豬瘟病毒致弱機制研究進展
2023-10-26 12:21:40陳紫昱夏應菊宋新宇劉業兵
中國獸藥雜志 2023年9期
關鍵詞:研究
陳紫昱,徐 璐,夏應菊,宋新宇,劉業兵
(中國獸醫藥品監察所國家豬瘟參考實驗室,北京 100081)
豬瘟(Classical Swine Fever,CSF)是由豬瘟病毒(Classical Swine Fever Virus,CSFV)引起的豬最重要的傳染病之一,給全球養豬業帶來了巨大的經濟損失,是世界動物衛生組織規定的必須上報的豬病之一,我國2022年將其調整為二類動物疫病。CSFV屬于黃病毒科瘟病毒屬,為二十面體的囊膜病毒,具有約12.3kb的單股正鏈RNA基因組。其基因組包含一個開放閱讀框(ORF),5’端和3’端各有一個非編碼區(UTR)。病毒顆粒由4種結構蛋白(核心蛋白C、包膜糖蛋白Erns、E1和E2)以及8種非結構蛋白(Npro、p7、NS2、NS3、NS4A、NS4B、NS5A和NS5B)組成[1]。針對CSF的防控,由強毒株致弱制成的減毒活疫苗是應用最為廣泛的。弱毒疫苗Thiverval株是OIE推薦的疫苗,該疫苗株是從原始強毒Alfort毒株衍生的減毒疫苗株,具有可靠的保護效力和快速的保護能力。除此之外,還有日本的減毒疫苗株GPE菌株,GPE菌株是由高毒性菌株ALD先在豬睪丸細胞中的142次傳代、在牛睪丸細胞中的36次傳代和在原代豚鼠腎(pGPK)細胞中的32次傳代獲得的,該疫苗株使得日本在20世紀免受CSF的困擾。菲律賓的Lapinized Philippines Coronel(LPC)疫苗也是一種減毒的CSFV毒株,是預防和控制CSFV感染的重要工具,在CSF流行區廣泛使用。20世紀五十年代,中國科學家經過兔體連續傳代和系統測試,培育出了一株適應于家兔的 CSFV兔化弱毒株(Hog Cholera Lapinized Virus,HCLV)。豬體試驗表明該毒株不僅對豬安全,還保持了優良的免疫原性。用該毒株免疫豬 4 天即可產生免疫保護力,一次接種產生的免疫保護期可達一年以上。該毒株于1956年用于生產弱毒疫苗,并在全國推廣使用。同年,中國政府將該毒株以先進科技成果先后無償贈送給前蘇聯、匈牙利、羅馬尼亞、朝鮮、保加利亞及越南等當時的社會主義友好國家。匈牙利應用證明,來自北京的兔化弱毒株優于美英當時的Rovac及SFV等商品弱毒苗,它對種豬、乳豬無殘余毒力。HCLV種毒之后被傳到大部分歐洲國家及一些拉美國家,并且該疫苗榮獲 1983 年國家發明一等獎。時至今日這一毒株仍在我國和世界上許多國家廣泛使用,是國際上公認最安全有效的豬瘟弱毒疫苗,因此國際上通常將該疫苗株稱為“C”株,即“中國”株。有研究表明,CSFV通過兔體傳代,導致E2基因關鍵氨基酸位點突變,促進豬瘟病毒侵入家兔脾臟淋巴細胞,進而賦予CSFV對家兔的適應性[2]。豬瘟兔化弱毒疫苗應用已有多年的歷史,但其毒力致弱的分子機制還尚未完全明確。并且目前使用的減毒活疫苗會干擾血清學診斷,使得鑒別診斷存在困難,因此繼續研發新的診斷工具和新型疫苗十分必要。隨著分子生物學技術的發展,對CSFV的毒力基因有了更為深入的研究。本文從病毒結構角度對CSFV致弱機制的國內外研究成果進行綜述,以期為后續的研究以及候選疫苗的開發提供參考。
1 基于結構蛋白E2的致弱機制研究
E2蛋白位于病毒囊膜表面,是一種功能重要的糖蛋白,分子量大小約為55kDa,其空間結構由三個疏水區和三個氨基端的鏈內二硫鍵構成。在靠近E2蛋白氨基端上游有一段信號肽序列,羧基端有一段跨膜區,E2蛋白由A、B、C、D四個抗原結構域組成(圖1)。E2蛋白是CSFV毒力的決定因子之一,也是免疫原性最好的結構蛋白,能誘導中和抗體的產生[3]。
