基于非靶向代謝組學分析谷氨酸棒桿菌分泌表達外源蛋白代謝差異
2023-11-08 06:10:54黃丹妮鄒宇劉秀霞楊艷坤白仲虎
食品與發酵工業 2023年20期
黃丹妮,鄒宇,劉秀霞,2,3,楊艷坤,2,3,白仲虎,2,3*
1(江南大學,江南大學糧食發酵與食品生物制造國家工程研究中心,江蘇 無錫,214122) 2(江南大學,工業生物技術教育部重點實驗室,江蘇 無錫,214122)3(江南大學 生物工程學院,江蘇 無錫,214122)
谷氨酸棒桿菌(Corynebacteriumglutamicum)是高GC含量的革蘭氏陽性菌,由于其無內毒素,胞外蛋白酶活性低以及有完善的分泌系統可用于分泌生產重組醫藥蛋白,被認為是很有潛力的外源蛋白表達宿主。目前,為提高外源蛋白表達產量,主要從表達元件、宿主細胞以及發酵過程等方面開展C.glutamicum蛋白表達系統的優化[1],并取得了一定的進展。例如,表達元件方面開發的雙順反子表達結構[2]和3A(three assembly)元件庫[3],基于蛋白酶敲除和常壓室溫等離子體(atmospheric and room temperature plasma, ARTP)技術[4]獲得的高效蛋白分泌菌株以及以OmlA抗原為模式蛋白開發的高通量菌株篩選和發酵流程[5]。
盡管取得了以上成果,但依然存在外源蛋白分泌表達過高造成宿主細胞生長代謝負擔,進而影響蛋白產量的問題。已有研究表明,外源蛋白合成和運輸的每一過程都需要能量驅動[6],比如肽鏈延伸、氨酰-tRNA合成、外源蛋白轉運等。合成人生長激素(human growth hormone, hGH)每單位需要消耗267個單位ATP及420個單位的NADPH[7]。輔因子工程與傳統代謝工程和合成生物學策略相結合,通過控制ATP水解降低其總量,提高糖酵解的通量到野生型通量的1.7倍[8],有效增加了目的代謝物產量及微生物細胞工廠的效率。NAD+激酶基因ppnk和NADH激酶基因pos5的共表達顯著改善了NADPH的供應,這種NADPH再生策略簡單有效,不伴有碳損失,可用于生產需要NADPH消耗的其他天然產物[9]從而提高產量。同時,氨基酸是直接參與蛋白質翻譯的特殊代謝物,外源蛋白高表達對菌體內氨基酸量提出新的要求。過量生產氨基酸的菌株和外源添加氨基酸都由于能提供足夠的相應氨基酸,加速了報告蛋白的翻譯[10]。氨基酸需求增長導致各類氨基酸合成代謝活躍,氨基酸基礎代謝途徑產物如乙酰CoA進入三羧酸循環反作用于生長代謝,從而組成復雜的蛋白表達和能量代謝系統。因此從代謝水平解析外源蛋白分泌表達過程中的關鍵代謝物及代謝網絡,有助于挖掘宿主細胞改造的靶點進而優化宿主細胞提高目的蛋白產量。
本文以C.glutamicumCGMCC1.15647菌株為研究對象,以來源于駱駝血清的抗體重鏈可變區(variable domain of heavy chain of heavy chain antibody, VHH)為模式蛋白[11],通過非靶向代謝組學檢測培養液上清的代謝產物的差異,并結合生物信息學分析,從代謝水平上對C.glutamicumCGMCC1.15647株分泌外源蛋白過程中的關鍵代謝物及代謝調控網絡進行了分析,為后續改造C.glutamicum菌株提高其外源蛋白表達產量提供重要的理論支撐。
