靈孢多糖通過抑制TLR4/NF-κB通路改善EAE小鼠的髓鞘脫失
2023-10-22 05:51:10李彥青楊智超宋麗娟王翰斌楊立志劉建春肖保國馬存根
中國藥理學通報 2023年10期
李彥青,王 青,楊智超,宋麗娟,王翰斌,楊立志,劉建春,肖保國,馬存根
(1. 山西中醫藥大學神經生物學研究中心,國家中醫藥管理局多發性硬化益氣活血重點研究室,山西 晉中 030619;2. 山西國潤制藥有限公司,山西 大同 038100;3. 復旦大學附屬華山醫院神經病研究所,上海 200025)
多發性硬化(multiple sclerosis,MS)是一種慢性免疫介導的中樞神經系統(central nervous system,CNS)疾病,通常以神經炎癥、白質脫髓鞘、膠質細胞增生和血腦屏障損傷等為特征[1]。MS主要發生在20~40歲,是導致年輕人非創傷性殘疾的主要原因[2]。雖然MS的致病機制尚不清楚,但T細胞、B細胞和其他免疫細胞似乎介導CNS的免疫應答。目前的臨床療法大多聚焦于通過發揮免疫抑制或免疫調節作用,在一定程度上降低復發率[3]。然而,目前的藥物僅對部分MS患者有效,對繼發性進展性MS幾乎沒有作用。因此,迫切需要開發新型有效的治療藥物。
用髓磷脂少突膠質細胞糖蛋白(myelin oligodendrocyte glycoprotein,MOG)免疫C57BL / 6小鼠誘導的實驗性自身免疫性腦脊髓炎(experimental autoimmune encephalomyelitis,EAE)是研究MS的理想模型,其通過誘導CD4+T細胞分化為效應T細胞并遷移至CNS引發一系列的炎癥反應,目前其參與CNS炎癥及免疫反應的調節機制尚不清楚[4]。相關研究發現,Toll樣受體4(Toll-like receptor 4,TLR4)在MS患者和EAE小鼠脊髓中上調,并且會激活下游核因子κB(nuclear factor kappa-B,NF-κB)的表達,產生更高水平的促炎細胞因子,加重髓鞘脫失和炎性浸潤導致的神經損傷[5]。因此,通過抑制TLR4/NF-κB信號通路可能是改善EAE進展的有效途徑。
靈芝是中國著名的中草藥,在傳統中醫中已有超過1000年的使用歷史,已被用于治療各種疾病并促進健康和長壽。靈孢多糖(ganoderma lucidum polysaccharides,GLPS)是靈芝水溶性提取物中的主要生物活性成分,具有免疫調節、抗血管生成和抗氧化以及抗炎的作用[6]。先前的研究表明,GLPS能夠通過減弱小膠質細胞介導的神經炎癥并且增強有益的吞噬作用來發揮神經保護作用,從而治療阿爾茨海默病(Alzheimer′s disease,AD)和帕金森病(Parkinson′s disease,PD)等神經退行性疾病[7]。然而,目前尚不清楚GLPS對EAE是否有治療作用。因此,本研究擬采用GLPS作用于EAE小鼠模型觀察其對髓鞘脫失及炎性浸潤的改善作用,并初步探討其作用機制及其與TLR4/NF-κB信號通路的相關性。
1 材料與方法
1.1 材料
1.1.1實驗動物 6~8周齡C57BL/6雄性小鼠(體質量18~20 g,SPF級)共30只,購買于北京維通利華實驗動物技術有限公司,生產許可證號:SYXK(京)2022-0052。小鼠于山西中醫藥大學動物房飼養,飼養條件為:溫度(20±2)℃,濕度40%~50%,晝夜交替規律,自由取食飲水。所有動物實驗均符合動物實驗倫理委員會的規定和要求。