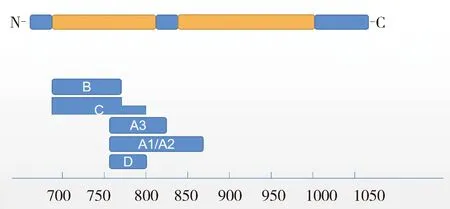
圖1 圖1 E2蛋白基因結構圖Fig 1 The structure diagram of E2 protein
早在2005年,Risatti等用疫苗株E2基因替換強毒株中的E2基因,構建的嵌合病毒在豬體內的毒力明顯降低,從而證明E2基因與毒力致弱直接相關。更加深入的研究發現,E2蛋白羧基端882-1032之間存在一段12個氨基酸的取代(T886M、P889L、Q892R、S927L、T928A、H955R、D958G、A975E、R979S、A988T、R994K和I1032V)可以降低病毒致病性[4]。研究表明,與C株同為1.1亞型的經典強毒株石門株在E2基因上存在差異位點,這一結果提示E2基因可能是導致C株致弱的原因之一。E2蛋白與宿主蛋白DCTN6之間的相互作用也可以影響病毒毒力,介導與DCTN6相互作用的關鍵E2位點突變后的重組突變體E2ΔcDCTN6v在豬中被完全減毒[5]。E2蛋白與核糖體蛋白的相互作用也會影響病毒毒力,敲除核糖體蛋白RPLP1基因可以使病毒滴度降低[6]。E2蛋白的糖基化位點是影響CSFV毒力的重要因素,研究發現C株E2蛋白在序列基序986 NYT988位點有獨特的糖基化,這可能是C株毒力減弱和提供完全保護的原因[7]。
E2糖蛋白對CSFV的免疫原性同樣有著十分重要的作用,它不僅可以誘導抗體的產生,而且也是誘導特異性細胞毒性T淋巴細胞(CTL)應答的主要蛋白。基于E2蛋白的免疫原性,將E2蛋白和CD154抗原結合制成的改良疫苗具有良好的保護效力[8],并且國內外也有商品化的豬瘟E2蛋白亞單位疫苗上市。最近的研究表明,E2蛋白CD結構域邊界鑒定出新的線性表位P33,具有中和活性并且可用于CSFV與牛病毒性腹瀉病毒(Bovine viral diarrhoea virus,BVDV)的鑒別診斷[9],但其對病毒致弱的作用有待進一步的探究。
E2蛋白和CSFV的其他區域存在相互作用,共同調控CSFV的感染過程。孫永科等[10]利用構建重組腺病毒的方法,對不同處理組的豬進行了觀察,發現E2和Erns結合的重組病毒可以對豬只提供保護。仇華吉等[11]研究發現表達C株Erns-E1-E2、Erns-E2或E1-E2的嵌合病毒,在兔子脾臟中可檢測到與C株相類似的病毒復制水平,這表明E2蛋白和Erns或E1協同作用很有可能是C株致弱的原因之一。更加深入的研究顯示,E2結構域I中的氨基酸P108和T109對于C株在家兔體內的致弱是關鍵的[2]。最近的研究表明,用石門株中的Erns或E2替代C株的相應部位構建的嵌合體病毒誘導輕度的發熱反應,而E2和Erns均被替換的嵌合體病毒則消除了發熱反應并且在兔體內沒有表現出適應性[12],這同樣表明E2蛋白和Erns之間的相互作用是C株在家兔體內致弱的關鍵。E2蛋白和3’UTR在減毒嵌合體病毒基因組的穩定性方面表現出協同作用,E2或3’UTR的替代均可降低病毒的復制。E2蛋白是目前研究最為廣泛的蛋白,對E2蛋白關鍵位點的綜述,將為研制新型疫苗和診斷制劑提供理論基礎。深入研究E2蛋白與病毒致弱的關鍵位點,有利于通過精準的基因編輯來實現新的標記疫苗的研發。
2 基于結構蛋白Erns的致弱機制研究
Erns是分子量為44kDa的囊膜糖蛋白,由227個氨基酸組成,共有7個糖基化位點。Erns通常以二聚體形式存在,是CSFV毒力的必需因子。Erns也能夠刺激中和抗體的產生,但是單獨的Erns不能誘導保護性免疫反應,需要和E2蛋白共同存在[3]。
