馬尾藻多糖拮抗LPS誘導的巨噬細胞極化及鐵死亡①
2023-11-13 09:30:10袁慧情胡佳敏劉思溢侯鑒基吳科鋒吳斌華廣東醫科大學湛江524023
中國免疫學雜志 2023年10期
袁慧情 胡佳敏 劉思溢 侯鑒基 吳科鋒 羅 輝 吳斌華 (廣東醫科大學,湛江 524023)
馬尾藻(Sargassum)是褐藻的一種,廣泛分布于 我國東海、南海,因其富含多種營養成分且具有一定的有益功效,廣泛用于食品和醫藥研究等[1]。馬尾藻多糖(sargassum polysaccharide,SP)是其主要成分,具有豐富的生物活性,包括抗氧化、抗癌、抗糖尿病、抗氧化等[2-5]。
目前SP 調控巨噬細胞抗炎作用的研究多集中于分析其影響的具體信號通路中,包括核因子-κB(nuclear factor kappa-B,NF-κB)、絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、Janus 激酶/信號轉導與轉錄激活子(Janus kinase/signal transducer and activator of transcription,JAK/STAT)等 炎癥通路[6],例如,SP能通過下調p38和ERK的磷酸化以及NF-κB p50和p65的移位而抑制MAPK和NF-κB信號激活,從而發揮抗炎作用[7]。研究發現,SP能提高單核巨噬細胞的吞噬能力,促進巨噬細胞增生和淋巴小結形成[8]。因此,選用小鼠單核巨噬細胞RAW264.7進行研究。巨噬細胞是一種位于組織內的白血球,源自單核細胞,在不同藥物刺激下極化為不同的功能表型[9-11]。由LPS 刺激激活為M1 表型,表現促炎活性,發揮促炎抑菌作用[12-13]。細胞由IL-4 刺激活化為M2 表型,表現抑炎活性,有利于促進組織生長和愈合[14-16]。
除了誘導巨噬細胞極化以外,脂多糖(lipopolysaccharide,LPS)還能誘導細胞鐵死亡[17]。鐵死亡是一種鐵依賴性、區別于細胞凋亡、壞死、自噬的新型細胞程序性死亡方式[18]。不同于其他形式調控細胞死亡的分子特征,其用于描述一種由活性氧積累引起的依賴鐵的細胞死亡調控形式[19-20]。研究發現,LPS 除了引起巨噬細胞極化以外,還可劑量依賴性地增加細胞內總鐵含量,Fer-1 可使其恢復[21];鐵死亡激活劑Erastin 可以通過激活STAT3 促進巨噬細胞M2 型極化增強上皮性卵巢癌的轉移能力[22];此外,鐵死亡還能通過釋放和攝取致癌KRAS 蛋白促進腫瘤相關巨噬細胞向M2 型極化,因此,巨噬細胞極化和鐵死亡之間存在密切聯系[23]。本研究重點探討SP 對LPS 誘導的巨噬細胞極化和鐵死亡這兩個因素的影響。
1 材料與方法
1.1 材料
1.1.1 多糖來源 從硇洲島海岸采集的SP,用3.5%NaCl 溶液清洗,冷凍干燥,用水/乙醇(3∶7)在室溫下提取24 h,攪拌,過濾。隨后,提取物在減壓下濃縮,殘渣冷凍干燥,得到粗粉[24]。該海藻的憑證標本保存于廣東醫科大學海洋醫藥研究院。