1 材料與方法
1.1 菌株與培養基
本文使用C.glutamicumBZH001菌株,中國普通微生物菌種保藏管理中心,保藏號為CGMCC 1.15647,在本實驗室保藏。所用培養基種類及配方見表1。

表1 培養基種類及配方Table 1 Type and formula of culture medium
1.2 儀器與設備
Vanquish UHPLC超高效液相、Q ExactiveTMHF-X高分辨質譜,Thermo Fisher(USA)公司。其余儀器與設備見附表1(https://doi.org/10.13995/j.cnki.11-1802/ts.034392)。
附表1 設備與儀器Table S1 Testing devices and instruments
1.3 實驗方法
1.3.1 質粒構建與轉化
使用引物對VHH_F/VHH_R,以實驗室前期構建的質粒擴增基因VHH[4],用plasmid_F/plasmid_R引物對使pXMJ19-control質粒線性化并在兩端添加20 bp同源序列,使用同源重組連接插入基因片段,得到質粒pXMJ19-Ptac-0949-VHH。
將pXMJ19-control和pXMJ19-Ptac-0949-VHH質粒轉化至谷氨酸棒桿菌BZH001中。平板上隨機挑選3個菌落PCR驗證為陽性的克隆,接種至LBB種子培養基于30 ℃,220 r/min恒溫振蕩培養箱過夜培養。次日以初始OD600=0.2轉接至培養基LBB中,培養2 h時添加1 mmol/L異丙基β-D-1-硫代吡喃半乳糖苷(isopropyl-beta-D-thiogalactopyranoside,IPTG)誘導VHH蛋白分泌表達。培養5 h后取樣并進行檢測。
1.3.2 蛋白表達及驗證
取培養5 h后的BZH001和VHH菌液,12 000 r/min離心1 min,吸取80 μL上清液,加入5×loading buffer,沸水煮10 min。使用翌圣15%預制膠,10 μL上樣,90 V,15 min;120 V,45 min。加熱5 min使用考馬斯亮藍對蛋白膠進行染色,脫色液加熱15 min,重復3次,直至底色從藍色變為透明。
1.3.3 樣品制備
胞外代謝物收集要求菌處于對數生長期,選取培養5 h的菌液快速從培養瓶中取出一定量的培養液,4 ℃條件下離心(1 000×g,10 min)。移取上清液500 μL至新的離心管中,迅速放入液氮中淬滅30 s;淬滅后放入-80 ℃中保存。共取3份樣品以滿足后續檢測需要及運輸損耗。
1.3.4 代謝物提取
取1 mL樣本于凍干機中凍干,加入100 μL的80%甲醇/水溶液;渦旋振蕩均勻后在冰水混合物中靜置5 min;在低溫高速離心機中保持4 ℃,15 000×g轉速離心15 min;使用LC-MS級別的水稀釋上清液至甲醇含量為53%;以15 000×g轉速在離心機中4 ℃離心15 min,吸取上清液,進樣LC-MS進行分析。
