高產四乙基愈創木酚耐鹽乳酸菌的篩選及其發酵特性研究
2023-12-08 03:34:50李祥弟徐婷劉占張麗杰王棟
食品與發酵工業 2023年22期
關鍵詞:檢測
李祥弟,徐婷,劉占,張麗杰,王棟*
1(釀造微生物與應用酶學研究室(江南大學),江蘇 無錫,214122) 2(工業生物技術教育部重點實驗室(江南大學), 江蘇 無錫,214122) 3(廣東美味鮮調味食品有限公司,廣東 中山,528400)
醬油是一種在東亞地區不可缺少的發酵調味品,因其濃郁的鮮味和獨特的香氣在世界范圍內也廣受歡迎。醬油的制作過程大致可分為4個部分,即原料蒸煮、制曲、醬醪發酵以及滅菌罐裝[1]。成熟醬醪的理化性質和風味特征與醬油品質密切相關,因此,醬醪發酵是醬油生產的關鍵階段之一。在醬醪發酵過程中,耐鹽乳酸菌起到了至關重要的作用[2]。在醬醪發酵前期,耐鹽乳酸菌會代謝產生有機酸,降低發酵體系pH以有利于產香酵母菌的增殖。同時,耐鹽乳酸菌可以起到提升風味[3]、抑制有害微生物生長[4]、抑制有毒物質形成[5-6]等作用。
嗜鹽四聯球菌(Tetragenococcushalophilus)屬于四聯球菌屬(Tetragenococcus),表現為兼性厭氧發酵模式,可以在高鹽(20% NaCl)或高糖的環境中生長,是我國高鹽稀態醬油發酵過程(自發發酵)中的主體耐鹽乳酸菌。有報道顯示,嗜鹽四聯球菌可以提高醬油中風味物質含量,是日本醬油、魚醬等純菌發酵過程中的初始發酵菌劑(starter)。不僅如此,嗜鹽四聯球菌與醬油主體產香菌魯氏結合酵母(Zygosaccharomycesrouxi)具有種間相互作用,前者可以有效提高魯氏結合酵母的耐鹽性[7]。因此兩菌種共培養可以進一步提高醬油中風味物質含量。然而,對嗜鹽四聯球菌的泛基因組研究表明,與菌種發酵性能對應,該種不同株的基因組呈現明顯的菌株水平特異性,即不同菌種在菌種安全性、產香性能、風味增強等方面具有明顯差異。因此,篩選具有優良特性的嗜鹽四聯球菌以用于我國高鹽稀態醬油醬醪發酵,對進一步提高醬油品質具有重要潛在價值[8-10]。
四乙基愈創木酚(4-ethylguaiacol, 4-EG)是醬油中一種關鍵的風味物質,呈煙熏及辛香味,具有緩和咸味的作用。四乙基愈創木酚風味閾值極低,樣品間0.5 mg/L的差異就能被人體感官識別,因此其含量與醬油品質密切相關。已有研究表明,微生物合成四乙基愈創木酚主要以阿魏酸為前體,通過阿魏酸脫羧酶,轉化為四乙烯基愈創木酚,后經四乙烯基愈創木酚還原酶轉化為四乙基愈創木酚。且已有研究表明強化四乙基愈創木酚產生菌可以提高醬油中四乙基愈創木酚的含量[11]。
本文嘗試從醬油醬醪中分離篩選出高產四乙基愈創木酚的耐鹽乳酸菌,對其進行種屬鑒定,并測試該菌種耐酸耐鹽性能及菌種安全性能。以該菌種為初始發酵菌劑用于模擬醬油發酵,與不加發酵劑的醬油發酵樣本相比,四乙基愈創木酚含量提高3.34倍。
1 材料與方法
1.1 材料與儀器
1.1.1 樣品
菌株篩選樣品,國內某醬油廠醬醪;制曲用米曲霉滬釀3.042,江南大學釀造微生物學與應用酶學研究室菌種庫。
1.1.2 試劑
