基于生物信息學(xué)預(yù)測(cè)分析C/EBPα靶基因及其在急性髓系白血病中的潛在機(jī)制
2023-12-07 06:33:42章瑞琪
麗水學(xué)院學(xué)報(bào) 2023年5期
章瑞琪,郭 虹
(麗水學(xué)院醫(yī)學(xué)院,浙江 麗水 323000)
急性髓系白血病(acute myeloid leukemia,AML)是一種骨髓和外周血髓系細(xì)胞發(fā)育受阻的惡性腫瘤,主要表現(xiàn)為髓系原始細(xì)胞增殖不受控制,骨髓分化受抑制,從而抑制正常血細(xì)胞的發(fā)育。該病占白血病總發(fā)病數(shù)的50%以上,其中在AML 亞型之中M2與M5的發(fā)病率較高[1]。而AML的發(fā)生、發(fā)展、治療和預(yù)后與轉(zhuǎn)錄因子CCAAT 增強(qiáng)子結(jié)合蛋白α(CCAAT/enhancer binding protein alpha,C/EBPα)的異常表達(dá)密切相關(guān)。在中國AML成年患者中,約有11.7%患者的CEBPA基因突變陽性[2],這干擾了CEBPA基因的正常表達(dá),阻礙了其功能的發(fā)揮。而正常表達(dá)的轉(zhuǎn)錄因子C/EBPα因含有堿性亮氨酸拉鏈結(jié)構(gòu)域,可以通過形成二聚體、與目標(biāo)基因的啟動(dòng)子區(qū)結(jié)合、調(diào)控轉(zhuǎn)錄激活等手段發(fā)揮其對(duì)維持血液系統(tǒng)穩(wěn)定的功能:維持造血干細(xì)胞的自我更新、支持粒細(xì)胞的分化,并參與調(diào)節(jié)細(xì)胞周期與新陳代謝過程。
為進(jìn)一步闡釋C/EBPα 的異常表達(dá)與AML 間的作用機(jī)制,筆者利用生物信息學(xué)技術(shù)探究C/EBPα的靶基因,并通過分析靶基因的功能與信號(hào)轉(zhuǎn)導(dǎo)通路,進(jìn)一步了解C/EBPα 的轉(zhuǎn)錄調(diào)控功能,且重點(diǎn)揭示靶基因信號(hào)轉(zhuǎn)導(dǎo)通路(JAK-STAT信號(hào)通路、MAPK通路、PI3K-Akt通路),總結(jié)它們對(duì)維持正常血液系統(tǒng)的功能和在AML 發(fā)生、發(fā)展及治療過程中的作用與影響,揭開C/EBPα通過靶基因信號(hào)轉(zhuǎn)導(dǎo)通路在AML 中的潛在機(jī)制,并進(jìn)而為AML臨床治療與預(yù)后提供依據(jù)。
1 材料與方法
1.1 數(shù)據(jù)獲取
通過GeneCards 數(shù)據(jù)庫(http://www.genecards.org)檢索得到107個(gè)與AML相關(guān)的基因,將其作為候選預(yù)測(cè)靶基因。在基因表達(dá)數(shù)據(jù)庫(https://www.ncbi.nlm.nih.gov/geo/)中檢索得到包含敲除Cebpa基因的實(shí)驗(yàn)組與對(duì)照組的GSE146288、GSE153622、GSE71687數(shù)據(jù)集。
1.2 篩選差異表達(dá)基因
運(yùn)用GEO 數(shù)據(jù)庫提供的GEO2R 分析工具對(duì)上述107 個(gè)候選預(yù)測(cè)靶基因的表達(dá)量進(jìn)行綜合分析,篩選Cebpa敲除前后的差異表達(dá)基因,得出其在Cebpa基因敲除前后的比較結(jié)果。
