一株彎曲固氮菌對多環芳烴的降解特性研究
2023-12-02 14:18:50張宏宏葉晨張曉昀孟恬王黎明黃玉屏
湖北工業大學學報 2023年1期
張宏宏 葉晨 張曉昀 孟恬 王黎明 黃玉屏
[摘 要]微生物修復在多環芳烴(polycyclic aromatic hydrocarbons, PAHs)污染治理方面有著優良特性。為了獲得高效降解PAHs的微生物,以熒蒽為唯一碳源進行篩選,分離得到一株高效降解細菌,命名為CE3。經16S rDNA序列分析法鑒定,菌株CE3屬于彎曲固氮菌屬(Azoarcus),這是首次發現該屬菌株對高分子量PAHs具有降解能力。菌株CE3除了能降解熒蒽,還能降解苯、菲、芘、熒蒽、苯并[a]蒽、3-4苯并芘和苯并[b]熒蒽,且對熒蒽和芘的降解率較高,均達到50%以上。分析各種條件對CE3菌降解熒蒽和芘的影響,發現CE3菌能在較廣的溫度和pH范圍降解熒蒽或芘,并且添加酵母提取物、蔗糖和果糖可使CE3菌降解能力提高。
[關鍵詞]多環芳烴;熒蒽;芘;降解;彎曲固氮菌
[中圖分類號]Q939.99; X53[文獻標識碼]A
多環芳烴(polycyclic aromatic hydrocarbons, PAHs)是一類含有兩個及兩個以上苯環的有機化合物,具有遺傳毒性、致突變性、致癌性、生物蓄積性和化學穩定性[1-3],是我國農用和居住用地土壤評估的重要指標之一。根據環數的不同,PAHs可以分為高分子量PAHs(4環及以上)和低分子量PAHs(2~3環),且高分子量PAHs比低分子量PAHs具有更高的穩定性、更強的致癌性、致畸性和致突變性,對人類健康和生態環境產生的危害更大[4-5]。我國大部分地區表層土壤中整體上都含有一定量的PAHs,并多處于中度污染水平,其中4環的PAHs熒蒽和芘含量較高,2環的PAHs含量較低[6]。例如本研究的供試土壤采樣區湖北黃石新冶鋼有限公司東鋼廠區,其焦化車間受到的多環芳烴污染十分嚴重,其中大部分為高分子量多環芳烴污染物。由于在環境中低分子量的PAHs比高分子量的容易降解,因此對高分子量PAHs的污染治理非常重要。在PAHs污染治理方法中,微生物治理方法因其環境友好、成本低、高效等特點,成為了PAHs污染治理的熱點[7]。然而,目前分離到的降解高分子量PAHs菌株非常有限,對其降解機制所知甚少。為此,迫切需要分離更多的高分子量PAHs降解菌,探明它們對高分子量PAHs的代謝機制,從而加快微生物在環境污染治理和修復中的應用。
1 材料與方法
1.1 實驗材料
1.1.1 土壤樣品的采集 本研究所用土壤樣品采自湖北省黃石市新冶鋼有限公司東鋼廠焦化車間,取距表層10 cm下的土壤。
1.1.2 培養基