面粉、麩皮、黃豆,市售;阿魏酸、四乙基愈創木酚、天冬氨酸、丙氨酸、組氨酸,百靈威科技公司;組胺、乳酸、乙酸、葡萄糖、甲酸,美國sigma公司;氯化鈉、三氯乙酸、氫氧化鈉、甲醛、酚酞、鄰苯二甲酸氫鉀、無水乙醇、乙腈、甲醇、硫酸,國藥集團化學試劑有限公司;細菌DNA提取試劑盒,南京諾唯贊生物科技股份有限公司。
1.1.3 培養基
改良MRS液體培養基,青島海博生物科技公司,按說明使用,并添加8 g/100 mL NaCl,自然pH。
改良MRS固體培養基:同改良MRS液體培養基配制方法,再加入2%(質量分數)的瓊脂粉。
發酵培養基(g/100 mL):MRS培養基53,NaCl 8,阿魏酸0.1,自然pH。
種曲培養基:按照m(麩皮)∶m(面粉)=7∶3的比例在250 mL三角瓶中加入20 g過10目篩的干料,并加入干料量70%(質量分數)的水,拌勻。
以上培養基均在121 ℃滅菌20 min。
1.1.4 儀器與設備
電子分析天平、pH計,Mettler Toledo公司;PCR儀,美國Bio-Rad公司;高壓濕熱自動蒸汽滅菌鍋,Tomy公司;超凈工作臺,蘇州凈化設備有限公司;培養箱,上海森信實驗儀器有限公司;高速離心機,Eppendorf公司;CytationTM3多功能酶標儀,BioTek公司;BX51光學顯微鏡,Olympus公司;超高效液相色譜儀,美國Waters公司;氣相質譜儀、高效液相色譜,安捷倫科技公司。
1.2 實驗方法
1.2.1 嗜鹽四聯球菌的獲取
無菌條件下取適量醬醪,溶于無菌水中,振蕩均勻,使用無菌生理鹽水進行稀釋,取不同稀釋梯度涂布于改良MRS固體培養基中,30 ℃厭氧靜置培養7 d,觀察菌株形態。挑取單菌落于改良MRS液體培養基中,30 ℃靜置培養48 h,甘油保藏于-80 ℃冰箱。
1.2.2 菌株的鑒定
a)形態觀察:經過發酵實驗篩選的目標菌株,將其菌懸液梯度稀釋后涂布于MRS平板培養基,對其進行細胞觀察及菌落形態觀察。
b)分子生物學鑒定:使用細菌基因組提取試劑盒對菌株基因組進行提取,使用通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′)對其16S rRNA進行PCR擴增[12],PCR產物送至上海生工生物公司進行測序,測序結果使用NCBI上Blast工具進行比對,鑒定菌株。下載四聯球菌同屬其他模式菌株的16S rRNA序列,采用MEGA 7.0軟件中的鄰接(neighbor joining, NJ)法構建系統發育樹。
1.2.3 高產四乙基愈創木酚菌株的篩選
將初篩所得嗜鹽四聯球菌活化為種子液,按3%比例接種至發酵培養基,30 ℃厭氧靜置培養10 d,結束后采用0.22 μm水系濾膜過濾發酵上清液,超高效液相色譜(ultra-performance liquid chromatography, UPLC)檢測發酵液中四乙基愈創木酚含量,篩選出高產菌株。
UPLC條件:色譜柱ACQUITY UPLC?HSS T3(1.8 μm×2.1 mm×150 mm);流動相:A(0.1%甲酸水),B(甲醇);洗脫程序:0~8 min,100%~48% A;8~10 min,48% A;10~15 min,48%~30% A;15~18 min,30%~100% A;流速為0.3 mL/min;檢測波長280 nm;柱溫30 ℃;進樣量10 μL[13]。