1.3 差異基因的GO功能與KEGG富集分析
為進(jìn)一步解釋差異表達(dá)基因在AML發(fā)病過程中發(fā)揮的作用,對(duì)C/EBPα的42個(gè)差異表達(dá)靶基因進(jìn)行分析。借助DAVID 數(shù)據(jù)庫綜合分析工具(http://david.ncifcrf.gov)進(jìn)行基因本體論(gene ontology,GO)富集分析、京都基因與基因組百科全書(kyoto encyclopedia of genes and genomes,KEGG)富集分析,篩選顯著差異的數(shù)據(jù)(P<0.05),探索差異基因潛在影響的功能與通路。
1.4 C/EBPα 與信號(hào)通路中差異表達(dá)基因的蛋白質(zhì)互作網(wǎng)絡(luò)
利用STRING 數(shù)據(jù)庫(https://cn.string-db.org/)分析調(diào)控網(wǎng)絡(luò)中的蛋白質(zhì)-蛋白質(zhì)相互作用(protein-protein interaction,PPI),對(duì)KEGG 富集分析結(jié)果中的JAK-STAT信號(hào)通路、MAPK信號(hào)通路與PI3K-Akt 信號(hào)通路中所包含的基因進(jìn)行分析,構(gòu)建其與C/EBPα之間的蛋白質(zhì)互作網(wǎng)絡(luò)。
2 結(jié)果
2.1 篩選差異表達(dá)基因結(jié)果
運(yùn)用GEO2R分析工具對(duì)上述107個(gè)候選預(yù)測(cè)靶基因的表達(dá)量進(jìn)行綜合分析,得出其在Cebpa基因敲除前后的比較結(jié)果(見表1),將表達(dá)量隨Cebpa基因敲除而下降的基因作為預(yù)測(cè)靶基因,共42個(gè),并對(duì)這些差異表達(dá)基因開展深入研究。

表1 Cebpa基因敲除后AML相關(guān)基因表達(dá)
2.2 差異基因的GO功能富集
借助DAVID 綜合分析工具對(duì)C/EBPα的42個(gè)差異表達(dá)靶基因進(jìn)行GO富集分析,篩選顯著差異的數(shù)據(jù)(P<0.05),得到GO 富集分析結(jié)果:主要富集于26個(gè)基因功能,按生物學(xué)過程、細(xì)胞組分和分子功能3類基因功能分類,分類結(jié)果如圖1所示。
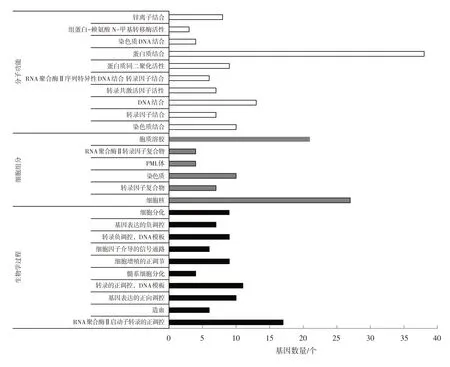
圖1 C/EBPα預(yù)測(cè)靶基因的GO功能富集分析
由觀察分析結(jié)果可知:在生物學(xué)過程分類中,差異基因主要富集于RNA聚合酶Ⅱ啟動(dòng)子轉(zhuǎn)錄的正調(diào)控、造血、基因表達(dá)的正向調(diào)控等。在細(xì)胞組分分類中,差異基因主要富集于細(xì)胞核、轉(zhuǎn)錄因子復(fù)合物、染色質(zhì)等。在分子功能分類中,差異基因主要富集于蛋白質(zhì)結(jié)合、染色質(zhì)結(jié)合、轉(zhuǎn)錄因子結(jié)合、DNA結(jié)合等。
2.3 差異基因的KEGG通路分析