1)無機鹽培養基 (1 L):338.8 mg KH2PO4、234.0 mg (NH4)2SO4、100.0 mg Na2CO3、3.9 mg CaCl2、59.3 mg MgSO4·7H2O、890.7 mg Na2HPO4·12H2O、0.3 mg FeSO4·7H2O,1 mL 微量元素母液。
2)微量元素母液(1 L):1500 mg FeCl2·4H2O、190 mg CoCl2·6H2O、100 mg MnSO4·7H2O、70 mg ZnCl2、24 mg NiCl2·6H2O、24 mg NaMoO4·2H2O、6 mg MnCl2·4H2O、2 mg CuCl2·2H2O。
1.1.3 試劑與儀器 除上述培養基配方中的化學試劑(購自國藥集團化學試劑有限公司)外,本研究試劑主要有:菲、熒蒽、芘、3-4苯并芘、苯并[a]蒽、苯并[b]熒蒽、丙酮、二氯甲烷(分析純)、甲醇(色譜純)(購自阿拉丁工業公司)。本研究所用實驗儀器包括島津UV2500型分光光度計、Agilent 1200高效液相色譜儀、Ultimate PAH高效液相色譜柱(5 μm, 250 mm×4.6 mm)等。
1.2 實驗方法
1.2.1 多環芳烴降解菌的分離純化 稱取10 g混合土壤樣品,加入90 mL無菌水和適量玻璃珠振蕩3 h;靜置待土壤沉降后,取10 mL上清液接入加有熒蒽的90 mL無機鹽培養基中,30℃、180 r/min培養7 d。每次均按10%的體積比移取培養液接入新鮮的液體無機鹽培養基中,進行富集培養,共重復5次。富集培養時培養基中熒蒽的濃度依次為10、20、40、60、80 mg/L。之后,將培養液進行稀釋涂布,并將單菌落在平板上進一步劃線分離純化。
1.2.2 降解菌的鑒定
1)菌株形態特征的觀察:a)挑取單菌落分別在LB平板和含有熒蒽的無機鹽平板上劃線,30℃培養,觀察菌落形態,并進行革蘭氏染色。b)將菌株接入含有熒蒽的無機鹽培養基中,30℃、180 r/min培養,觀察其在液體無機鹽培養基中的生長狀態。
2) 菌株的分子生物學鑒定:用細菌 16S rDNA 通用引物 ( 27F: 5-CAGCGGTACCAGAGTTTGATCCTGGCTCAG-3和1492R: 5-CTCTCTGCAGTACGGCTACCTTGTTACGACTT-3)進行PCR擴增,將PCR產物送公司測序。將序列信息輸入NCBI 網站,經 Blast 程序與 GenBank 中已有的核酸序列進行序列同源性比對分析,并用MEGA 11.0軟件采用Neighbor-Joining法構建菌株的系統發育樹。