制定標準曲線:取標準品阿魏酸、四乙基愈創木酚,溶于甲醇配制質量濃度為100 mg/L的混合標準溶液,依次梯度稀釋為2.5、12.5、25、50、100 mg/L。以標準溶液濃度為橫坐標,峰面積為縱坐標制定標準曲線,建立線性方程[14]。
1.2.4 菌株生理特征
a)菌株主體代謝產物檢測:將菌株活化為種子液,按3%接種量接種于改良MRS培養基中,30 ℃厭氧發酵10 d,每隔2 d取樣,結束后采用0.22 μm水系濾膜過濾發酵上清液,HPLC檢測發酵液中主要代謝產物含量,方法參考文獻[15]。
b)氨基酸檢測:使用含有0.3%(質量分數)天冬氨酸的MRS培養基,按3%接種量接種,30 ℃培養7 d,取樣使用三氯乙酸處理后采用0.22 μm水系濾膜過濾發酵上清液,采用HPLC在線衍生進行檢測。
c)組胺檢測:使用含有0.1%(質量分數)組氨酸的MRS培養基,按3%接種量接種,30 ℃培養7 d,取樣使用三氯乙酸處理后采用0.22 μm水系濾膜過濾發酵上清液,衍生后使用UPLC進行檢測。
d)菌株生長曲線測定:使用含6%(質量分數)NaCl的MRS培養基,調節pH值到6.0,按3%接種量接入種子液進行培養,培養時間為72 h,每隔8 h取樣,檢測OD600值。
e)菌株在不同pH及鹽度下生長情況:將菌株活化為種子液,按3%接種量接種到含6%、10%、15%、20%(質量分數)NaCl和pH 3.0、4.0、5.0、6.0的MRS培養基中,30 ℃厭氧靜置培養72 h,檢測發酵結束時OD600值。
1.2.5 高鹽稀態醬油發酵工藝
a)種曲制備:將平板培養菌種的孢子用接種針接入種曲培養基,搖勻后30 ℃培養4 d,每日攪拌使曲料松散[16]。
b)大曲制備:大豆用自來水沖洗3次除雜,1.5倍溫水浸泡8 h,瀝干水分。121 ℃滅菌15 min,冷卻至室溫。將大豆、面粉和種曲進行混合,置于恒溫恒濕條件下培養,及時翻曲,制曲48 h,待曲料表面布滿黃色菌絲體,即可出曲。
c)發酵工藝:將大曲與20% NaCl的鹽水按質量比1∶2的比例混合。將混合好的曲料加入含0.1%阿魏酸的500 mL瓶子中,將嗜鹽四聯球菌活化為種子液,離心收集菌體,使用生理鹽水懸浮,按107CFU/g 接種,30 ℃發酵40 d,每隔10 d取樣。
1.2.6 理化指標檢測
pH使用pH計檢測。總酸和氨基酸態氮檢測參照GB 5009.235—2016《食品安全國家標準 食品中氨基酸態氮的測定》方法。游離氨基酸測定具體操作參考已有研究[17],使用氨基酸分析儀進行檢測。揮發性化合物采用固相微萃取聯合氣質聯用技術(phase microextraction-mass spectrometry coupled with gas chromatography-mass spectrometry, SPME-GC-MS)進行測定,詳細處理辦法參照鄭鵬飛等[18]的方法。
2 結果與分析
2.1 高產四乙基愈創木酚菌株的篩選
耐鹽乳酸菌可以在含5%~25%(質量分數)NaCl的厭氧或微耗氧環境生長,最適生長溫度為30 ℃[19],通常呈現白色、表面光滑的圓形菌落形態,為盡可能提高所選耐鹽乳酸菌的菌株多樣性,本研究以我國華東、華南、華中、華北四區域中的10家高鹽發酵調味品公司的高鹽發酵樣品(包括醬醪、魚醬醪等)為篩選源,在含有10%(質量分數)NaCl的MRS培養基,置于30 ℃的厭氧環境中進行耐鹽乳酸菌的篩選。