1.1.2 試劑及儀器 10%胎牛血清、cDNA 合成試劑盒[(賽默飛世爾科技(中國)有限公司,2279604CP、01071349)];DMEM 完全培養基[(賽默飛世爾科技(中國)有限公司,8120365)];LPS、CCK-8 試劑、PMSF、ROS 檢測試劑盒(碧云天生物技術有限 公 司,022621210706、073020200930、ST506-2、011521210621);AG RNAex Pro RNA 提取試劑(湖南艾科瑞生物工程有限公司,A3A0031)。
Epoch 酶標儀(美國博騰儀器有限公司,Epoch);傅里葉變換紅外光譜儀(日本島津公司,IRAffinity-1);基因擴增儀(杭州柏恒科技有限公司,GE9612T);熒光定量基因擴增儀(德國ANALYTIKJENA,qTOWER384G);超分辨轉盤共聚焦顯微鏡(Olympus,Ixplore SpinSR)。
1.2 方法
1.2.1 細胞培養 巨噬細胞株小鼠RAW264.7 來自本實驗室。細胞培養于含抗生素及10%胎牛血清的DMEM 完全培養基中,于37 ℃、5%CO2的孵箱中培養。細胞種板貼壁后,隨機分為4組:①空白對照組(blank control,BC 組):細胞用DMEM 完全培養基培養,未加任何其他材料;②不同濃度SP組:細胞以含不同濃度SP 的DMEM 完全培養基進行培養;③LPS 組:使用含終濃度為1 μg/ml LPS 的DMEM 完全培養基培養細胞;④SP+LPS 組:使用含有終濃度為1 μg/ml LPS 和不同濃度SP 的DMEM 完全培養基。細胞按各組要求加入相應培養基后,繼續培養24 h,進行后續實驗。
1.2.2 紫外吸光度檢測 采用Epoch 酶標儀在200~400 nm 對SP 進行紫外吸光度分析,以判斷其純度及是否存在蛋白污染。
1.2.3 紅外分光光度法分析SP結構 采用傅里葉變換紅外光譜儀對SP進行紅外吸光度掃描檢測。
1.2.4 CCK-8 法檢測RAW264.7 細胞增殖情況
取對數生長期細胞計數,以5 000 個/孔均勻地接種于96 孔板,細胞貼壁后,分別用不同濃度的LPS、SP予以相應處理;同時設置對照組,每個樣品6個重復孔。放入孵箱繼續培養24 h 后,每孔加入10 μl CCK-8 溶液,操作過程避光。放入孵箱繼續孵育2 h,避光輕微振蕩,待培養板內結晶完全溶解后,酶標儀于450 nm 處測定OD 值,并根據下列公式計算細胞存活率。細胞存活率(%)=(實驗孔OD-空白孔OD)/(對照孔OD-空白孔OD)×100%
1.2.5 RT-qPCR RT-qPCR 試驗參考SUH 等[25]的實驗,取預處理、裝于6 孔板中的細胞,采用AG RNAex Pro RNA 提取試劑抽提細胞總RNA。采用cDNA 合成試劑盒合成cDNA,以基因擴增儀進行逆轉錄操作,收集逆轉錄產物,使用熒光定量基因擴增儀進行PCR 反應,PCR 反應條件:95 ℃ 30 s,95 ℃5 s,60 ℃ 30 s,40 個循環。以GAPDH 作為內參,讀取各樣品Ct值,結果用2-ΔΔCt表示。引物序列見表1。