1.2.3 多環芳烴濃度的測定 將活化之后的菌液8000 r/min離心5 min,棄上清,用無機鹽培養基洗滌2次,再用無機鹽培養基將菌體重懸;將菌液OD600調至1.0,按所需的接種量轉接于50 mL加有多環芳烴為單一碳源的無機鹽培養基中,30℃、180 r/min培養7 d。
1)分光光度法檢測多環芳烴
a)配制各種多環芳烴溶于甲醇的標準品,用島津UV2500分光光度計做全波段掃描,來確定每種多環芳烴對應的最大吸收波長。
b)分別在各種多環芳烴的最大吸收波長處測定不同濃度梯度標準樣品的吸光度,以吸光度值為縱坐標,多環芳烴的濃度為橫坐標,繪制標準曲線。測定加標回收率,計算變異系數,分析重復性。
c)向50 mL 培養液中加入25 mL 二氯甲烷,振蕩萃取30 min,之后倒入分液漏斗中,靜置15 min,分層后收集下層的有機相,上層的水相按同樣的方法連續萃取2 次。
d)合并3次萃取獲得的有機相,離心去掉有機相中殘留的水分,并測量有機相的體積。
e)測定于上述有機相樣品在最大吸收波長下的吸收值,并根據標準曲線計算多環芳烴的濃度和降解率。
2)高效液相色譜法測定熒蒽和芘
a)繪制熒蒽和芘的標準曲線
配制不同濃度梯度的熒蒽和芘的標準樣品,上機分析。分析的色譜條件為:流動相為V(甲醇)∶V(水)=9∶1,紫外檢測器室溫檢測,設置流速為1 mL/min,進樣量為30 μL,檢測波長為254 nm,分析時間為30 min。以峰面積為縱坐標,濃度為橫坐標分別繪制熒蒽和芘的標準曲線。
b)培養液中加入等體積的二氯甲烷萃取,振蕩萃取30 min,倒入分液漏斗中,靜置15 min,分層后收集有機相,水相重復萃取2次。
c)將有機相離心,去掉殘留的水分后,用旋轉蒸發器進行旋轉蒸發,水浴溫度為40℃;向蒸干后的圓底燒瓶中加入50 mL色譜純的甲醇,充分溶解多環芳烴。
d)取1 mL上述多環芳烴甲醇溶液,過濾除去雜質,制得待測樣品。將待測樣品上機檢測,根據檢測結果和標準曲線計算熒蒽及芘的濃度和降解率。
2 結果與討論
2.1 菌株CE3的形態特征
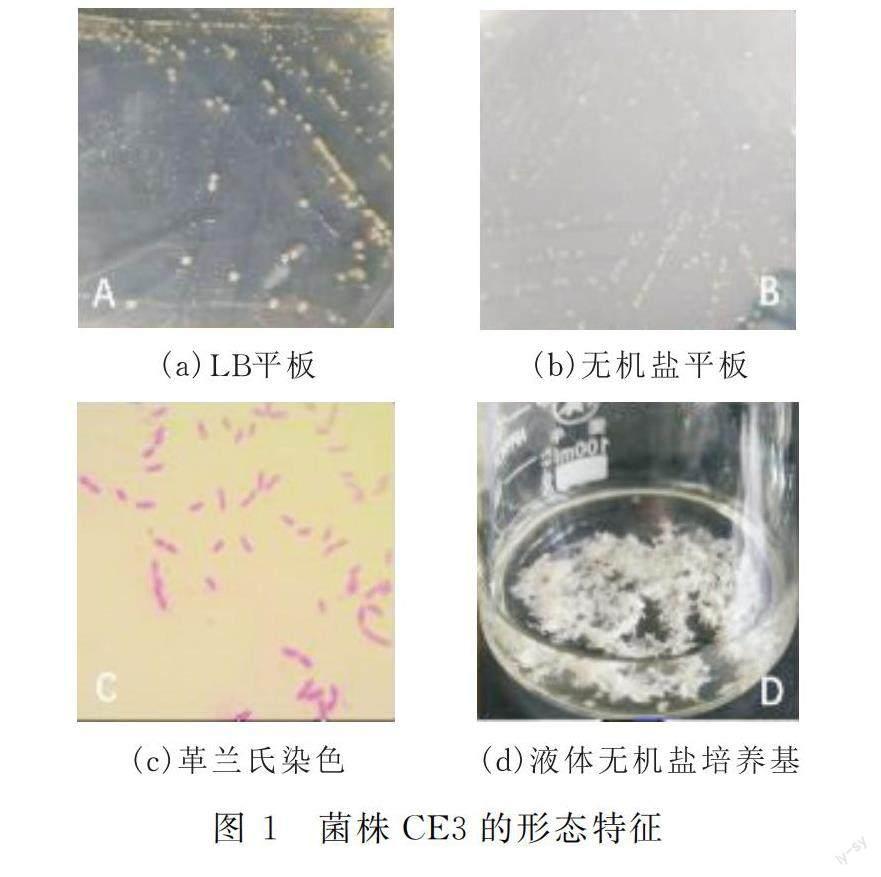
通過逐次提高培養基中的熒蒽濃度,并用平板劃線分離純化,篩選得到一株高效降解熒蒽的菌株,編號CE3。菌株CE3在LB平板(圖1a)和無機鹽平板(圖1b)上的菌落呈規則圓形,透明,淡黃色,凸起,表面光滑濕潤,質地粘稠易挑起。菌株CE3為革蘭氏陰性菌,菌體呈略彎曲的棒狀(圖1c)。菌株CE3在含有熒蒽的無機鹽培養基中培養時,隨著熒蒽被降解,培養液逐漸變清亮,菌體呈絮狀聚集(圖1d)。
2.2 菌株CE3的分子生物學鑒定
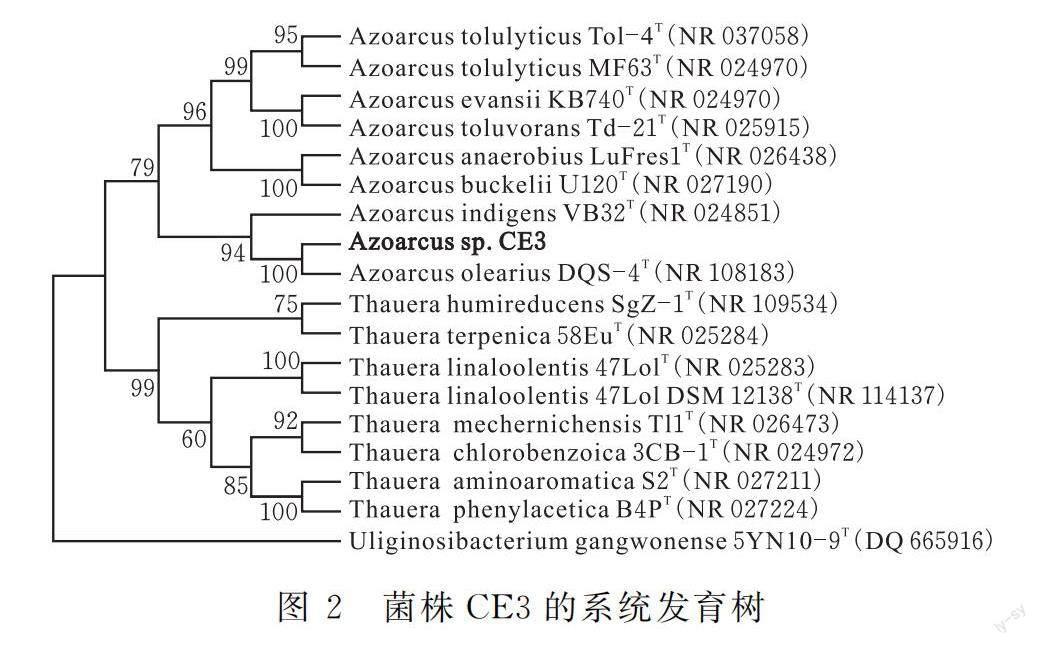