對上述高鹽發酵樣品進行梯度稀釋涂布平板,分別靜置培養3、5、7 d后,通過初步的形態觀察,總共隨機挑選出350株具有乳酸菌形態特征的菌落。生長性能良好是工業菌種的前提,本研究首先根據培養液菌株生長情況,從中挑選出90株長勢較好的菌株,并進行進一步純化培養。將純化菌種轉接至含0.1%(質量分數)阿魏酸的發酵培養基培養8 d,使用UPLC檢測各發酵液中的四乙基愈創木酚,如圖1所示,菌株LBM 20027展現出突出的四乙基愈創木酚合成能力,含量達到26.8 mg/L。與之相比,嗜鹽四聯球菌模式菌株DSM 20339的產量僅為4.2 mg/L,由此可見耐鹽乳酸菌LBM 20027具有較優的四乙基愈創木酚合成能力。因此使用菌株LBM 20027進行后續研究。
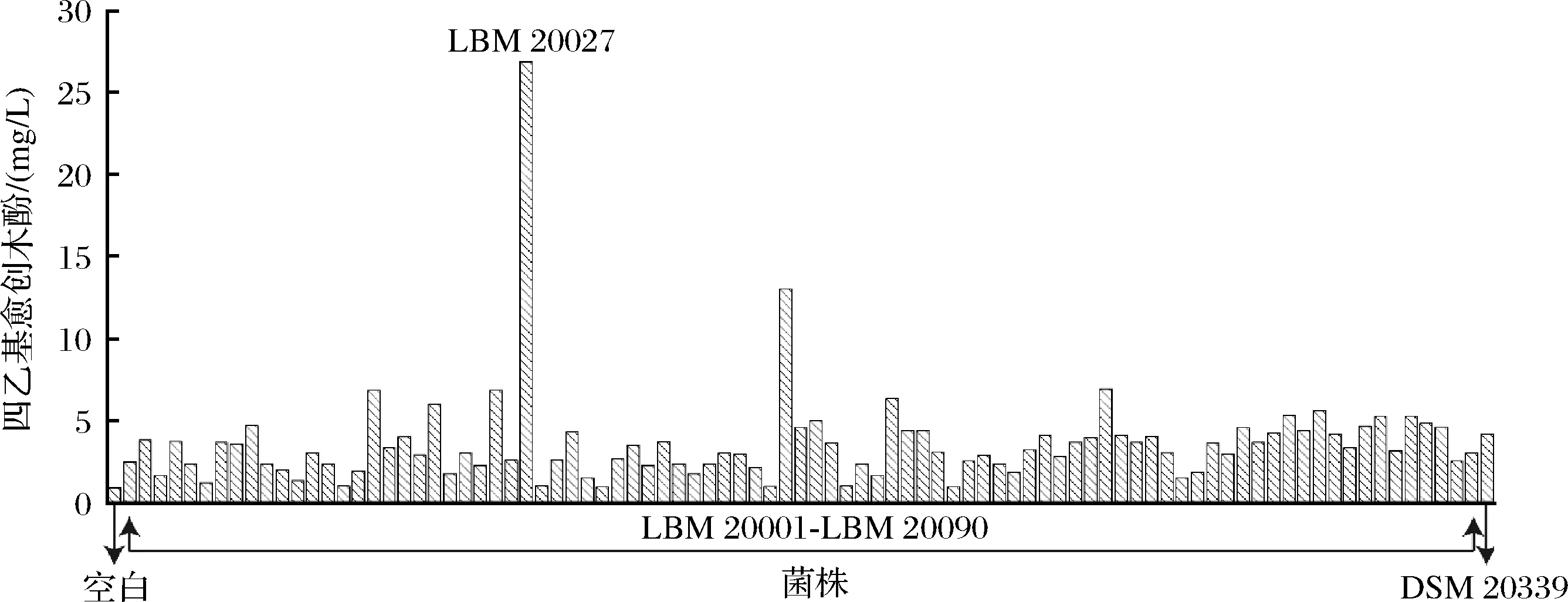
圖1 不同乳酸菌株發酵液中四乙基愈創木酚含量Fig.1 4-Ethylguaiacol concentration in fermentation broth of different lactic acid bacteria strains
2.2 菌株LBM 20027的鑒定
2.2.1 形態學鑒定
如圖2-a所示,菌株LBM 20027菌落形態呈現為乳白色,觀察其邊緣比較規則表面平滑,顯示為圓形菌落。在光學顯微鏡視野下,菌株LBM 20027多呈四聯球型,也有細胞成簇聚集,排列方式多種多樣(圖2-b)。在高精度的掃描電子顯微鏡視野下,高產四乙基愈創木酚的菌株LBM 20027呈典型的成對或四聯球形態(圖2-c)。

a-菌落形態;b-顯微鏡下形態;c-電鏡下形態;d-基于16S rDNA的系統發育樹圖2 菌株LBM 20027形態特征及系統發育樹Fig.2 Morphological characteristics and phylogenetic tree of strain LBM 20027
2.2.2 分子生物學鑒定
使用PCR擴增16S rDNA序列,對高產四乙基愈創木酚菌株LBM 20027進行分子生物學鑒定。測序后獲得基因序列,將其于Genebank數據庫進行 BLAST 比對,比對結果表明該菌株與四聯球菌屬(Tetragenococcus)的幾個菌種具有較高的同源性,可認定其為四聯球菌屬。通過多序列同源性分析,使用MEGA X 7.0的NJ法構建菌株間系統發育樹如圖2-d所示,結果表明菌株LBM 20027與嗜鹽四聯球菌(Tetragenococcushalophilus)的親緣關系最近。根據分子生物學鑒定菌株LBM 20027屬于嗜鹽四聯球菌。而嗜鹽四聯球菌作為醬油發酵中的主體乳酸菌,LBM 20027有潛力更好的在醬油發酵環境下生長,同時其高產四乙基愈創木酚的能力可能有助于醬油風味的提升。
2.3 菌株的發酵性能