1.1.2試劑 MOG35-55(國泰生物);結核分枝桿菌H37Ra(tuberculosis myco-bacterium,TB,BD Bioscience);百日咳毒素(pertussis toxin,PTX,Sigma);完全弗氏佐劑(complete Freund's adjuvant,CFA,Alexis);GLPS注射液(山西國潤制藥有限公司,H20003510);固藍染色試劑盒(luxol fast blue,LFB,Solarbio,G3245);蘇木素-伊紅染色試劑盒(hematoxylin-eosin staining,HE,Solarbio,G1120);流式抗體干擾素-γ(interferon-gamma,IFN-γ)、白細胞介素17(interleukin 17,IL-17)、叉頭樣轉錄因子3(forkhead transcription factor 3,Foxp3)均購買于Thermo Fisher;TLR4(proteintech,19811-1-AP);MBP(Abcam,EPR21188);MyD88(Abclonal,A0980);NF-κB(Cell Signaling Technology,D14E12);p-NF-κB(Cell Signaling Technology,93H1);β-actin(Bioworld,AP0060);HRP標記羊抗兔二抗(Boster,BA1055);RIPA裂解液(Servicebio,G2002)。
1.1.3儀器 熒光顯微鏡(Leica,DM40008/DFC450C);流式細胞儀(BD Bioscience,Accuri C6);電泳儀(Bio-Rad,165-8004);低溫型研磨儀(Servicebio,KZ-III-F);增強型ECL化學發光儀(AZURE Biosystem,C300);低溫冷凍離心機(Thermo fisher,ST8);多功能酶標儀(Thermo fisher,Multiskan SkyHigh);冰凍切片機(Leica,CM 1950)。
1.2 方法
1.2.1小鼠分組及EAE模型制備 將30只小鼠按照體質量隨機平均分為正常對照組、EAE模型組、GLPS組,將MOG35-55多肽稀釋成5 g·L-1,與等體積含TB的CFA混合,在EAE模型組和GLPS組小鼠脊椎周圍任選4點進行皮下注射(0.1 mL/只),正常對照組不予抗原處理。免疫后第0 h和48 h腹腔注射PTX(400 ng/只)增強免疫應答。從免疫當天開始,GLPS組小鼠每天腹腔注射給予GLPS[8](5 mg·kg-1),正常對照組及EAE模型組小鼠腹腔注射等體積生理鹽水,連續21 d。
1.2.2小鼠神經功能評分 造模后每天同一時間段記錄小鼠體質量變化。采用5分評分法進行神經功能評分[9]:0分,四肢正常,尾部沒有癥狀不發病;1分,尾部張力下降或無力;2分,尾部全癱或肢體輕癱;3分,雙后肢癱瘓或肢體全癱;4分,四肢癱瘓且被動翻身后不能復位;5分,瀕死或死亡。
1.2.3固藍染色檢測脊髓組織病理情況 將小鼠脊髓組織制成厚度為10 μm的冰凍切片,置于70%乙醇溶液中15 min后,取出切片放入配制好的LFB染液中,于60 ℃孵育過夜,之后依次放入95%乙醇、去離子水、0.05%碳酸鋰溶液中充分浸洗。經不同濃度乙醇梯度脫水、二甲苯透明、中性樹膠封片后置于顯微鏡下觀察。
1.2.4HE染色檢測脊髓組織炎性浸潤情況 將冰凍切片置于蘇木精染液中5 min,去離子水洗去浮色,滴加分化液分化1 min,去離子水浸洗兩次后(每次3 min),置于伊紅染液中1 min,取出切片依次放入75%乙醇、85%乙醇、95%乙醇、無水乙醇梯度脫水3 s,之后再置于無水乙醇中浸洗1 min、二甲苯透明、中性樹膠封固后鏡下觀察。
1.2.5MBP免疫熒光染色檢測髓鞘脫失情況 取出冰凍切片恢復至室溫后放入1× PBS中洗去包埋劑,用1% BSA+0.3% Triton X-100 (1× PBS配制)封閉通透30 min,加入按比例稀釋的MBP一抗,4 ℃冰箱過夜。第2天,1×PBS浸洗3次,每次5 min,加入對應種屬的熒光二抗,室溫避光孵育2 h后,1× PBS浸洗3次,用抗熒光淬滅封片劑封片,顯微鏡下觀察。
1.2.6流式細胞術檢測脾臟及淋巴結組織中IL-17、Foxp3的表達 將EAE模型組、GLPS組小鼠脾臟及淋巴結組織研磨過濾離心后用1%BSA制備成細胞懸液,分裝于相應的流式管中,分別加入檢測的流式抗體,于室溫下避光20 min。PBS洗滌3次,最后每管加入500 μL PBS,用流式細胞儀檢測IL-17、Foxp3的表達。