Erns對病毒的毒力有十分重要的意義,缺失Erns蛋白的糖基化位點能使病毒毒力減弱。Borca等[13]基于Brescia Erns基因構建了一組糖基化突變體,發現N269糖基化位點使病毒毒力減弱,這表明Erns與CSFV毒力致弱直接相關。Tucakov等[14]的研究發現缺乏半胱氨酸171的CSFV突變體影響了Erns二聚體的形成從而在豬體內被減弱;體內自發補償突變使Erns二聚體恢復后CSFV恢復了毒力,這證實了Erns二聚體的形成對CSFV毒力的重要性。Erns還具有細胞毒性作用,它可以抑制淋巴細胞的增殖和蛋白質的合成,并且尚未發現其細胞毒性作用具有種屬特異性,這種細胞毒性作用有可能是Erns與毒力相關的另一原因。Erns對CSFV毒力的影響可能是因為它具有核糖核酸酶(RNase)活性,而RNase具有神經毒性、免疫抑制、抗蠕蟲和抗腫瘤等作用。王苗苗等[15]構建了一株基于低毒力野生型CSFV的缺乏RNase活性的cDNA克隆,Erns的RNase活性的缺失增強了仔豬體液免疫反應,降低了病毒的毒力、傳染性和持久性。與RNase活性有關的關鍵氨基酸位點被取代后,CSFV的致病性減弱或喪失。Iqbal等[16]研究發現,對CSFV和BVDV的Erns基因進行修飾,使其喪失RNase活性后病毒致病性減弱,這表明RNase活性與毒力致弱直接相關。RNase活性還可以阻止ssRNA、dsRNA誘導的干擾素(IFN)表達,在自然界發現的任何瘟病毒表達的Erns幾乎都是IFN的拮抗劑[17],影響RNase活性的突變(H30F、PR突變)會顯著減弱病毒對IFN表達的抑制作用[18]。Python等[19]構建了無RNase活性的CSFV突變體,結果顯示,無RNase活性的突變體誘導的IFN-α含量是野生型CSFV誘導的10倍。而逃避IFN反應很有可能是CSFV誘導持續性感染的核心因素[20],已有研究表明C株疫苗在接種的早期緩慢復制,避免了IFN-α和IFN-β的過早過度表達,這種抑制IFN產生的特性可能是C株致弱的機制,這也表明Erns影響毒力的機制可能是其RNase活性對IFN表達的調控。Erns蛋白是決定病毒毒力的重要因素,更好的了解Erns與病毒復制以及病毒毒力的關系為開發針對CSFV的DIVA疫苗提供了新的研究方向,從而有利于區分CSF與其他瘟疫病毒,并為有效控制和根除CSF做出貢獻。
3 基于非結構蛋白的致弱機制研究
CSFV病毒顆粒包含8種非結構蛋白(Npro、p7、NS2、NS3、NS4A、NS4B、NS5A和NS5B),這些非結構蛋白在CSFV感染過程中發揮著各自的作用。Npro是CSFV特有的一種半胱氨酸樣蛋白,它可以從正在翻譯的多聚蛋白上斷裂并成熟,其分子量約為19kDa。Npro有可能與病毒毒力相關,Mayer等[21]構建了缺失Npro基因的突變體,發現中等毒力和強毒株突變體都導致毒力的減弱,這表明Npro與病毒毒力致弱相關。研究發現,Npro蛋白可以抑制宿主的免疫應答,Npro既可以拮抗dsRNA介導的TRL3依賴和非依賴性途徑中Ⅰ型IFN的誘導,又可以通過降解TRL3的蛋白酶體抑制I型IFN的誘導[22],同時還可以通過抑制干擾素調節因子(IRF1)的表達來抑制Ⅲ型IFN的產生[23]。Npro蛋白很有可能參與細胞蛋白的泛素化修飾,抑制細胞增殖,誘導I型輔助性T淋巴細胞的凋亡。Npro還可以拮抗TRL3和視黃酸誘導基因蛋白Ⅰ(RIG1)介導的細胞凋亡,從而使病毒不易被清除,這可能是CSFV免疫逃避的一種機制[24],這種拮抗宿主對CSFV入侵的反應是否是Npro影響毒力的原因還有待更加深入的研究。Npro與核糖體蛋白S20(U10)的相互作用可以阻止CSFV在體內的復制,并且在CSFV感染過程中發揮抗病毒作用[25],然而S20降低病毒在體內含量的機制尚不明確。Npro蛋白對病毒復制以及宿主免疫應答的作用可能是缺失Npro基因的突變體毒力致弱的原因之一,但更深入的機制還值得更進一步的研究。