質控(quality control, QC)樣本:從每個實驗樣本中取等體積樣本混勻作為QC樣本,檢測時利用QC樣本進行質控,本文中所有原始數據的質量控制均基于QC樣本進行。
1.3.5 色譜及質譜條件
色譜條件:搭載Hypesil Gold column(C18)色譜柱(100 mm×2.1 mm,1.9 μm)。正離子電離模式:流動相A:0.1%甲酸;流動相B:甲醇。負離子電離模式:流動相A:5 mmol/L醋酸銨,pH 9.0,流動相B:甲醇。洗脫條件:0~1.5 min,2% B;1.5~3 min,85% B;3~10 min,100% B;10~10.1 min,2% B;10.1~12.0 min,2% B;流速0.2 mL/min,柱溫為40 ℃。
質譜條件:掃描范圍選擇m/z為100~1 500;ESI源的設置如下:噴霧電壓3.5 kV;鞘氣流速35 arb;輔助氣流速10 arb;離子傳輸管溫度320 ℃;離子導入射頻電平(S-lens RF level)60;輔助氣加熱器溫度350 ℃;極性:positive,negative;MS/MS二級掃描為數據依賴性掃描(datadependent scans)。
1.3.6 差異代謝物分析
采用有監督的正交偏最小二乘判別分析(orthogonal partial least squares discrimination analysis, OPLS-DA)模型對G1和G2組進行區分以展示其組間代謝總體差異特性。其中以變量投影重要性(variable importance projection, VIP)>1,P<0.05為標準對代謝物進行篩選并將符合標準的代謝物認為是表達顯著差異的代謝物,可能是潛在的生物標志物。將差異代謝物通過KEGG數據庫(https://www.kegg.jpkeggpathway.html/)中的谷氨酸棒桿菌數據庫(Select prefix:cgl,cgm)進行代謝通路注釋,獲得差異代謝物參與的通路。使用R軟件包ggplot2(Version 4.2.1)進行主成分分析(principal component analysis, PCA)和可視化,MetaboAnalyst 5.0數據庫(https://www.metaboanalyst.ca)和Omicshare(https://www.omicshare.com/)進行KEGG通路富集分析。
2 結果與分析
2.1 樣品制備