表1 引物序列Tab.1 Primer sequences
1.2.6 Western blot 檢測 收集藥物干預后的細胞,使用加入PMSF 的細胞蛋白裂解液,冰上裂解,4 ℃、14 000 g離心20 min后,將蛋白樣品與5×上樣緩沖液按4∶1 體積混合,100 ℃煮沸10 min。通過SDS-PAGE電泳后,轉移至PVDF膜,用5%脫脂奶粉封閉2 h 后,加入一抗(1∶1 000)4 ℃過夜孵育,TBST 洗滌3 次。二抗孵育1 h,TBST 洗膜3 次后,用免疫印跡化學發光試劑(ECL)顯影。抗體來源見表2。
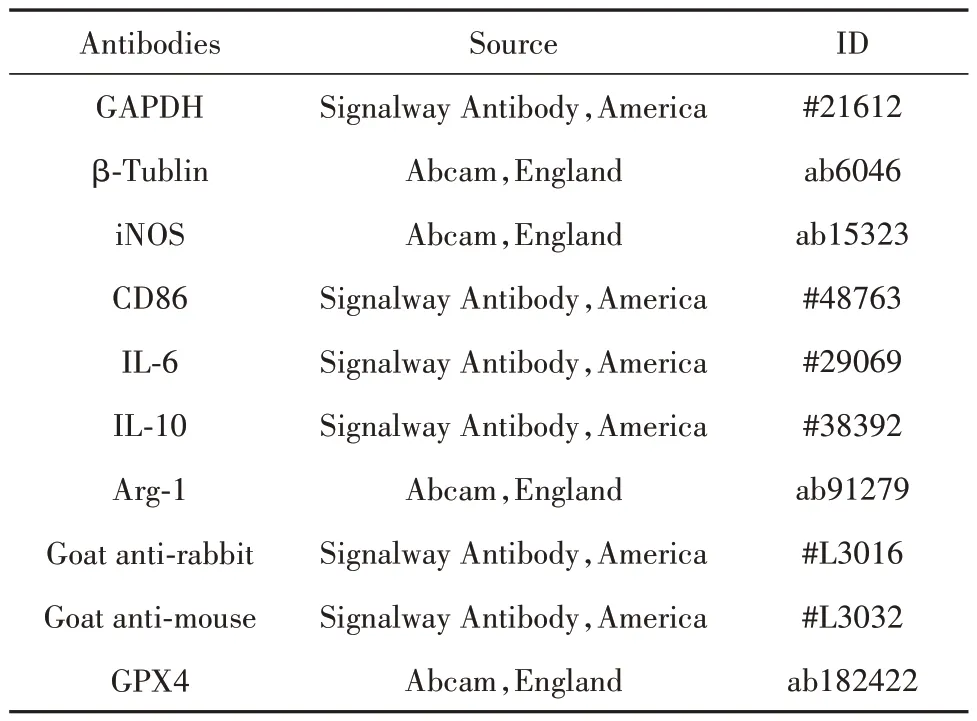
表2 抗體來源Tab.2 Antibody source
1.2.7 ROS 檢測 實驗操作參考HUANG 等[26]方法。在細胞中加入1 ml 含有稀釋DCFH 的PBS。實驗后37 ℃下孵育20 min,每隔3 min混合1次。然后用無血清DMEM培養基洗滌細胞3次,最后以500 μl的PBS重懸。在超分辨轉盤共聚焦顯微鏡下觀察拍照,用Image J 計算出綠色熒光強度并對每組數據進行統計分析。
1.3 統計學處理 實驗結果采用GraphPad Prism軟件進行數據分析,兩組間比較采用獨立樣本t檢驗,多組間比較采用單因素方差分析,以P<0.05 為差異有統計學意義。
2 結果
2.1 紅外吸光度檢測結果 將分離的SP進行紅外光譜分析,結果顯示,在3 087.83、1 520.14、1 183.61、1 061.94 cm-1處均顯示吸收峰,分別為-OH、C=O、CO 和C-O-C 的特征吸收峰。C=O 說明其中含有醛基,C-O-C 是吡喃糖的特征吸收峰,因此,可以判定SP為吡喃型多糖。見圖1。
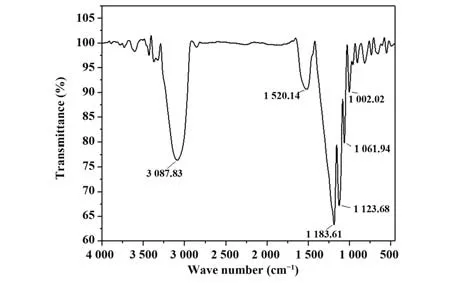
圖1 SP紅外吸收掃描結果Fig.1 Infrared absorption scanning results of SP
2.2 紫外吸光度檢測結果 紫外吸光度顯示在280 nm 處無吸收峰,提示樣品中無蛋白類物質,見圖2。
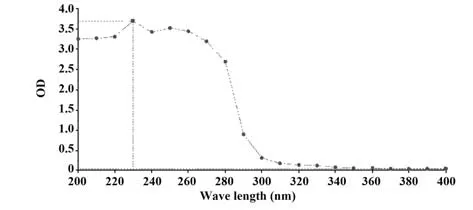
圖2 SP紫外吸收掃描結果Fig.2 Ultraviolet absorption scanning results of SP
