料大棚內進行日常管護。試驗共設5個處理:以蒸餾水灌溉為對照(CK),并設質量分數分別為0.4%、0.8%、1.2%和1.6% NaCl處理,每個處理3次重復(每重復4盆),每3~5 d澆等量水,若底部托盤中有滲出液則澆回盆中,以避免土壤鹽分的流失。8月中旬結束控制實驗,在各處理每個重復中隨機取6株,測定完株高、基徑后采集完整植株用于測定各器官干重,并采集其余植株的完全展開葉片,立即進行液氮速凍,放入-80 ℃超低溫冰箱,用于測定生理指標。
1.3 測定指標與方法
1.3.1 生長指標測量植株基部到生長點的長度為株高;測量地面根頸處的植株直徑為基徑。然后將每個處理植株各部分器官分開,用烘箱先調節至105 ℃殺青30 min,然后75 ℃下烘干至恒重,自然冷卻后,用天平(精度0.001 g)稱其根、莖、葉干重,并計算根冠比。[根生物量/(莖生物量+葉生物量)]。
1.3.2 葉片葉綠素含量
取各處理的裸果木鮮葉0.3 g,采用乙醇提取法[14]測定葉片葉綠素含量,每處理測定3個重復。
1.3.3 葉片滲透調節物質含量
葉片滲透調節物質含量參照高俊鳳[20]方法測定。取不同處理的裸果木鮮葉0.3 g分別放入3支具塞試管中,加入10 mL蒸餾水,在100 ℃水浴下煮沸提取2次各30 min,定容至25 mL刻度容量瓶中,采用蒽酮法測定可溶性糖含量。取不同處理的裸果木鮮葉0.3 g,加入少許石英砂和蒸餾水在預冷的研缽中研磨成勻漿后倒入10 mL容量瓶中,在10 000 r/min離心10 min,上清液即為可溶性蛋白提取液,其含量采用考馬斯亮藍G-250染色法測定。取不同處理的裸果木鮮葉0.5 g,剪碎混勻置于具塞試管中,加入10 mL 3%的磺基水楊酸溶液,在沸水浴中提取10 min(提取過程中要經常搖動),上清液即為脯氨酸的提取液,其含量采用磺基水楊酸提取法測定。
1.3.4 葉片相對電導率
采用電導儀法測定[21]。取各處理的裸果木鮮葉0.3 g,加10 mL純水,用真空泵抽氣15 min左右,抽出間隙中的空氣后,再重新緩緩放入空氣,在室溫保持30 min,用DDS-11A型電導儀測定各處理電導率,然后在沸水浴中保持5 min,自然冷卻后再用電導儀測定煮沸后電導率,各處理重復3次。
1.3.5 葉片丙二醛含量
取不同處理裸果木鮮葉0.3 g樣品于研缽,加5%三氯乙酸2 mL,研磨后,將勻漿倒入離心管,用三氯乙酸定容至5 mL,在3 000 r/min下離心10 min,取上清液即為丙二醛提取液,采用硫代巴比妥酸法測定丙二醛含量[20]。
1.3.6 葉片抗氧化酶活性
葉片抗氧化酶活性參照高俊鳳[20]方法測定。取不同處理裸果木鮮葉0.2 g,置于預冷的研缽中,加入50 mmol/L pH為7.8的磷酸緩沖液,研磨成勻漿定容,在4 ℃條件下10 000 r/min離心20 min,上清液即為酶粗測定液。分別采用氮藍四唑光還原法、愈創木酚法、紫外吸收法測定超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)活性。
1.4 數據分析