谷氨酸棒桿菌重組菌株構建及其外源蛋白表達驗證如下。將pXMJ19-control和pXMJ19-Ptac-0949-VHH質粒分別轉入C.glutamicum,并命名為G1和G2,對其進行生長曲線測定,以探究表達外源蛋白對C.glutamicum生長的影響。由圖1-a可知,2組均在發酵4 h進入對數生長期,10 h進入穩定期;發酵2 h后添加了IPTG誘導VHH表達,G2組OD600值略低于G1說明表達外源蛋白對菌株生長有輕微影響,但總體趨勢與對照組一致且穩定期菌體密度差異小。培養5、6、7 h的G2組如圖1-b中7~9條帶所示,結合生長曲線認為5 h處于對數生長期并已有蛋白分泌故選擇5 h 作為合適的取樣時間。選取發酵5 h的菌液上清作為分析樣品,與對照組G1(圖1-b,條帶1)相比G2組在15 kDa處有明顯條帶(圖1-b,條帶2~6)。
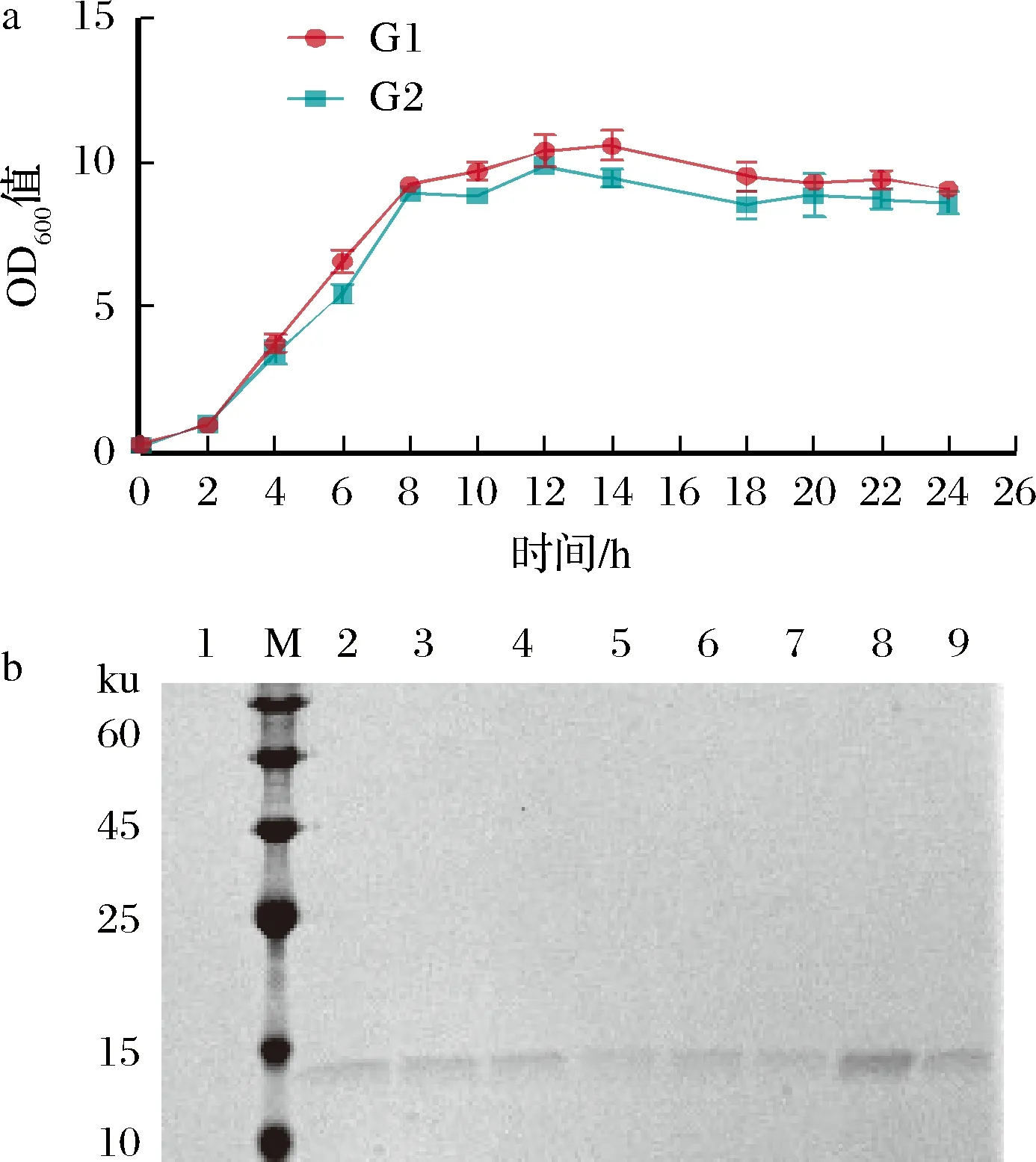
a-G1和G2的4 h生長曲線;b-菌液上清SDS-PAGE驗證(M-三色預染Marker;條帶1-攜帶pXMJ19-control的G1的發酵5 h的上清液;條帶2~6-攜帶pXMJ19-Ptac-0949-VHH的G2發酵5 h的上清液;條帶7~8-G2發酵5 、6 、7 h的取樣上清液)圖1 生長曲線和外源蛋白表達驗證Fig.1 Growth curves and validation of exogenous protein expression
2.2 代謝組學結果分析
2.2.1 差異代謝物分析
采用非靶向代謝組學手段,鑒定G1和G2兩組發酵液中代謝物并分析其變化規律。對代謝組結果進行可視化處理,結果顯示如圖2-a和圖2-c所示,組間PCA分離明顯,R2X均>0.5(附表2,https://doi.org/10.13995/j.cnki.11-1802/ts.034392)判斷組內樣本聚集良好。對2組代謝物進行了OPLS-DA分析,結果顯示R2X>0.5,R2Y和Q2值均接近于1(附表2),表明模型愈為穩定可信性高。進一步采用置換檢驗來驗證模型的預測能力和魯棒性,經過200次排列,正負離子電離模式下R2Y和Q2建模值與真實值組成的回歸線截距分別如圖2-b、圖2-d所示。R2在正負離子中分別為0.936和0.699,Q2回歸線截距在正負離子中分別為-0.15和-0.417,表明OPLS-DA模型擬合處于正常狀態,后續分析的數據可靠、結果可信。
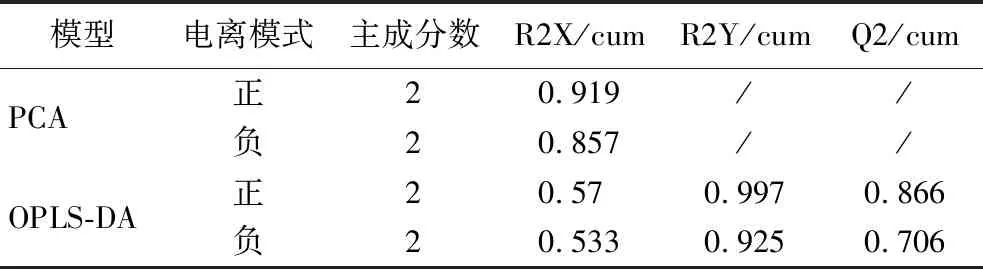
附表2 PCA和OPLS-DA模型參數Table S2 PCA and OPLS-DA model parameters
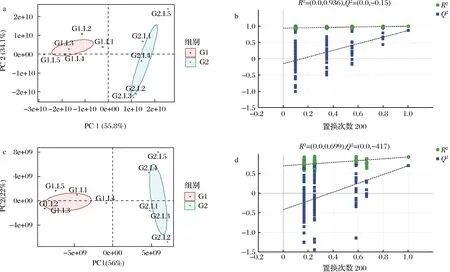
a-正離子電離模式下PCA得分散點圖;b-正離子電離模式下OPLS-DA模型的置換檢驗結果;c-負離子電離模式下PCA得分散點圖;d-負離子電離模式下OPLS-DA模型的置換檢驗結果圖2 正離子和負離子電離模式下G1和G2的PCA模型和置換檢驗。Fig.2 PCA model and permutation test results of G1 and G2

表2 本實驗所用引物Table 2 Primes used in this study
2.2.2 差異代謝物篩選