2.3 SP 對LPS 處 理 的RAW264.7 細 胞 增 殖 的 影響 采用CCK-8 法考察細胞經不同濃度SP 和不同濃度LPS 處理RAW264.7 細胞后增殖活性的變化,結果見圖3。圖3A顯示,與BC組比較,25~200 μg/ml組細胞增殖無顯著影響(P>0.05),但400 μg/ml SP 對RAW264.7 組細胞的增殖率為113.5%,與BC 組比較顯著升高(P<0.05),提示400 μg/ml SP 本身對RAW264.7細胞有促增殖作用。為避免多糖的促增殖功能,采用0、50、100、200 μg/ml 進行后續實驗。采用不同濃度LPS 處理RAW264.7 細胞(圖3B),觀察LPS 對巨噬細胞增殖的影響,結果發現1、10、100 μg/ml LPS 對RAW264.7 細胞增殖有明顯抑制作 用,抑 制 率 分 別 為(89.83±5.00)%、(86.26±1.19)%、(2.04±4.24)%,與BC 組比較有明顯抑制作用(均P<0.05),說明1、10、100 μg/ml LPS 對巨噬細胞增殖具有明顯抑制作用。參考文獻[27],在后續實驗中采用1 μg/ml LPS進行實驗。
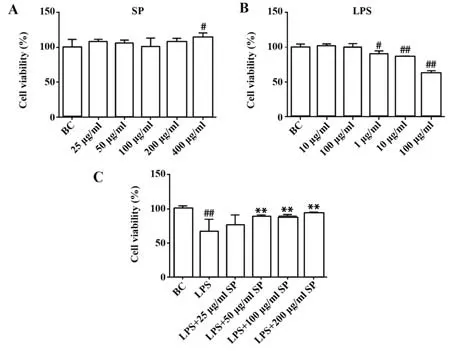
圖3 SP及LPS對RAW264.7細胞增殖的影響Fig.3 Effects of SP and LPS on proliferation of RAW264.7 cells
進一步觀察SP 對LPS 誘導的巨噬細胞損傷的保護情況(圖3C),結果顯示,25 μg/ml 濃度組細胞增殖抑制率為(76.05±15.41)%,與LPS 組比較差異無統計學意義(P>0.05),而50、100、200 μg/ml 濃度組 細 胞 活 力 分 別 為(88.86±2.64)% 、(89.92±2.14)%、(94.29±1.47)%,與LPS 組比較顯著升高(均P<0.05)。這一結果說明高濃度的多糖對LPS誘導的細胞增殖抑制具有顯著保護作用。
2.4 SP對巨噬細胞極化的調控
2.4.1 SP 對巨噬細胞極化相關標志物轉錄水平的影響 qPCR 結 果顯示:25、100、200 μg/ml SP 組CD163 轉錄水平分別是BC 組的0.55 倍(P>0.05)、2.88 倍(P<0.01)、11.90 倍(P<0.01);IL-10 轉錄水平分別是BC 組的2.67 倍(P>0.05)、6.88 倍(P<0.01)、4.44 倍(P<0.01);Arg-1 轉錄水平分別是BC組的0.90 倍(P>0.05)、6.16 倍(P<0.01)、12.17 倍(P<0.01);由此得出一定濃度的SP能使RAW264.7細胞的IL-10、CD163和Arg-1表達量升高。見圖4。
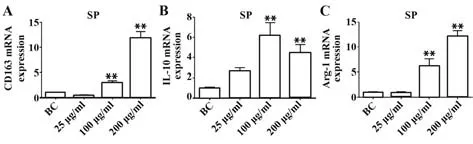
圖4 SP對IL-10、CD163和Arg-1的mRNA表達的影響Fig.4 Effect of SP on mRNA expressions of IL-10,CD163 and Arg-1
2.4.2 SP 對巨噬細胞極化相關標志物的蛋白表達水平的影響 如圖5 所示,與BC 組比較,iNOS、CD86 的蛋白表達量均隨著SP 劑量的增加而降低,而IL-10在加入SP后蛋白表達量增加(P<0.05)。結果與圖4 的趨勢一致。表明SP 單獨作用小鼠巨噬細胞,可能導致細胞向M2型極化。