P7蛋白是一種分子量約為6kDa~7kDa的疏水性小跨膜蛋白,在CSFV毒力中起著重要作用。Gladue等[26]通過丙氨酸掃描突變創建了一組在P7基因上含有3~6個連續氨基酸殘基替換的重組突變體,研究了接種動物的臨床癥狀和體內病毒滴度,發現P7蛋白與病毒毒力致弱直接相關。林志等[27]利用質粒pEGFP-p7轉染巨噬細胞,對IL-1β的mRNA蛋白含量進行監測,發現P7可以誘導巨噬細胞分泌IL-1β,這表明P7蛋白參與CSFV在體內介導的炎癥反應,但其與病毒致弱的關系有待進一步的研究。趙成等[28]的研究結果顯示,P7蛋白TM1區的3個氨基酸取代(p7TDI18/19/20AAA,p7EVV21/22/23AAA和p7YFY25/26/30AAA)抑制了感染性病毒的產生,病毒滴度顯著降低。P7可以與宿主蛋白CAMLG相互作用,破壞這種作用會影響CSFV在細胞培養中復制的能力[29],這為病毒毒力致弱機制的研究提供了一定的參考。
NS3蛋白在CSFV的復制、轉錄和翻譯以及病毒多聚蛋白的加工過程中起到重要作用[30]。朱洪昌等[31]用丙氨酸或天冬氨酸單取代或全部取代NS3蛋白上保守的四個堿性氨基酸殘基以破壞NS3蛋白上保守的帶正電荷的區域,結果顯示突變體VH24A、VR50A、VH24D、VR50D或VK74D的病毒滴度顯著降低,這表明 NS3是病毒致弱的潛在靶點。鄧少峰等[30]研究發現宿主蛋白PSMB10通過泛素-蛋白酶體途徑刺激NS3蛋白降解來抑制CSFV增殖。李小蒙[32]等發現腫瘤壞死因子受體相關因子5(TRAF5)與NS3相互作用,TRAF5通過激活p38 MAPK通路促進CSFV復制。NS3與病毒復制密切相關,然而關于是否可以突變NS3抑制病毒復制,從而降低病毒滴度,導致病毒對豬體毒力下降的研究還需深入的探索。
非結構蛋白NS4A、NS4B、NS5A、NS5B通常與病毒復制與合成相關,影響病毒復制的位點是影響毒力的潛在位點。病毒在豬體內復制效率下降,可能會導致病毒滴度降低.從而導致病毒對豬體的毒力下降.成為對豬體不產生致病效應的減毒株[33]。馬立新等[34]研究發現CSFV的NS4A蛋白是一種抑制哺乳動物細胞中RNA干擾(RNAi)的新RNA沉默病毒抑制因子(VSR),VSR作用可以幫助CSFV逃避宿主的抗病毒作用,VSR缺陷型CSFV的復制被減弱,這為開發減毒活疫苗提供了依據。張彥明等[35]證明NS4A蛋白可以通過增強MAVS途徑誘導IL-8產生,并促進CSFV復制。NS4B蛋白可以與宿主的核糖體蛋白RPLP1相互作用,通過促進病毒基因組的轉譯增加病毒產量[36]。曹志等[37]研究表明,NS4B的表達可以顯著抑制TRIF、IRF3和NF-κB p65的表達,從而抑制poly (I:C)刺激后IFN-β和IL-6的分泌,影響TLR介導的先天免疫應答。張彥明等[38]利用慢病毒介導的組成型過表達和真核質粒瞬時過表達方法,對Rab5敲除后細胞中病毒RNA含量進行了研究,結果表明,敲除Rab5對CSFV產生有抑制作用,并且參與NS4B復合物的形成。