1.2.7Western blot檢測脊髓組織中MBP、TLR4、MyD88、NF-κB的表達 取小鼠脊髓組織加入RIPA裂解液,于低溫組織研磨儀研磨成組織勻漿,將其放入4 ℃離心機,10 000 r·min-1離心15 min后取上清,采用BCA法測定蛋白濃度,取上清加入Loading Buffer并煮沸10 min備用。通過SDS-PAGE分離等量的蛋白質(20 μg)并濕轉到PVDF膜上,5%脫脂奶粉封閉1 h后,按照稀釋比例分別加入相應的一抗并放入4 ℃冰箱過夜。第2天,TBST浸洗3次后,加入對應種屬的二抗,室溫孵育2 h,TBST浸洗3次。通過增強型ECL化學發光儀檢測并攝取蛋白條帶圖像,ImageJ軟件分析條帶圖像的平均灰度值。
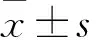
2 結果
2.1 靈孢多糖對EAE小鼠體質量和神經功能的影響Fig 1A結果顯示,與正常對照組相比,EAE模型組小鼠出現明顯的精神不振、進食量減少等情況,從第9天開始體質量出現明顯下降趨勢(P<0.05,P<0.01);與EAE模型組相比,GLPS組體質量下降情況相比于模型組更為緩和,從第13天開始明顯減緩(P<0.05和P<0.01);而且,EAE模型組小鼠在第11天開始出現神經系統功能障礙,伴隨尾部張力下降、后肢癱瘓或前肢癱瘓、大小便失禁等表現(P<0.05),第16天開始進入高峰期,表現更為明顯(P<0.01);而GLPS組小鼠各時間段神經功能臨床評分均低于EAE模型組,發病潛伏期推遲,進入高峰期評分明顯降低(P<0.01),見Fig 1B。實驗結果表明,GLPS能夠減輕EAE誘導的小鼠體質量減輕并且緩解其臨床癥狀。
2.2 靈孢多糖減輕EAE小鼠的髓鞘脫失和炎性浸潤Fig 2結果顯示,與正常對照組相比,EAE模型組小鼠脊髓白質可見大范圍髓鞘脫失區域,伴隨明顯的空泡形成(P<0.01),大量炎性細胞浸潤(P<0.01),并且MBP表達明顯低(P<0.01);與EAE模型組相比,GLPS組小鼠脊髓白質脫髓鞘范圍與空泡形成均減少(P<0.01),僅有少量炎性細胞浸潤(P<0.01),MBP表達明顯增高(P<0.01)。說明GLPS能夠減輕EAE小鼠的髓鞘脫失和炎性浸潤。
2.3 靈孢多糖影響EAE小鼠脾臟和淋巴結中IL-17、Foxp3的表達Fig 3流式細胞術結果顯示,與EAE模型組比較,GLPS組小鼠脾臟及淋巴結組織中IL-17明顯降低(P<0.01),而Foxp3明顯增高(P<0.01)。說明GLPS能夠通過下調促炎因子IL-17的表達減輕炎癥程度,上調Foxp3的表達抑制免疫反應,調節免疫平衡。

Fig 1 GLPS intervention reduced weight loss and clinical behavior in EAE mice
2.4 靈孢多糖對EAE小鼠脊髓組織中TLR4/NF-κB通路蛋白表達水平的影響Fig 4結果顯示,與正常對照組相比,EAE模型組小鼠脊髓組織中TLR4、MyD88、p-NF-κB蛋白表達量均明顯升高(P<0.01);與EAE模型組比較,GLPS組小鼠脊髓組織中TLR4、MyD88、p-NF-κB蛋白表達量均明顯降低(P<0.01)。可以推測,GLPS可以通過抑制TLR4/NF-κB通路來改善EAE小鼠炎性反應,改善髓鞘脫失。
3 討論
近年來,雖然針對MS患者的疾病修飾療法取得了一定的進展,但MS的整體治療效果尚未最大化,開發新的安全有效的藥物仍然是一個非常重要的目標。GLPS作為天然藥物靈芝的主要生物活性成分,具有抗氧化、抗炎、免疫調節等作用,并且藥性穩定、毒副作用小[6]。研究表明,GLPS能夠通過抑制MAPK信號通路保護成纖維細胞免受紫外線誘導的氧化衰老作用[10],并且在多種疾病中表現出抗炎或免疫抑制作用,包括AD、PD等神經退行性疾病[7]。在本研究中,通過構建小鼠EAE模型,我們發現小鼠出現體質量下降以及不同程度的神經功能障礙,病理檢測觀察到小鼠脊髓組織中大范圍髓鞘脫失及炎性細胞浸潤情況,同時維持髓鞘結構和功能的穩定的標志物MBP表達顯著降低,說明模型是成功的。GLPS干預有效地抑制了EAE小鼠的體質量的下降及神經功能評分的升高,改善了小鼠脊髓組織的髓鞘脫失和炎性浸潤,增加了MBP的表達,證實了GLPS對EAE小鼠有較好的治療作用。