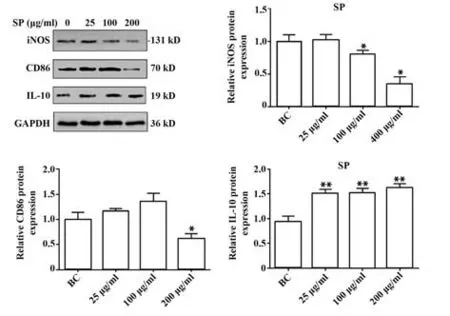
圖5 SP對iNOS、IL-10、CD86蛋白表達的影響Fig.5 Effects of SP on protein expressions of iNOS,IL-10 and CD86
2.4.3 SP 拮抗LPS 誘導的RAW264.7 細胞極化進一步觀察SP 能否拮抗LPS 誘導的巨噬細胞M1 型極化,RT-qPCR 結果(圖6A)顯示,在經過LPS 單獨處理后,M1 極化指標CD80、iNOS 和TNF-α 的轉錄水平均顯著升高(P<0.01),經過LPS 和SP 處理后,相較于LPS 組各組均顯著降低(P<0.05)。LPS 組CD80 的 轉 錄 水 平 升 高 為BC 組 的70.78 倍(P<0.01),LPS 和25、100 μg/ml SP 處理以后,降低為BC組的37.46、44.40 倍(P<0.01);LPS 組iNOS 的轉錄水平升高到BC 組的33.69 倍(P<0.01),經過LPS 和25、100 μg/ml SP 處理以后,降低為BC 組的11.96、13.29 倍(P<0.01);LPS 組TNF-α 的轉錄水平升高為BC 組的14.14 倍(P<0.05),LPS 和25、 100 μg/ml SP 處理以后,降低為BC 組的8.70、8.53 倍(P<0.05)。M2 極 化 標 志 物CD163 經 過LPS 和25、100 μg/ml SP 處理后其轉錄水平顯著升高為BC 組的14.85、55.84倍(P<0.05)。
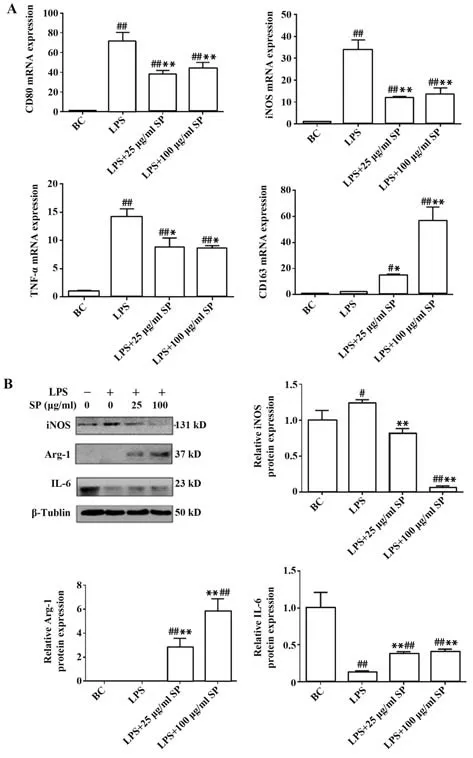
圖6 SP對LPS誘導的RAW264.7細胞極化的影響Fig.6 Effects of SP on LPS-induced polarization of RAW264.7 cells
Western blot 結果顯示(圖6B),在經過LPS 單獨處理以后,M1 極化相關標志物iNOS 的蛋白表達水平顯著升高(P<0.05),經過LPS和SP處理以后顯著降低(P<0.05)。BC 組和LPS 組的M2 極化標志物Arg-1 表達均不明顯,因此無法進行比較,但經過LPS 和SP 處理后,其蛋白表達水平均增高并存在劑量依賴性(P<0.05)。M2 極化標志物IL-6 經過LPS單獨處理以后,蛋白表達水平顯著降低(P<0.01),經過LPS 和SP 共同處理,與LPS 組相比IL-P 表達水平顯著升高(P<0.01)。SP 能拮抗LPS 誘導的巨噬細胞M1極化。
2.5 SP拮抗LPS誘導的細胞鐵死亡
2.5.1 SP 對LPS 損傷RAW264.7 細胞內ROS 的影響 如圖7 所示,與BC 組相比,1 μg/ml LPS 損傷細胞24 h 后細胞內的ROS 水平明顯升高(P<0.01),而SP處理后的細胞內ROS相較于LPS組明顯降低(P<0.01),并呈劑量依賴性。