菌株CE3的16S rDNA序列GenBank 登錄號為 MT 498785,Blast序列同源性比對顯示其菌株CE3與Azoarcus(固氮彎曲菌)屬細菌的16S rDNA序列同源性高,核苷酸一致性均達95%以上,其中與Azoarcus evansii DQS-4的16S rDNA序列同源性最高。系統發育分析也表明菌株CE3與彎曲固氮菌屬(Azoarcus)的菌株親緣關系較近(圖2),故初步將菌株CE3鑒定為Azoarcus sp. CE3。
2.3 菌株CE3的多環芳烴利用范圍
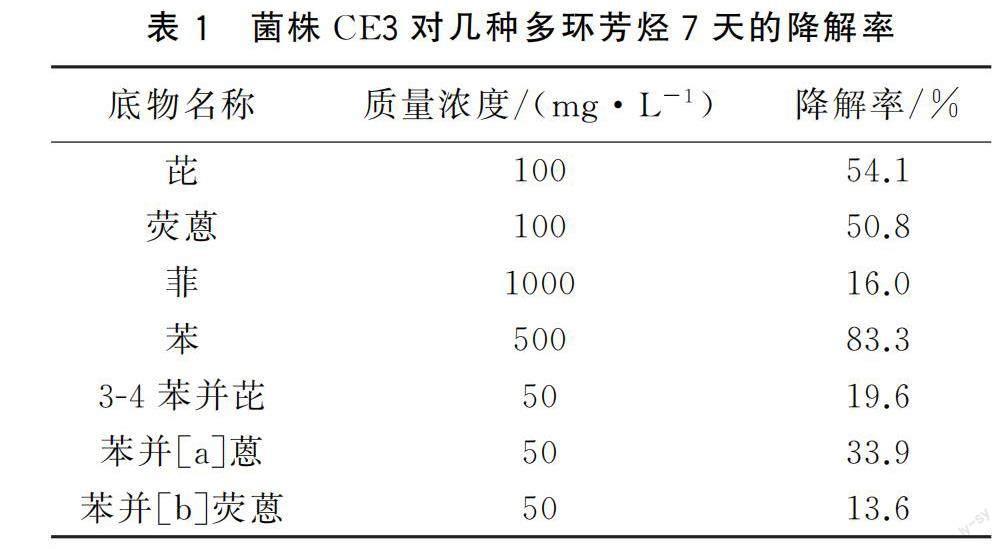
實驗發現,除熒蒽外,菌株CE3能在以苯、芘、菲、3-4苯并芘、苯并[a]蒽、苯并[b]熒蒽為唯一碳源的無機鹽培養基中生長。為此,檢測了菌株CE3對各種高分子量PAHs的降解能力(表1)。結果表明菌株CE3對幾種高分子量PAHs都有一定的降解能力,對熒蒽和芘的降解率超過了50%。故后續對菌株CE3在不同條件下降解熒蒽和芘的能力進行了詳細分析。
2.4 菌株CE3降解特性的研究
2.4.1 培養溫度對菌株CE3降解率的影響 未接種細菌的對照組中熒蒽和芘濃度在不同培養溫度下無明顯差別,說明溫度對熒蒽和芘的自身散逸沒有顯著影響。菌株CE3降解熒蒽和芘的最適溫度均為30℃,溫度提高到35℃時,CE3菌對芘的降解率略有降低,但對熒蒽的降解率則下降了一半(圖3)。菌株CE3在25~45℃條件下都能生長,但溫度超過40℃時CE3菌的生長受到明顯抑制,降解率也明顯降低。因此,后續的實驗都在30℃條件下進行。
2.4.2 不同接菌量對菌株CE3降解率的影響 降解體系中的PAHs含量恒定時,不同的接菌量對降解率也有明顯影響。實驗結果表明,接菌量從2%到8%時,菌株CE3對熒蒽的降解率逐漸提高,最高降解率達到58.9%;增加接種量到10%時,降解率反而降低。芘的降解率變化與熒蒽的不同,當接菌量為6%時,菌株CE3對芘的降解率達到最大值60.2%。其他不同比例接菌量下,芘的降解率無明顯差別(圖4)。
2.4.3 熒蒽或芘初始濃度對菌株CE3降解能力的影響 菌株對PAHs耐受能力的研究是探究其降解特性的重要一環,本研究測定了不同初始培養濃度下菌株CE3對熒蒽和芘的降解效率(圖5)。初始濃度為100 mg/L時,菌株CE3對熒蒽和芘的降解率最高,分別達到50.8%和65.3%,因此,后續實驗中熒蒽或芘的初始濃度均為100 mg/L。在熒蒽和芘濃度較高時,菌株CE3仍具有一定的降解能力,說明菌株CE3能耐受高濃度熒蒽和芘,具有應用于高濃度熒蒽或芘污染治理的潛力。