采用Microsoft Excel 2010整理數據;采用 SPSS 26.1軟件進行方差分析(ANOVA),采用Duncan法檢驗處理間差異的顯著性;采用Origin 2018作圖。
2 結果與分析
2.1 鹽脅迫對裸果木幼苗生長的影響
表1顯示,隨著鹽脅迫濃度的增加,裸果木幼苗株高、基徑和干重均逐漸降低,而其根冠比逐漸增加。其中,裸果木幼苗株高、基徑和根干重在0.4% NaCl處理下比對照略有降低,而在0.8% NaCl處理下則比對照分別顯著降低45.8%、53.3%和33.3%,表明裸果木幼苗生長受到了高鹽脅迫(0.8%、1.2%和1.6%)的顯著抑制;同時,裸果木幼苗莖干重、葉干重和總干重均表現出相同的變化趨勢,即在各濃度NaCl脅迫處理下均顯著低于對照,且1.6% NaCl處理下比對照分別顯著降低80.9%、90.9%和76.3%;另外,裸果木幼苗根冠比則在各濃度NaCl處理下均比對照顯著升高,在1.6%鹽脅迫下達到對照的2.5倍。
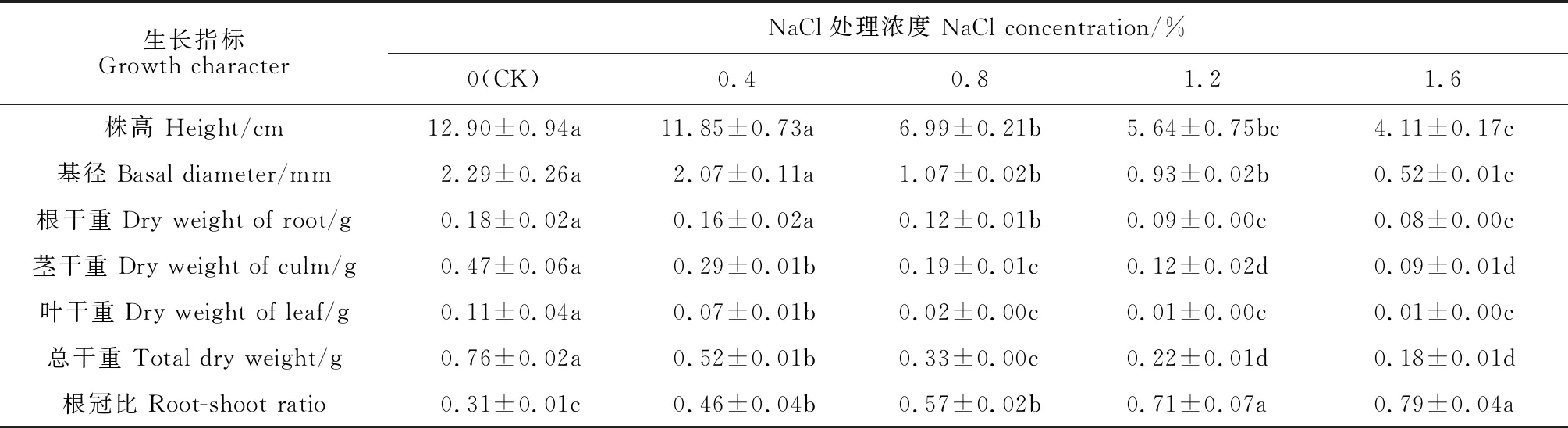
表1 鹽脅迫下裸果木幼苗形態生長指標的變化
總體來看,鹽脅迫嚴重抑制了裸果木幼苗的形態生長,且NaCl處理濃度越高抑制程度越大;鹽脅迫顯著限制了裸果木幼苗總生物量的積累,且對葉、莖生長的限制作用大于根系。
2.2 鹽脅迫對裸果木幼苗葉片滲透調節物質含量的影響
從圖1可知,隨著NaCl脅迫濃度的增加,裸果木幼苗葉片可溶性糖(SS)和可溶性蛋白(SP)含量均呈現先上升后下降趨勢,并均在0.4% NaCl處理下達到最大值,且各脅迫處理均顯著高于對照水平;同時相應的葉片脯氨酸(Pro)含量則表現出逐漸大幅度增加的趨勢,且除0.4% NaCl處理外也均與對照差異顯著。其中,裸果木幼苗葉片SS和SP含量在0.4% NaCl處理下均達到最大值,較對照分別顯著增加了70.2%和136.8%;當鹽濃度超過0.4%時,葉片SS和SP含量則逐漸下降,但仍均顯著高于對照,增幅分別為23.0%~33.2%和32.0%~75.1%,0.8%和1.2% NaCl處理之間差異均不顯著;裸果木幼苗葉片Pro含量在0.8%、1.2%和1.6% NaCl處理下較對照分別顯著提高116.9%、126.0%和169.3%,而0.8%與1.2% NaCl處理間無顯著差異。以上結果說明,裸果木幼苗在0.4% NaCl處理(低鹽脅迫)下主要積累可溶性糖和可溶性蛋白,在0.8%和1.2% NaCl處理(中高鹽脅迫)下主動顯著增加脯氨酸積累來彌補可溶性糖和可溶性蛋白積累的不足,而在1.6% NaCl處理時(高鹽脅迫),進一步主動大幅增加脯氨酸積累,三者相互協作來共同發揮滲透調節作用,表現出一定的補償效應。
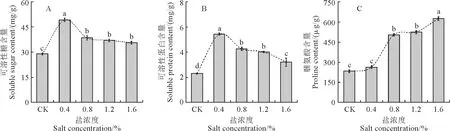
不同小寫字母表示處理間在0.05水平存在顯著性差異。下同。
2.3 鹽脅迫對裸果木幼苗葉片丙二醛含量和相對電導率的影響