在GO功能富集分析的基礎(chǔ)上,利用DAVID綜合分析工具進(jìn)行KEGG富集分析,篩選差異顯著的數(shù)據(jù)(P<0.05),得到KEGG 富集分析結(jié)果,結(jié)果如圖2所示:差異基因主要富集于癌癥的通路、急性髓性白血病、癌癥中的轉(zhuǎn)錄失調(diào)、癌癥中的微小RNA、慢性粒細(xì)胞白血病、PI3K-Akt 信號(hào)通路、JAKSTAT信號(hào)通路、乙型肝炎、造血細(xì)胞譜系、MAPK信號(hào)通路。差異基因在癌癥的通路中富集最多,共有15 個(gè)基因富集于此通路。借助STRING 在線數(shù)據(jù)庫,繪制PI3K-Akt 信號(hào)通路、JAK-STAT信號(hào)通路與MAPK信號(hào)通路網(wǎng)絡(luò)圖,結(jié)果如圖3所示。
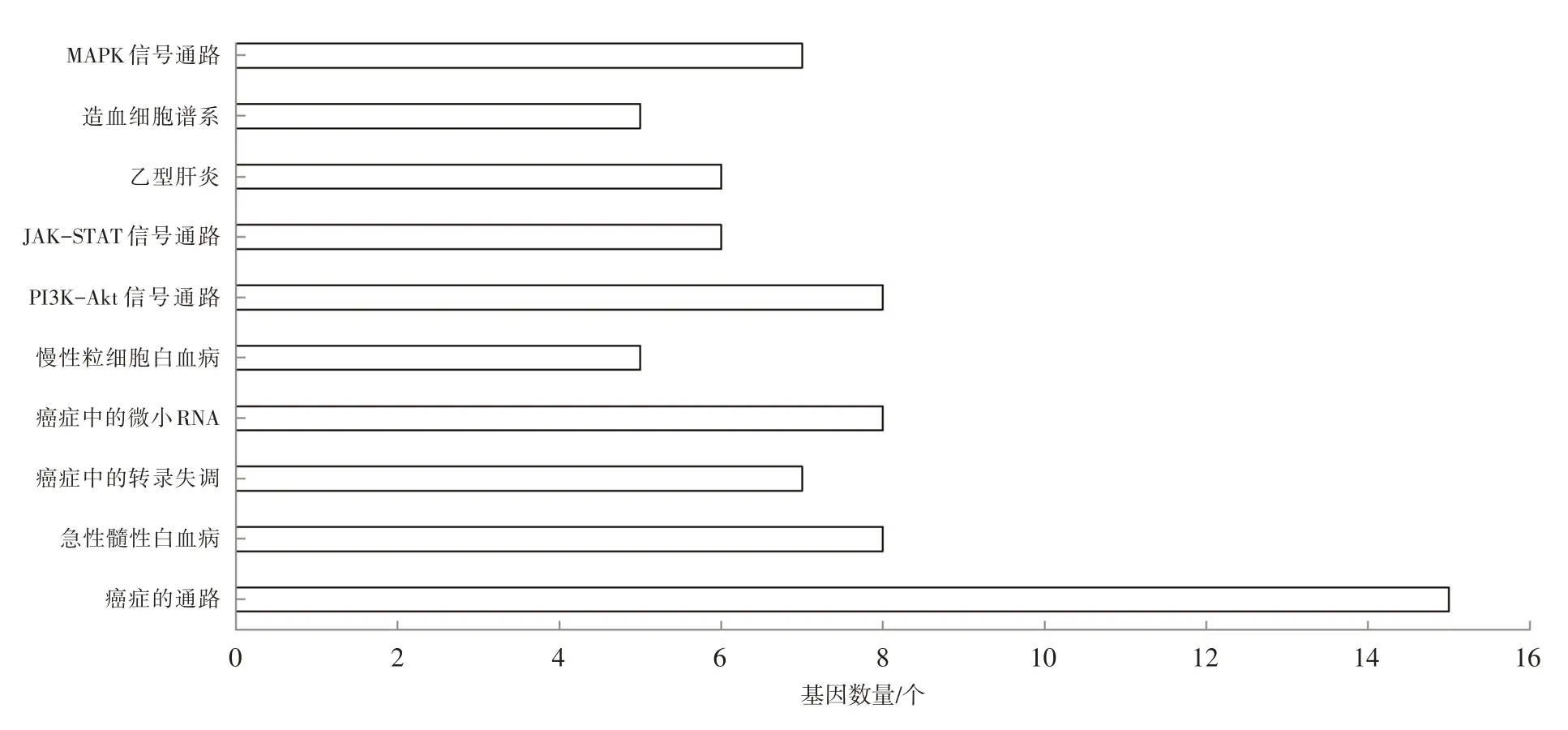
圖2 C/EBPα預(yù)測(cè)靶基因的KEGG信號(hào)轉(zhuǎn)導(dǎo)富集分析

圖3 KEGG信號(hào)通路的蛋白質(zhì)互作網(wǎng)絡(luò)
2.4 C/EBPα與信號(hào)通路的蛋白質(zhì)互作網(wǎng)絡(luò)
分別將JAK-STAT信號(hào)通路、MAPK信號(hào)通路與PI3K-Akt 信號(hào)通路所包含的蛋白基因?qū)隨TRING在線數(shù)據(jù)庫,獲取上述通路中所包含差異基因與C/EBPα的相互作用網(wǎng)絡(luò),結(jié)果如圖4所示:C/EBPα與JAK-STAT信號(hào)通路所包含的差異基因間共有6 條相互作用關(guān)系;C/EBPα 與MAPK 信號(hào)通路所包含的差異基因間共有6條相互作用關(guān)系;C/EBPα 與PI3K-Akt 信號(hào)通路所包含的差異基因間共有5條相互作用關(guān)系。

圖4 C/EBPα與MAPK、JAK-STAT及PI3K-Akt信號(hào)通路的蛋白質(zhì)互作網(wǎng)絡(luò)
3 討論
轉(zhuǎn)錄因子C/EBPα 的異常表達(dá)對(duì)AML 的發(fā)病及預(yù)后產(chǎn)生重要影響,研究表明C/EBPα能夠調(diào)節(jié)造血干細(xì)胞的細(xì)胞周期,促進(jìn)中性粒細(xì)胞的分化,對(duì)骨髓發(fā)育起著重要作用。大部分AML患者的C/EBPα 表達(dá)量下降,但C/EBPα 不同的突變形式影響著AML 患者的預(yù)后,C/EBPα 單突變的AML 患者往往預(yù)后較差[3],而C/EBPα雙突變的AML患者的死亡率較低,往往預(yù)后較好[4]。這說明C/EBPα的不同表達(dá)會(huì)對(duì)AML的發(fā)病與預(yù)后產(chǎn)生不同的影響,借助生物信息學(xué)技術(shù),研究C/EBPα 靶基因功能及其信號(hào)通路,有利于進(jìn)一步揭示AML 的發(fā)病機(jī)理,同時(shí)為臨床開發(fā)針對(duì)C/EBPα靶點(diǎn)的免疫治療藥物提供理論支持。
本研究主要運(yùn)用生物信息學(xué)技術(shù),在GEO 數(shù)據(jù)庫中篩選敲除Cebpa基因的實(shí)驗(yàn)組及正常造血干細(xì)胞間的差異表達(dá)基因,發(fā)現(xiàn)差異表達(dá)基因共86 個(gè),其中共有42 個(gè)基因隨Cebpa基因的敲除而表達(dá)下降,將其作為預(yù)測(cè)靶基因進(jìn)行了GO 功能分析和KEGG 通路富集分析,結(jié)果發(fā)現(xiàn)這些基因在RNA聚合酶Ⅱ啟動(dòng)子轉(zhuǎn)錄的正調(diào)控、造血、基因表達(dá)的正向調(diào)控等功能上富集,并且很可能通過JAK-STAT信號(hào)通路、MAPK信號(hào)通路及PI3K-Akt信號(hào)通路發(fā)揮作用。