2.4.4培養基的初始pH對菌株CE3降解率的影響 pH值會影響酶的活性、物質的轉運過程和營養物質的利用[8],從而影響微生物的代謝過程,因此本研究探究了培養基初始pH對菌株CE3降解熒蒽和芘效率的影響(圖6)。培養基初始pH值在5~10之間時,菌株CE3對熒蒽和芘的降解率先隨著pH值的增加而增加,當pH為8時CE3菌對熒蒽的降解率達到最高,隨后pH越高降解率越低;而菌株CE3對芘的降解率在pH 9時達到峰值,當pH為10 或8時,芘的降解率也沒有明顯降低。由此可見,菌株CE3比較適應弱堿性環境,而酸性環境對其生長和降解活動明顯不利。
2.4.5 不同外源營養物質對菌株CE3降解率的影響 微生物需要碳源、氮源、磷源、鉀和鐵等保證其正常代謝和生長[9],不同的共代謝底物也是影響微生物降解PAHs的因素之一[10]。本研究選擇蔗糖(A)、果糖(B)、葡萄糖(C)、麥芽糖(D)、木糖(E)、酵母提取物(F)、蛋白胨(G)作為外源營養物質,探究它們對菌株CE3降解熒蒽和芘能力的影響。結果顯所示,各種營養物質對菌株CE3降解熒蒽和芘的影響不同,酵母提取物使菌株對熒蒽和芘的降解率分別提高了12.3%和5.8%,蛋白胨使菌株對熒蒽和芘的降解率分別提高了5.7%和3.6%;而蔗糖使菌株對熒蒽的降解率提高了11.5%,但抑制了對芘的降解;與之相反,果糖使菌株對芘的降解率提高了13.9%,抑制了對熒蒽的降解(圖7)。因此,為了促進菌株CE3對熒蒽和芘的降解,可在降解體系中添加酵母提取物和蛋白胨,作為共代謝底物促進菌株CE3對熒蒽和芘的降解;如果只需降解熒蒽,可以選取蔗糖作為外源碳源物質促進降解;如果只需降解芘,則可選取果糖作為外源碳源物質促進降解。
3 討論
本文從PAHs污染土壤中以熒蒽為唯一碳源分離到一株PAHs降解菌CE3,通過分子生物學方法鑒定,該菌為Azoarcus sp. CE3。雖然早先有報道Azoarcus屬的菌能夠厭氧降解含單個苯環的芳香烴和單個六元環結構如間二甲苯、二氧己烷等有機物[11-13],還有報道通過宏基因組測序發現一種能降解熒蒽和菲的混合菌中Azoarcus屬細菌的含量高達58.5%[14],但本研究是首次分離到能降解PAHs的Azoarcus屬菌株。CE3菌能以多種PAHs為唯一碳源進行生長,特別是能降解幾種高分子量PAHs,說明CE3菌具有應用于環境PAHs污染治理的潛力。探究各種環境條件對菌株CE3降解熒蒽或芘的影響,發現菌株CE3能在較廣的溫度和pH范圍降解熒蒽或芘;同時,添加酵母提取物、蔗糖和果糖可使CE3菌降解率提高。因此,菌株CE3在PAHs類污染物的修復中具有廣闊的應用前景,本實驗為CE3菌應用于環境PAHs污染治理及修復奠定了良好的基礎。
由于目前僅完成實驗室水平的液體體系降解測試,為實現CE3菌在PAHs污染土壤或水體中的應用,后期需要繼續探究CE3菌在應用于環境治理時是否能保持其高效利用底物的特性,是否會與環境中原有的微生物群落相互作用等問題。
[ 參 考 文 獻 ]
[1] MOORTHY B, CHU C, CARLIN D J. Polycyclic aromatic hydrocarbons: from metabolism to lung cancer[J]. Toxicological Sciences, 2015, 145(01): 5-15.
[2] LABIB S, WILLIAMS A, KUO B, et al. A framework for the use of single-chemical transcriptomics data in predicting the hazards associated with complex mixtures of polycyclic aromatic hydrocarbons[J]. Archives of Toxicology, 2017, 91(7): 2599-2616.
[3] IZZO S A, QUINTANA S, ESPINOSA M, et al. First characterization of PAH-degrading bacteria from Rio de la Plata and high-resolution melting: an encouraging step toward bioremediation[J]. Environmental Technology, 2019, 40(10): 1250-1261.
[4] SHI W, GUO Y, NING G, et al. Remediation of soil polluted with HMW-PAHs by alfalfa or brome in combination with fungi and starch[J]. Journal of Hazardous Materials, 2018, 360: 115-121.
[5] HESHAM A L, KHAN S, TAO Y, et al. Biodegradation of high molecular weight PAHs using isolated yeast mixtures: application of meta-genomic methods for community structure analyses[J]. Environmental Science and Pollution Research, 2012, 19(8): 3568-3578.