由圖2可知,裸果木幼苗葉片丙二醛(MDA)含量和相對電導率(REC)均隨著NaCl處理濃度的增加呈逐漸升高趨勢。其中,裸果木幼苗葉片MDA含量在0.4% NaCl處理下僅比對照稍高,而在0.8%~1.6% NaCl處理下比對照顯著升高了56.8%~122.1%,且0.8%和1.2%兩個NaCl處理之間差異不顯著;在0.4%~1.6% NaCl處理下,裸果木幼苗葉片REC比對照顯著上升了24.1%~59.9%,而1.2%和1.6% NaCl處理之間差異不顯著性。以上結果表明,當NaCl處理濃度超過0.4%時,裸果木葉片膜系統受到了明顯的過氧化損傷,造成電解質大量外滲。

圖2 鹽脅迫下裸果木幼苗葉片丙二醛含量及相對電導率的變化
2.4 鹽脅迫對裸果木幼苗葉片抗氧化酶活性的影響
裸果木幼苗葉片超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)活性在各濃度NaCl脅迫下均比對照不同程度提高,且增幅大多達到顯著水平(圖3)。其中,隨著NaCl處理濃度的增加,葉片SOD和POD活性均呈先升高后下降的變化趨勢,它們分別在0.8%和0.4% NaCl處理下達到最大值,分別較對照顯著升高了43.4%和45.9%(圖3,A、B);而葉片CAT活性則隨著NaCl處理濃度的增大而逐漸升高,且都與對照差異達到顯著水平,增幅在57.6%~195.1%之間(圖3,C)。以上結果表明,裸果木幼苗葉片POD和CAT活性在各濃度鹽脅迫下均顯著增強,而其SOD活性在鹽濃度超過0.4%時也顯著增強,均對清除ROS發揮了顯著作用,有效緩解自由基對膜系統的損傷,且CAT活性增強的幅度更大。
2.5 鹽脅迫對裸果木幼苗葉片葉綠素含量的影響
隨著NaCl處理濃度的增加,裸果木幼苗葉片葉綠素a、葉綠素b、類胡蘿卜素和總葉綠素含量均逐漸降低,但在0.4% NaCl處理下比對照降低均不顯著,在0.8%~1.6% NaCl處理下均比對照顯著降低,降幅分別為45.8%~54.2%、67.1%~94.9%、74.3%~82.6%和54.0%~70.0%(表2)。由此可見,各濃度NaCl處理均抑制了裸果木幼苗葉片光合色素的合成,導致光合色素含量均不同程度降低,且NaCl處理濃度越高下降幅度越大。
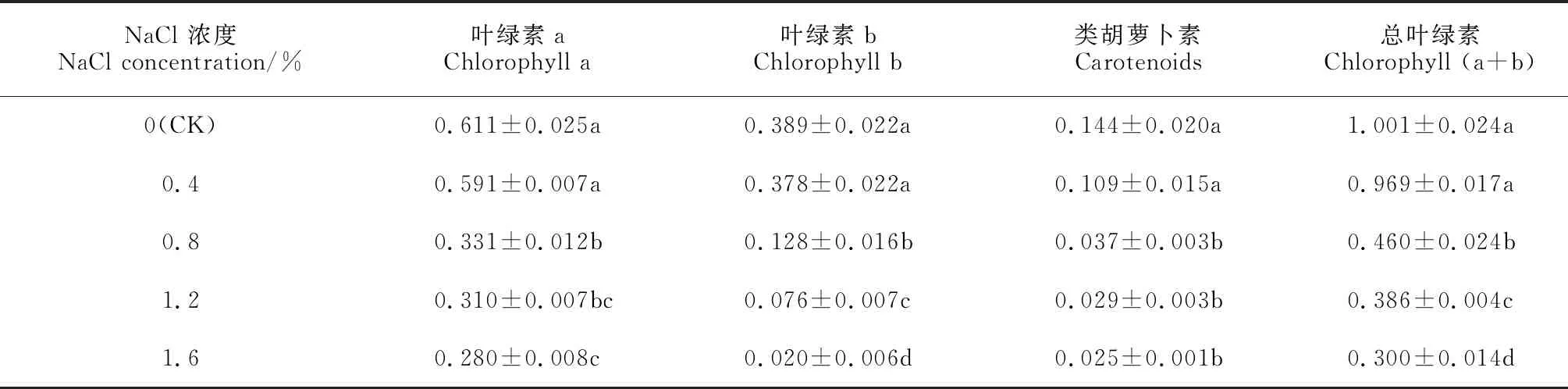
表2 鹽脅迫下裸果木幼苗葉片葉綠素含量的變化
3 討 論
鹽脅迫對植物的傷害作用主要包括滲透脅迫、離子毒害和營養失衡等方面,但最終體現在植物生長抑制,根冠比增加,甚至導致死亡[22]。生長量和植株形態作為評估鹽脅迫程度和植物抗性的最直接指標[23],相關研究表明,在鹽脅迫條件下,藜麥幼苗根部和莖部生長均受到抑制,且NaCl處理濃度越高,受抑制現象越明顯[24]。本試驗中低鹽脅迫對裸果木生長影響較小,但當NaCl處理超過0.4%時,根、莖、葉干重以及株高、基徑均顯著降低,同時植株矮小、葉色變淡。這表明裸果木具有一定的耐鹽性。