在正負離子電離模式下,共檢測到933個代謝物,其中正離子631個,負離子302個。根據t檢驗得到的P值、OPLS-DA模型計算出的VIP值和代謝物濃度倍數值(fold change, FC),繪制火山圖以展示代謝物豐度差異(圖3)。以VIP>1和P<0.05為標準篩選出BZH001菌株中分泌表達VHH蛋白導致產生差異濃度的代謝物,共176個代謝物,其中正離子100個,負離子76個。
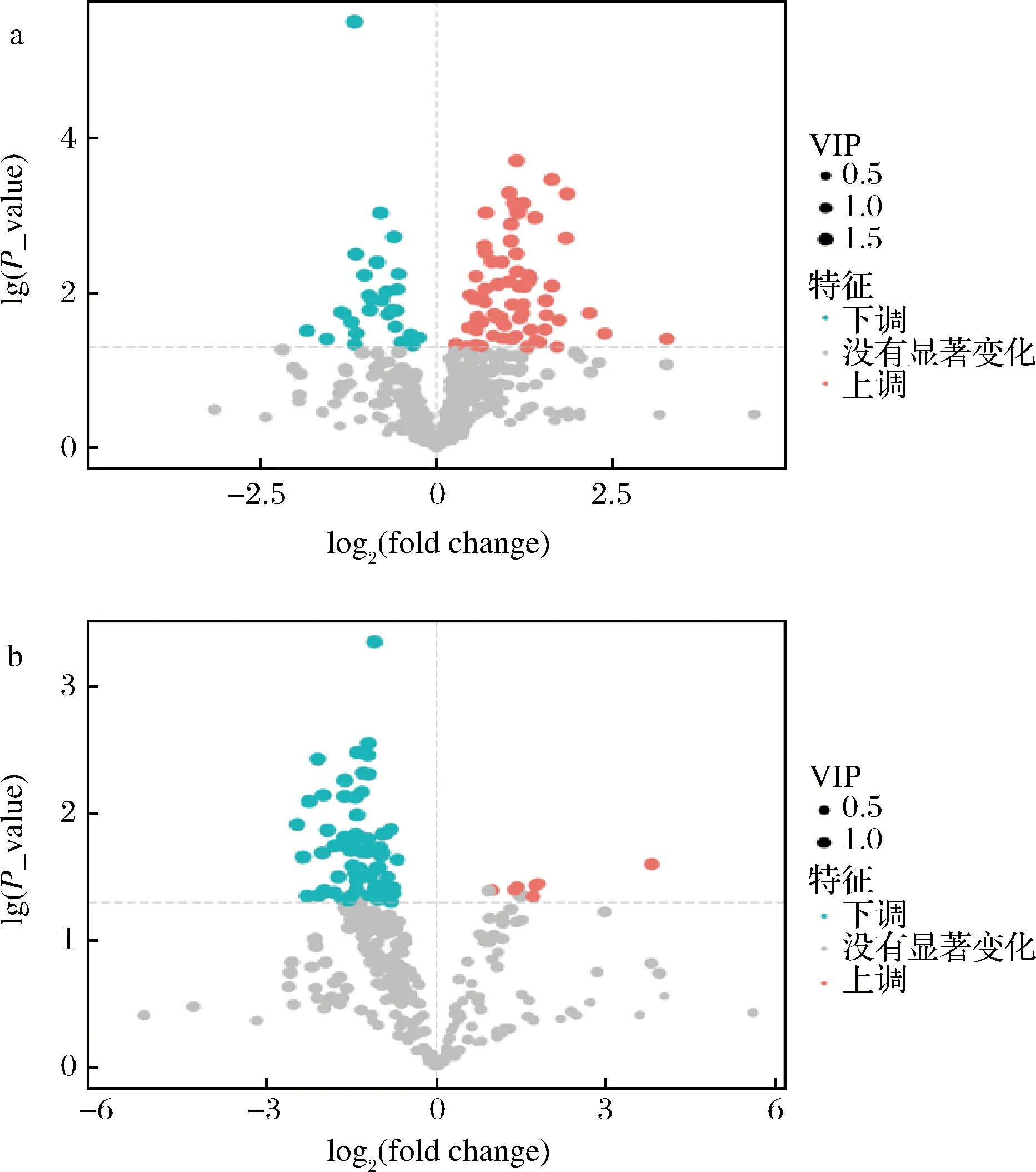
a-正離子下代謝物火山圖;b-負離子下代謝物火山圖圖3 正離子和負離子電離模式下G1和G2的代謝物篩選火山圖Fig.3 Volcano plots of screening metabolites
取代謝物豐度log轉化值并繪制差異代謝物聚類熱圖,可以直觀地看出各差異代謝物相對濃度的變化。如圖4所示,差異代謝物2組樣本中可明顯區分,說明分泌表達外源蛋白導致菌株代謝發生了顯著變化。分泌表達外源蛋白VHH導致其中79個代謝物在VHH組中顯著增加,其中正離子電離模式72個,負離子電離模式7個;97個代謝物顯著降低,其中正離子電離模式28個,負離子電離模式69個。
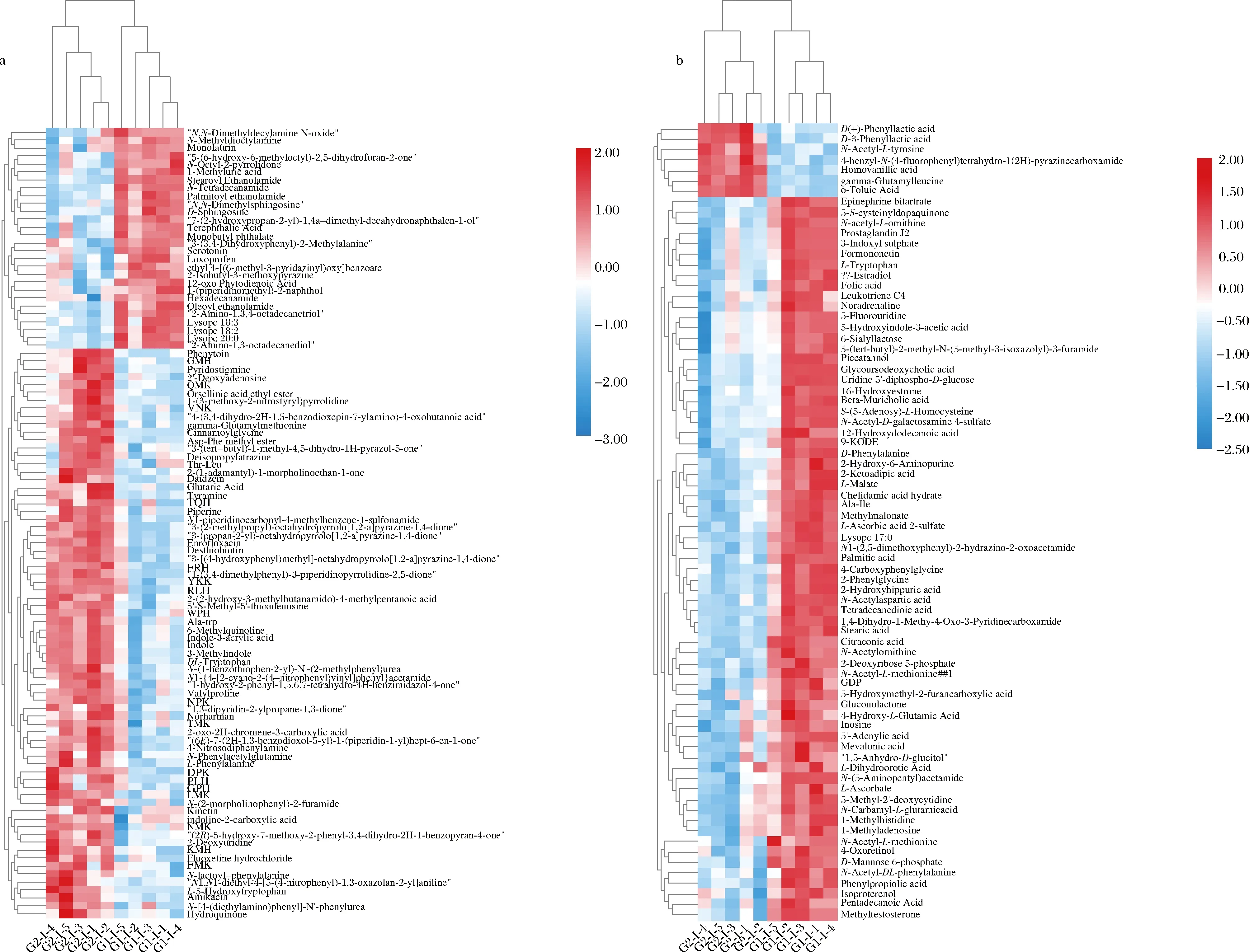
a-正離子電離模式;b-負離子電離模式圖4 正、負離子電離模式下G1和G2的差異代謝物聚類熱圖Fig.4 Heat map visualization of differential metabolites
|log2(FC)|值越大,說明谷氨酸棒桿菌培養液上清中的代謝物濃度變化越大。因此選取|log2(FC)|排序前25的差異代謝物構建代謝物差異倍數條形圖(附圖1,https://doi.org/10.13995/j.cnki.11-1802/ts.034392),其中17個代謝物顯著下降,僅8個代謝物上升,D-mannose 6-phosphate和5′-S-methyl-5′-thioadenosine分別為濃度下降和上升中最顯著的代謝物。

附圖1 代謝物差異倍數條形圖Fig.S1 Differential metabolites fold change bar chart
2.2.3 差異代謝物KEGG通路富集分析
以篩選得出的176種差異性代謝物為對象通過MetaboAnalyst 5.0數據庫進行生物信息學分析,匹配KEGG的數據庫獲得代謝物參與的通路信息(圖5-a)。97個下調代謝物中,34個富集到了KEGG代謝途徑,上調共79個代謝物有18個富集到了KEGG代謝途徑。KEGG通路富集表明分泌表達外源蛋白引發了代謝紊亂,特別是新陳代謝途徑(metabolic pathways)和氨基酸代謝途徑(amino acid metabolism pathways)受到影響最大,分別有39個和21個代謝物發生了顯著性變化,這些代謝通路可能在谷氨酸棒桿菌表達外源蛋白的研究中有重要意義。

a-差異代謝物KEGG富集通路圖;b-顯著差異代謝物代謝通路分析圖5 差異代謝物通路分析Fig.5 Differential metabolite pathway analysis