非結構蛋白NS5B是一種RNA依賴的RNA聚合酶,CSFV NS5B的162位點的氨基酸殘基由脯氨酸突變為蘇氨酸,可以增加RNA依賴RNA聚合酶(RdRP)活性,促進CSFV基因組的復制[39]。非結構蛋白NS5A在低濃度時可以與NS5B結合形成復制復合體,促進病毒RNA的合成;當NS5A的濃度超過NS5B時,NS5A可以與NS5B和3’UTR相互作用,抑制病毒的復制[40],這可能是NS5A通過濃度變化調控病毒復制的原因。肖明等[41]利用谷胱甘肽S-轉移酶下拉試驗和免疫沉淀分析表明位于NS5A末端的的氨基酸K399、T401、E406和L413對NS5A-NS5B相互作用是必需的,這種相互作用可能是NS5B蛋白和NS5A蛋白對核糖體內部位點介導(IRES)的翻譯產生影響的機制。
綜上所述,非結構蛋白對病毒毒力和病毒生命周期至關重要。我們可以更加全面地了解非結構蛋白對病毒感染的作用,以便更深入地研究這些蛋白中負責病毒毒力的專門區域,有助于設計更有效的生物工具來控制CSF的發生。
4 基于3’UTR的致弱機制研究
CSFV的3’非編碼區(3’UTR)是一個富含AU的區域,該區域也與病毒的毒力相關。王毅等[33]發現在C株的3’UTR中存在12個堿基序列(CUUUUUUCUUUU)的插入,這是導致中國豬瘟兔化弱毒疫苗減毒的關鍵因素。低毒力的經典豬瘟病毒Pinar del3’UTR中發現了一段平均長度為36個尿苷的新型聚尿苷束(poly-U),這可能是病毒毒力衰減的原因[42]。在減毒活疫苗Thiverval 3’UTR的233~259 bp之間,存在著一段6-32nt的可變長度的富T序列,推測該插入是Thiverval毒力衰減的原因之一[43]。王苗苗等[44]在最近的研究中也發現在3’端插入poly-U可以導致病毒衰減,減少受感染仔豬排毒量,降低CSFV在體內的復制水平和疾病的嚴重程度。據報道,CSFV 3’UTR的3342位點的脯氨酸僅存在于兔化減毒株中,包括C株、Riems、LPC和HCLV-India株,這表明3342位點的脯氨酸可能是病毒在兔體內致弱的原因之一[39]。3’UTR還可以與Erns的RNase功能協同作用來調節宿主的先天性和適應性免疫應答,從而有利于CSFV在靶組織中的復制[45]。上述研究表明,3’UTR有著十分廣闊的研究前景,可以通過對3’UTR進行不同的修飾從而研究病毒毒力的變化,從而進一步揭示影響病毒致病力的分子機制。
5 小結與展望
近年來,隨著分子生物學等技術手段的發展,有關CSFV致弱機制的研究得到了極大的發展。CSFV的結構蛋白、非結構蛋白以及3’UTR對病毒致弱有著不同程度的影響。E2蛋白是病毒毒力的決定因子之一,也是免疫原性最好的結構蛋白。Erns蛋白是毒力的必需因子,通常和E2蛋白協同作用共同調節病毒毒力。非結構蛋白Npro可以調節病毒毒力,而NS3、NS4A、NS4B、NS5A和NS5B則通常在病毒復制過程中發揮作用。3’UTR通過堿基修飾可以使病毒致弱。上述研究成果均提示我們,豬瘟病毒的致弱機制非常的復雜,任何病毒基因的變化可能都會影響病毒的毒力,因此,對豬瘟病毒致弱毒株的研究和開發是一個系統的工程。現有的豬瘟弱毒疫苗雖然具有良好的保護性和有效性,但仍然面臨著新的病毒突變體和不能進行鑒別診斷的挑戰。本文綜述了CSFV可能存在的致弱機制,這些研究啟發未來新型標記疫苗的研發可以通過對CSFV結構蛋白和非結構蛋白的關鍵位點的基因修飾來實現,同時也為抗病毒藥物的開發提供新的研究思路。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