葉綠素是植物進行光合作用的主要化學物質,其含量多少能夠直接反映植物葉片光合代謝能力的強弱[25]。鹽脅迫環境下,植物葉綠體結構受損,葉綠素合成與降解失衡,從而使其光合作用受到抑制。本研究結果發現,裸果木幼苗葉片葉綠素a、葉綠素b、類胡蘿卜素和總葉綠素含量在鹽脅迫下均明顯下降,且隨著鹽脅迫程度的加重,其含量下降幅度越大,這可能是由于較高濃度的鹽脅迫破壞了土壤中的離子穩態,使得與葉綠素合成有關的離子(Mg2+、Cu2+、Fe2+)沉淀,進而減緩了葉綠體對光能的吸收能力,降低了色素與色素蛋白之間的親合能力,使類囊體膜破壞,最終導致葉綠素含量降低[26-28]。類胡蘿卜素既是光合色素又是細胞內源抗氧化劑,既可以吸收部分光能,又能吸收PSⅡ的過剩能量,在一定程度上可以保護葉綠體[29],本研究中類胡蘿卜素含量下降可能是鹽脅迫下產生的活性氧消耗了類胡蘿卜素[5],具體的機理有待進一步研究。
鹽脅迫環境條件下,植物細胞會主動積累一些滲透調節物質,維持高的細胞質滲透壓,以保障植物細胞能抵御鹽脅迫[30],可溶性糖、可溶性蛋白和脯氨酸是植物體內主要滲透調節物質,這些物質的積累便于植物在鹽分環境下對水分的吸收,從而保證植物的正常生理活動[31-33]。在本研究中,NaCl濃度低于0.4%時,裸果木幼苗葉片SS和SP含量顯著高于對照,說明葉片通過積累大量的SS和SP來抵御NaCl脅迫對裸果木的傷害;但隨著NaCl脅迫的加重, SS和SP含量有所下降,這可能是由于細胞內葉綠素含量下降,光合速率減弱,合成物減少而導致SS含量下降和SP合成受限,本研究與羅達等[34]的研究結果相似。本研究中NaCl處理濃度超過0.4%時,裸果木幼苗葉片Pro含量顯著提高,且在1.6% NaCl處理下達到最大值,表明高鹽環境能顯著誘導Pro的滲透調節作用,曹齊衛等[35]的研究也得出了類似的結論。從鹽脅迫下3種滲透調節物質的變化規律來看,Pro與SS和SP的積累進程存在補償作用。
植物細胞膜主要起著調節和控制細胞內外物質交換和運輸的作用,主要有磷脂和蛋白質組成,具有選擇通透性[36]。鹽脅迫導致細胞膜結構受損,膜透性增大,細胞內的電解質外滲,繼而導致電導率升高。因此,相對電導率可表征植物遭受鹽脅迫傷害的程度,即相對電導率越大,細胞膜透性越大[37]。此外,鹽脅迫還會引起植物的脂質過氧化作用,其中MDA作為膜脂過氧化反應的主要產物之一,其含量的多少在一定程度上能夠反映出膜損傷程度的大小[38-39]。本研究中,裸果木幼苗MDA含量隨著NaCl脅迫的加重而逐漸上升,但其在0.4% NaCl處理下與對照相比增加較少且含量較低;而當NaCl處理高于0.8%時,MDA含量增加顯著,這說明當鹽濃度較低時,裸果木葉片細胞膜傷害的程度小,隨著鹽濃度的增加,膜結構嚴重受損。

4 結 論
本研究通過對裸果木在NaCl脅迫下的生長狀況、滲透調節物質、膜系統的穩定性、抗氧化酶系統和葉綠素含量的分析可知,在0.4%NaCl處理下,裸果木幼苗的株高、基徑、根干重、膜脂過氧化程度和光合色素含量變化不明顯,可溶性糖和可溶性蛋白顯著積累,POD和CAT活性顯著增強,對鹽脅迫表現出一定的耐受能力。當NaCl處理濃度≥0.8%時,裸果木幼苗葉片葉綠素含量顯著下降,膜脂過氧化作用明顯加劇,膜透性顯著增大,可溶性糖和可溶性蛋白含量降低,抗氧化酶活性減弱,干物質的合成與積累受到顯著抑制。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