對|log2(FC)|數值最大的5個顯著差異代謝物進行代謝通路富集分析,除了對新陳代謝途徑產生的影響,其余主要涉及代謝通路均在表3中顯示,5個顯著差異代謝物共涉及11個代謝途徑包括碳水化合物代謝、聚糖的生物合成和代謝、核苷酸代謝及跨膜運輸等。

表3 部分顯著差異代謝物Table 3 Top5 differentially expressed metabolites in BZH001and VHH
獲得不同途徑代謝物通路的匹配信息后,根據相應的通路數據庫對系統進行檢索分析和代謝產物通路的富集分析(圖5-b),富集因子(rich factor)和對應途徑的富集程度呈正相關。P值越小,富集因子越大,表明對該通路的影響越強。通路富集分析的前5名分別是色氨酸代謝、甲烷代謝、雙組分系統、苯丙氨酸,酪氨酸和色氨酸的生物合成及酪氨酸代謝(附表3,https://doi.org/10.13995/j.cnki.11-1802/ts.034392),說明分泌表達外源蛋白導致芳香族氨基酸代謝在菌株中發生顯著變化。
3 討論
本研究采用非靶向代謝組學技術對分泌表達外源蛋白VHH的C.glutamicumBZH001(CGMCC1.15647)中代謝物進行比對和篩選,獲得關鍵差異代謝物,研究分泌表達外源蛋白對BZH001菌株自身代謝的影響,為優化其作為優良的外源蛋白表達宿主提供理論依據。
依據VIP>1和P<0.05標準,本次分析共篩選到176個差異代謝物,其中下調97個代謝物,上調79個代謝物。下調代謝物34個富集到了KEGG代謝途徑,上調代謝物中有18個富集到了KEGG代謝途徑。通過代謝通路分析發現差異表達的代謝物主要與氨基酸代謝、核苷酸代謝及碳代謝相關(附圖2,https://doi.org/10.13995/j.cnki.11-1802/ts.034392)。
D-甘露糖-6-磷酸是變化最顯著的代謝物,為對照組的0.235倍,共參與谷氨酸棒桿菌中的5條代謝通路(表3)。在氨基糖和核苷酸糖代謝、O-抗原核苷酸糖生物合成途徑中,D-甘露糖-6-磷酸在磷酸甘露糖異構酶催化的可逆作用下轉化為果糖6-磷酸,進入糖酵解后為表達外源蛋白提供能量[12]。在果糖和甘露糖代謝中,Cgl0687、Cgl0746基因編碼的磷酸甘露糖變位酶(phosphomannomutase, PMM)催化變位反應,合成鳥苷二磷酸甘露糖的前提。鳥苷二磷酸甘露糖用于多萜醇磷酸甘露糖的合成,之后參與N-糖基化[13]。在革蘭氏陰性菌空腸彎曲菌(Campylobacterjejuni)已證明N-糖基化途徑影響蛋白質穩定性、營養轉運、應激反應和抗菌素耐藥性等。分泌表達外源蛋白可能需要促進N-糖基化從而穩定胞內蛋白質同時構建適合轉運的細胞膜結構。