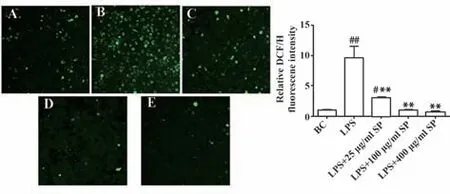
圖7 DCFH-DA熒光探針檢測細胞內ROS水平Fig.7 Intracellular ROS level was detected by DCFH-DA fluorescent probe
2.5.2 SP 對巨噬細胞內鐵死亡相關基因GPX4 表達的影響 Western blot實驗分析細胞中GPX4蛋白的表達,結果如圖8 顯示,與BC 組比較,LPS 組中的GPX4蛋白表達水平顯著降低(P<0.01);經25 μg/ml馬尾藻多糖處理后,GPX4 蛋白表達略微升高,但差異無統計學意義(P>0.05),劑量加大后,400 μg/ml SP 與LPS 組相比,GPX4 蛋白表達顯著升高,差異有統 計 學 意 義(P<0.01)。SP 能 拮 抗LPS 引 起 的RAW264.7細胞GPX4蛋白表達水平升高,ROS水平升高。因此,SP能拮抗LPS誘導的細胞鐵死亡。
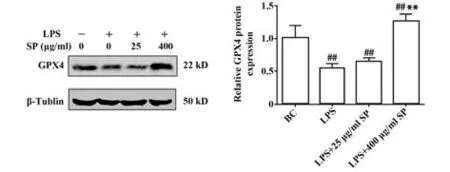
圖8 LPS和SP對GPX4蛋白表達的影響Fig.8 Effects of LPS and SP on expression of GPX4 protein
3 討論
近年來,海洋藥物研究一直是國內外研究的熱點,海洋生物由于生活環境的不同,與其他物質相比,通常具有特殊化學結構和獨特生物活性,比如抗氧化、抗衰老、抗炎、抗腫瘤、抗菌和神經保護等作用[28-29]。其中,海洋藥物中許多萜類、生物堿、多糖類和多肽類等均具有抗炎活性,海藻多糖通常通過調控NF-κB、MAPK、JAK/STAT等信號通路發揮其抗炎作用,對海藻多糖的研究可以用于新型抗炎藥的研發或者作為新型抗炎藥的前體[30-31]。
巨噬細胞M2 極化有促進細胞生長和組織修復的功能[32]。前期已有研究證明石斛多糖、枸杞多糖能通過促進RAW264.7 細胞增殖作用來發揮其抗炎活性[33-34],基于此,本實驗采用CCK-8法觀察SP各濃度對RAW264.7 細胞增殖的影響,選取對細胞增殖無明顯抑制作用的SP(25、50、100、200 μg/ml) 開展研究,發現各濃度SP促進細胞增殖。研究結果表明,1 μg/ml LPS 對細胞增殖有顯著影響,因此在后續實驗中,均選取1 μg/ml LPS 進行操作。此外,檢測SP 對LPS 誘導巨噬細胞的影響。以1 μg/ml LPS刺激RAW264.7 細胞24 h,再用不同濃度的SP 處理細胞,50、100、200 μg/ml SP 能顯著拮抗LPS 引起的細胞活力下降。