[6] 馬妍, 程蘆, 阮子淵,等. 近20年中國表層土壤中多環芳烴時空分布特征及源解析[J]. 環境科學, 2021, 42(03): 1065-1072.
[7] GHOSAL D, GHOSH S, DUTTA T K, et al. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A Review[J]. Frontiers in Microbiology, 2016, 7: 1369.
[8] PATEL V, PATEL J, MADAMWAR D. Biodegradation of phenanthrene in bioaugmented microcosm by consortium ASP developed from coastal sediment of Alang-Sosiya ship breaking yard[J]. Marine Pollution Bulletin, 2013, 74(01): 199-207.
[9] LEE K, PARK J W, AHN I S. Effect of additional carbon source on naphthalene biodegradation by Pseudomonas putida G7[J]. Journal of Hazardous Materials, 2003, 105(01-03): 157-167.
[10]VAIDYA S, DEVPURA N, JAIN K, et al. Degradation of chrysene by enriched bacterial consortium[J]. Frontiers in Microbiology, 2018, 9: 1333.
[11]FERNANDEZ H, PRANDONI N, FERNANDEZ-PASCUAL M, et al. Azoarcus sp. CIB, an anaerobic biodegrader of aromatic compounds shows an endophytic lifestyle[J]. PLoS One, 2014, 9(10): e110771.
[12]DENG D, PHAM D N, LI F, et al. Discovery of an inducible toluene monooxygenase that cooxidizes 1,4-dioxane and 1,1-dichloroethylene in propanotrophic Azoarcus sp. strain DD4[J]. Applied and Environmental Microbiology, 2020, 86(17): e01163-20.
[13]BLAZQUEZ B, CARMONA M, DIAZ E. Transcriptional regulation of the peripheral pathway for the anaerobic catabolism of toluene and m-xylene in Azoarcus sp. CIB[J]. Frontiers in Microbiology, 2018, 9: 506.
[14]PATEL A B, SINGH S, PATEL A, et al. Synergistic biodegradation of phenanthrene and fluoranthene by mixed bacterial cultures[J]. Bioresource Technology, 2019, 284: 115-120.
Degradation Characteristics of Polycyclic Aromatic
Hydrocarbons by Azoarcus sp. CE3
ZHANG Honghong1, YE Chen1, ZHANG Xiaoyun2, MENG Tian1,WANG Liming2, HUANG Yuping1
(1 College of Life Sciences, Wuhan University, Wuhan 430072, China;
2 Hubei Environmental Remediation & Governance
Techonlogical Research Co., ltd.,Huangshi 435000,China)
Abstract:Microbial remediation has excellent characteristics in the control of polycyclic aromatic hydrocarbons (PAHs) pollution. In order to obtain microorganisms that can efficiently degrade PAHs, fluoranthene was used as the only carbon source to screen microorganisms. And then a high-efficiency degrading bacterium named CE3 was isolated. Strain CE3 was identified to belong to Azoarcus genus based on its 16S rDNA sequence analysis. To our knowledge, this is the first report that the strain of the Azoarcus genus has the ability to degrade high molecular weight PAHs. In addition to fluoranthene, strain CE3 also could degrade benzene, phenanthrene, pyrene, fluoranthene, benzo [a]anthracene, benzo [3,4]pyrene and benzo [b]fluoranthene. The degradation rates of fluoranthene and pyrene are more than 50% and higher than the degradation rates of other PAHs. Then, the degradation rates of fluoranthene and pyrene by strain CE3 under various conditions were analyzed in detail by HPLC method. The results showed that Azoarcus sp. CE3 could degrade fluoranthene or pyrene in a wide range of temperature and pH; moreover, the addition of yeast extract, sucrose and fructose to the medium could improve its degradation ability.
Keywords:polycyclic aromatic hydrocarbons (PAHs); fluoranthene; pyrene; degradation; Azoarcus
[責任編校:張 眾]