不僅如此,我們還證明了C/EBPα 與JAK-STAT 信號(hào)通路、MAPK 信號(hào)通路、PI3K-Akt信號(hào)通路中所包含的差異表達(dá)基因有互相作用,提示C/EBPα 功能的發(fā)揮與上述3 條通路的信號(hào)傳導(dǎo)具有密切聯(lián)系,推測(cè)轉(zhuǎn)錄因子C/EBPα可能通過JAK-STAT 信號(hào)通路、MAPK 信號(hào)通路、PI3K-Akt信號(hào)通路的傳導(dǎo)對(duì)急性髓系白血病的發(fā)病與預(yù)后產(chǎn)生一定影響。
已有研究已論證:JAK-STAT信號(hào)通路在非典型C/EBPα雙突變的AML患者中會(huì)被異常激活,破壞其原有抗腫瘤作用,使得穩(wěn)態(tài)失衡,進(jìn)而加劇AML患者的病情發(fā)展[5];MAPK信號(hào)通路主要包括MEK1/2-ERK1/2 通路、JNKs 通路、p38 通路及MEK5-ERK5-MEF2C 通路4 類信號(hào)通路,在調(diào)控細(xì)胞增殖、凋亡及細(xì)胞周期方面發(fā)揮著重要作用[6-8],其中MEK1/2-ERK1/2 通路活性的增強(qiáng),會(huì)下調(diào)C/EBPα 的表達(dá)量,從而抑制粒細(xì)胞分化,進(jìn)而誘導(dǎo)AML 的發(fā)生[9-10],而MEK5-ERK5-MEF2C通路也能夠通過調(diào)控轉(zhuǎn)錄因子C/EBPα 與C/EBPβ來引導(dǎo)髓系分化[11];PI3K-Akt 信號(hào)通路能夠控制C/EBPα 的磷酸化,以此介導(dǎo)骨髓細(xì)胞正常發(fā)育[12],而該通路的過度激活往往會(huì)導(dǎo)致AML 細(xì)胞不受控制的增殖[13]。這些論據(jù)均說明C/EBPα 的異常表達(dá)對(duì)AML患者的影響與JAK-STAT信號(hào)通路、MAPK 信號(hào)通路、PI3K-Akt 信號(hào)通路的傳導(dǎo)密不可分。
綜上所述,本研究篩選出與C/EBPα表達(dá)相關(guān)的差異基因,并分析所述基因的功能及其相關(guān)信號(hào)通路,研究結(jié)果有助于探索尋求臨床治療AML的免疫靶點(diǎn),為揭示AML 發(fā)生發(fā)展過程提供理論基礎(chǔ)。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
小天使·一年級(jí)語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
電子制作(2018年11期)2018-08-04 03:25:42
財(cái)經(jīng)(2017年2期)2017-03-10 14:35:35
財(cái)經(jīng)(2016年15期)2016-06-03 07:38:02
財(cái)經(jīng)(2016年3期)2016-03-07 07:44:46
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