Fig 2 GLPS reduced myelin loss and inflammatory infiltration in EAE mice
自身反應性CD4+T細胞在EAE的發生發展過程中起主要作用,活化的CD4+T細胞從外周遷移到CNS,在效應性T細胞活化過程中,細胞因子和趨化因子的產生影響局部微環境,最終決定免疫反應的結果[11]。其中IL17產生跨髓磷脂成分的T細胞自身反應是MS發病機制的核心,IL-17是T細胞誘導的炎癥反應的早期啟動因子,可以通過促進釋放炎性細胞因子來放大炎癥反應[12]。對于機體內自身免疫效應的過度活化,往往需要調節性T細胞(regulatory T cells,Tregs)來維持免疫自我耐受性并預防多種自身免疫性疾病,Tregs缺乏或抑制功能受損會導致嚴重的自身免疫性疾病和慢性炎癥,而Foxp3被認為是Tregs發育,穩定性,特別是抑制功能的主要轉錄因子[13]。在先前的研究中,已經證明了通過降低IL-17的含量,增加Foxp3的表達對EAE治療有明顯的改善作用[14]。本研究顯示,與EAE模型組比較,經GLPS干預后EAE小鼠脾臟和淋巴結組織中IL-17表達顯著降低,Foxp3表達顯著升高,表明GLPS可以通過抑制炎性細胞因子以及效應性T細胞的活化來減輕EAE小鼠的炎癥反應和過度的免疫效應。
TLR4/NF-κB信號通路是一條關鍵的炎癥信號轉導途徑,密切參與細胞分化、增殖、凋亡和促炎反應[15]。其中TLR4是第一個被鑒定的哺乳動物TLR,是在炎癥信號轉導中起關鍵作用的天然免疫受體。相關研究表明,使用TLR4配體(如脂多糖)干預會加重EAE,而靶向破壞TLR4會阻止EAE的進展[16]。激活的TLR4能夠通過其關鍵接頭分子MyD88形成復合物,并最終激活NF-κB,NF-κB作為TLR4信號通路的下游效應子,是調節T細胞、巨噬細胞和小膠質細胞等炎癥細胞活性的主要轉錄因子之一,可介導多種炎癥過程。已經證明NF-κB在MS患者的CNS的多種細胞類型中被激活,對疾病的發展有明顯的影響[17]。Yoon等[18]的研究顯示,靈芝的乙醇提取物能夠通過抑制TLR4/NF-κB途徑來抑制脂多糖誘導的小膠質細胞中的炎癥反應。但對GLPS通過抑制TLR4/NF-κB途徑發揮抗炎作用機制的研究還知之甚少。本研究發現,EAE模型組小鼠脊髓組織中TLR4/NF-κB蛋白表達水平相較于正常對照組明顯升高,經GLPS干預后TLR4/NF-κB蛋白表達水平明顯降低,說明GLPS可能通過抑制TLR4/NF-κB通路的活化來減緩炎癥反應從而治療EAE小鼠。

Fig 3 Effects of GLPS on levels of Th17 and Treg cells in spleen and lymph nodes of EAE

Fig 4 Effects of GLPS on expression of TLR4/NF-κB in spinal cord of EAE mice **P<0. 01 vs EAE group
綜上所述,GLPS能夠明顯改善EAE小鼠的髓鞘脫失和炎癥反應,其作用機制可能與抑制TLR4/NF-κB信號通路激活有關。但這項研究仍有一定的局限性,GLPS如何抑制TLR4/NF-κB信號通路活化以及這條通路的相關作用性的關鍵程度仍然需要進一步的體外實驗去驗證。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
數學物理學報(2020年2期)2020-06-02 11:29:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00