菌株的發酵性能,包括生長速率、耐酸耐鹽性能等,是評判該菌株能否在食品發酵過程中成功定植并發揮風味增強作用的關鍵因素之一。另外,對于食品發酵行業來說,菌種安全性是其能否用于食品發酵的重要前提。嗜鹽四聯球菌已在日本醬油生產中有半個世紀的應用歷史,暗示該菌種可能具有食品安全的特性。然而,嗜鹽四聯球菌并非在我國《可用于食品的菌種名單》中,因此,本研究對篩選得到的嗜鹽四聯球菌LBM 20027進行了必要的菌種安全及發酵性能測試。
2.3.1 菌株生長曲線
由圖3-a可知,菌株LBM 20027在培養初期(0~40 h)生長較慢,為生長遲緩期;而在40~48 h生長速率明顯增加,此時為對數增長期;48 h后生長速率逐漸變慢,到56 h進入穩定期,此后OD600值再無明顯變化。

a-生長曲線;b-基礎代謝;c-丙氨酸;d-組胺;e-耐酸性;d-耐鹽性圖3 菌株LBM 20027的生長特性Fig.3 Growth characteristics of strain LBM 20027
2.3.2 菌株主體代謝產物
通過HPLC檢測菌株LBM 20027發酵過程中葡萄糖消耗以及主體代謝產物生成情況,結果如圖3-b所示。可以看出,菌株LBM 20027主要代謝葡萄糖,產生乳酸和乙酸,表現為兼性乳酸發酵形式。在發酵前4 d,葡萄糖被大量消耗,同時迅速生成乳酸和乙酸,第4天后,葡萄糖消耗速率變慢,乳酸和乙酸的生成也變慢,趨于平穩,發酵第10天,生成3.35 g/L乳酸和1.14 g/L乙酸。
2.3.3 菌株氨基酸轉化
嗜鹽四聯球菌可能具有將天冬氨酸轉化為甜味氨基酸——丙氨酸的能力,而丙氨酸呈現甜味,能豐富醬油的口感。因此,是否能將天冬氨酸有效轉化為丙氨酸是嗜鹽四聯球菌性能的重要指標之一。本研究對高產四乙基愈創木酚的菌種LBM 20027和模式菌株DSM 20339的天冬氨酸轉化能力進行檢測。結果如圖3-c所示,在含有0.3%天冬氨酸的MRS培養基培養7 d后,LBM 20027生成了28 mg/L的丙氨酸,而模式菌株DSM 20339僅生成了10 mg/L的丙氨酸。因此,LBM 20027不僅在風味化合物4-乙基愈創木酚合成方面具有優良特性,其轉化天冬氨酸為丙氨酸的能力很強,是有潛力的工程菌種。
2.3.4 關于生物胺產生的安全性分析
已有報道顯示,嗜鹽四聯球菌在合成組胺時具有菌株水平特異性,即有些嗜鹽四聯球菌會利用組氨酸合成對人體健康有害的組胺,而有些菌株不具有上述催化反應。本研究通過液相檢測組胺濃度,從而對菌株LBM 20027的組胺合成能力進行檢測。結果如圖3-d所示,以具有組胺合成能力的嗜鹽四聯球菌LBM 20159作為對照,經過7 d發酵,組胺產生菌LBM 20159生成組胺602.5 mg/L,而LBM 20027無法合成組胺。因此,LBM 20027在組胺形成方面具有生物安全性。
2.3.5 菌株耐受性檢測
本研究對菌株LBM 20027耐受酸性及NaCl能力進行檢測。結果如圖3-e、圖3-f所示,LBM 20027在pH 6.0~7.0時具有良好的生長性能,可判定該菌株的最適pH值為6.0~7.0,在pH 5.0以下生長性能較弱。相較于模式菌株DSM 20339,菌株LBM 20027在酸性環境下顯示出更好的耐受性。菌株的耐鹽能力如圖3-f所示,可以看出菌株的生長能力隨著NaCl濃度的升高而降低,在6%(質量分數)NaCl條件下,展現出最佳生長性能,與模式菌株DSM 20339相比,生長能力略高。