葉酸作為顯著差異排名前五的代謝物,其含量下降超70%,主要參與葉酸一碳庫(one carbon pool by folate)和葉酸合成(folate biosynthesis)途徑。進入機體的葉酸在經Cgl0848基因編碼的二氫葉酸還原酶(EC:1.5.1.3)作用下轉變為二氫葉酸,再經該酶轉化為5,6,7,8-四氫葉酸(5,6,7,8-tetrahydrofolate, THF)。THF是活躍的可以傳遞一碳基團的必需輔酶,參與例如甲烷代謝、氨酰tRNA生物合成等生物體內的各種反應。通過map00679分析,在C.glutamicum中THF可多路徑間接或直接轉化為5,10-亞甲基四氫葉酸,并多數為可逆反應以保障體內葉酸庫的穩定。
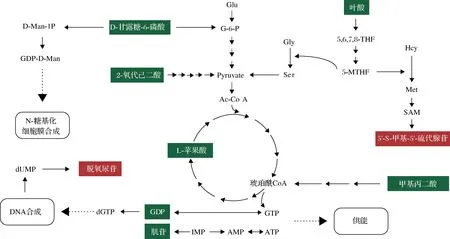
附圖2 差異代謝物代謝網絡分析Fig.S2 metabolic network analysis of differential metabolites
肌苷和GDP均參與嘌呤代謝途徑。肌苷減少為對照組的0.32,首先在產生路徑中次黃嘌呤核苷酸(inosinemonphosphate, IMP)減少向肌苷轉化并且轉向AMP,更易轉為ATP為菌株提供能量。其次,肌苷增加向次黃嘌呤的消耗,經Cgl2697基因所編碼的酶被催化為IMP,最終促進腺苷的生成。GDP涌向dGTP方向,用于DNA合成,同時轉化為GMP釋放高能磷酸基團和能量,供菌株使用。
5′-S-甲基-5′-硫代腺苷在G2組中上升至3.6倍。在由Cgl1603編碼的S-腺苷甲硫氨酸合成酶催化下,甲硫氨酸獲得來自ATP的能量形成擁有高能甲硫鍵的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM),再經亞精胺合成酶(Cgl2702)轉化為5′-S-甲基-5′-硫代腺苷。分析認為甲硫氨酸合成酶催化同型半胱氨酸(L-homocysteine, Hcy),在5-甲基四氫葉酸提供甲基條件下,可形成甲硫氨酸用于菌株代謝和滿足蛋白表達需要。結合葉酸含量顯著降低,大量甲硫氨酸合成,由于總量上升導致甲硫氨酸降解途徑加劇從而積累5′-S-甲基-5′-硫代腺苷[14]。
富集顯著性分析中多條氨基酸相關路徑得分靠前,說明外源表達分泌蛋白不僅影響C.glutamicumBZH001能量代謝還與氨基酸合成、利用緊密相關。其中最主要的是芳香族氨基酸的代謝[15],包括色氨酸代謝、酪氨酸代謝及苯丙氨酸、酪氨酸和色氨酸的生物合成[16]。
在苯丙氨酸、酪氨酸和色氨酸的生物合成途徑中,分支酸處流向色氨酸途徑或被催化生成預苯酸,后者作為前體參與L-苯丙氨酸及L-酪氨酸的合成。有研究表明表達異源蛋白可以使生物體內產生D型氨基酸[17],G1和G2組中均有異源質粒的導入,故可檢測到D-苯丙氨酸的存在。D-苯丙氨酸經D-氨基酸脫氫酶氧化為苯丙酮酸,D-3-苯乳酸可逆轉化為苯丙酮酸,苯丙酮酸作為前體經組氨醇磷酸氨基轉移酶反應為L-苯丙氨酸。L-苯丙氨酸量增加60%有助于氨基酸累積[18-19],形成疏水區保護胞內蛋白質[20]從而穩定細胞內環境以抵抗表達外源蛋白造成的脅迫。