結合其他“多糖通過調控小鼠單核巨噬細胞極化來發揮抗炎作用”的研究,比如黑靈芝多糖可顯著降低巨噬細胞吞噬能力并抑制IL-1β、NO 和ROS的生成,由此得出結論黑靈芝多糖能抑制RAW264.7 細胞的M1 極化而發揮抗炎作用[35]。本研究基于M1、M2 巨噬細胞活化相關的表面膜蛋白iNOS、CD80、TNF-α、IL-10、Arg-1、CD206 等作為活化指標,qPCR、Wstern blot 結果均顯示:當單獨給予SP 刺激RAW264.7 細胞,特異性M2 亞型巨噬細胞的膜蛋白高表達CD206,IL-10、Arg-1 等,而iNOS、CD80、TNF-α 等特異性M1 亞型巨噬細胞各指標呈低表達,SP 通過活化巨噬細胞,分泌具有生物活性的細胞因子參與細胞的免疫功能,從而起到調節機體免疫的效應[36]。
研究發現,LPS 能誘導氧化應激、非血紅素鐵和血紅素水平升高[37]。Ferrostatin-1 能通過抑制鐵死亡減輕LPS引起的急性肺損傷[38]。鐵死亡產生的主要原因是細胞內脂質活性氧生成與降解失衡,脂質活性氧堆積,導致細胞氧化性死亡[39]。GPX4、Nrf2等可以通過限制細胞對鐵的攝入和減少ROS 的產生作為負性調節因子發揮抑制鐵死亡作用[40]。本實驗用LPS 誘導巨噬細胞,發現GPX4 蛋白表達降低,ROS產生增加,因此得出LPS能誘導巨噬細胞鐵死亡[41]。再用不同濃度的馬尾藻多糖處理LPS誘導過的RAW264.7 細胞,發現馬尾藻多糖GPX4 的表達相比較于LPS 組顯著降低,ROS 的產生也明顯下降,充分說明馬尾藻多糖能通過抑制ROS 的產生來拮抗LPS誘導的RAW264.7細胞鐵死亡。
總之,SP 能促進巨噬細胞增殖,并保護LPS 引起的RAW264.7 細胞損傷。此外,它還通過抑制iNOS、TNF-α、CD80 和促進IL-10、Arg-1、CD206 蛋白及轉錄水平的表達,從而逆轉LPS 誘導的RAW264.7 細胞向M1 型極化,其作用機制與抑制iNOS、TNF-α、CD80 和促進IL-10、Arg-1、CD206 蛋白及轉錄水平的表達有關。LPS 能通過抑制GPX4 蛋白表達、促進ROS 產生來誘導RAW264.7 細胞鐵死亡,SP能逆轉這一現象。由此說明SP能拮抗LPS誘導的RAW264.7細胞鐵死亡,提示SP在抗炎方面具有巨大的開發潛力。為此,將進一步探討SP炎癥保護作用的具體分子機制,尋找其他更有效的下游分子靶點及信號調控通路,為相關藥理研究和藥物開發提供理論依據。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
火花(2019年12期)2019-12-26 01:00:28
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2019年12期)2019-05-21 02:55:32
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
學苑創造·A版(2015年11期)2016-01-14 09:03:27
實驗流體力學(2011年5期)2011-01-14 01:25:28