在色氨酸代謝中,L-色氨酸、2-氧代己二酸、5-羥色胺、5-羥基吲哚-3-乙酸4個代謝物下降,吲哚、L-5-羥基色氨酸和3-甲基吲哚3個代謝物顯著上升,說明色氨酸代謝途徑在分泌外源蛋白過程中發揮重要作用。其中L-色氨酸下降[21]影響細胞膜的通透性,從而加快物質吸收及蛋白分泌。2-氧代己二酸(2-ketoadipic acid)與未表達外源蛋白菌株相比下調57%,從代謝通路map00380分析,此步驟所消耗的2-氧代己二酸明顯增多,經過5步反應生成乙酰輔酶A參與糖酵解進而參與能量代謝。同時,三羧酸循環途徑中L-蘋果酸下調,說明三羧酸循環活躍,為菌株提供更多能量。分泌表達外源蛋白導致BZH001菌株能量消耗大量增加,需要上調中心代謝以滿足菌株生長和生產的需要。
與核苷酸代謝相關的7個差異代謝物中,在嘧啶代謝相關3個差異代謝物中,L-二氫乳清酸(L-dihydroorotic acid)和甲基丙二酸(methylmalonate)均顯著下降。在嘧啶生物合成過程中有多種嘧啶環中間產物生成,L-二氫乳清酸作為最先生成的物質在脫氫后轉化為乳清酸。通過路徑分析(map00240)其與精氨酸代謝、丙氨酸、天冬氨酸和谷氨酰胺代謝以及纈氨酸、亮氨酸和異亮氨酸代謝相關[22],菌株大量表達外源蛋白時大量氨基酸流向蛋白合成路徑導致相關水解產物下降。在嘌呤代謝中,脫氧尿苷(2-deoxyuridine)兩倍上升,dUMP在5′-核苷酸酶和2′,3′-環腺苷酸-3′-磷酸二酯酶等酶簇催化下釋放磷酸基團和能量,同時尿苷轉化為脫氧尿苷。在N-乙酰葡萄糖胺-1-磷酸尿苷酰轉移酶(N-acetylglucosamine-1-phosphate uridyltransferase, GlmU)作用下尿苷被催化成為尿苷二磷酸-N-乙酰葡萄糖胺(UDP-N-acetylglucosamine, UDP-GlcNAc),此物質在革蘭氏陽性菌中有重要作用,主要涉及肽聚糖生物合成[23],肽聚糖是谷氨酸棒桿菌細胞壁的主要成分之一,推測蛋白分泌影響細胞壁的生物合成,改變肽聚糖層交聯網絡的孔隙進而造成細胞壁通透性的變化。
綜上分析,分泌表達外源蛋白影響C.glutamicumBZH001的代謝,特別是能量的代謝通路和氨基酸的代謝通路。糖酵解和三羧酸循環加劇以提供能量滿足BZH001表達外源蛋白的需求以及脅迫下的自身生長。氨基酸代謝的改變在滿足提供外源蛋白合成原料之外還影響胞內環境穩定性和細胞壁合成,有利于外源蛋白的分泌。
4 結論
本研究代謝組分析結果表示,當C.glutamicumBZH001分泌表達外源蛋白VHH時需要大量的能量同時吸引氨基酸前往合成路徑,具體表現為中心代謝加劇以滿足生產和生長的雙向要求,氨基酸大量流向蛋白合成方向導致嘧啶代謝中與氨基酸相關的代謝物顯著下調。C.glutamicumBZH001菌株通過調節體內三羧酸循環、核苷酸代謝和氨基酸代謝等提高能量供給以應對外源蛋白合成及分泌造成的細胞應激反應,利用其他組學聯合分析,理清關鍵性差異代謝物并進行驗證性研究,為后續改造BZH001菌株成為更好的外源蛋白表達宿主提供理論支持。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