2.4 模擬發酵
2.4.1 基本理化特征
如上所示,菌株LBM 20027在四乙基愈創木酚合成及天冬氨酸轉化方面具有顯著優勢。本研究將該菌株作為模擬醬油起始發酵菌劑,來探究優質菌劑添加對醬油品質的影響。本研究通過模擬醬油發酵實驗,在發酵第0天添加106CFU/mL的菌株LBM 20027,在發酵0、10、20、30、40 d取樣,對模擬高鹽稀態醬油發酵過程中的基本理化指標pH、總酸、氨基酸態氮以及四乙基愈創木酚進行檢測,來檢驗LBM 20027的發酵性能。
如圖4所示,嗜鹽四聯球菌作為起始發酵菌劑被加入醬油發酵體系中后,pH略有下降(圖4-a),在發酵前10 d,醬醪pH迅速降低,10 d后pH降低速度變緩,到20 d趨于穩定,此時pH值為4.4;總酸變化如圖4-b所示,在發酵前10 d,快速提升,后續趨于穩定,達到11 g/100 mL,相較于空白提升了22.2%。氨基酸態氮變化如圖4-c所示,前10 d增長迅速,而10 d以后趨于穩定,發酵結束時達到0.5 g/100 mL,比空白組略高。

a-pH變化;b-總酸變化;c-氨基酸態氮變化;d-四乙基愈創木酚變化圖4 模擬發酵過程理化特征及風味化合物變化Fig.4 Physicochemical characteristics and changes of flavor compounds during simulated fermentation
2.4.2 模擬發酵過程中四乙基愈創木酚變化
在模擬醬油發酵過程中,對四乙基愈創木酚的含量進行追蹤,結果如圖4-d所示。四乙基愈創木酚在醬油模擬發酵過程前20 d呈現快速增長的趨勢,20 d后增長趨于平穩,暗示嗜鹽四聯球菌LBM 20027由于具有良好的耐鹽耐酸性能,在發酵前20 d成功定植并發揮風味物質合成能力。如圖4-d所示,在發酵40 d時,與添加模式菌株DSM 20339和不接發酵劑的空白組比較,接種嗜鹽四聯球菌LBM 20027的模擬醬油發酵樣品中四乙基愈創木酚含量分別是它們的1.44和3.34倍。
目前國際上已報道的具有高產四乙基愈創木酚能力的微生物主要以芽孢桿菌和酵母菌為主。郭明威等[14]報道了一株枯草芽孢桿菌(Bacillussubtilis),其四乙基愈創木酚產量達到29 mg/L;王少磊等[20]報道了一株解淀粉芽孢桿菌(Bacillusamyloliquefaciens),其四乙基愈創木酚產量達到33 mg/L;鄒謀勇等[21]報道了一株擬威克酵母(Wickerhamiellaversatilis),其四乙基愈創木酚產量達到630 mg/L。如上四乙基愈創木酚高產菌種有潛力高效提升醬油中四乙基愈創木酚含量。然而,醬油的濃郁風味來源于包括四乙基愈創木酚在內的多種風味成分而非單一成分。本研究篩選的高產四乙基愈創木酚的菌種為嗜鹽四聯球菌,該菌種為醬油釀造過程中的優勢微生物,貫穿整個醬油發酵周期,且能發酵生成多種風味成分或風味物質前體,已作為一種優良的起始發酵劑用于在日韓高鹽發酵食品生產中。如圖1及圖4-d所示,相比于嗜鹽四聯球菌的模式菌株DSM 20339,嗜鹽四聯球菌LBM 20027在單菌發酵及模擬醬油發酵中均展現了高效的四乙基愈創木酚合成能力。
2.4.3 揮發性物質
為了驗證接種嗜鹽四聯球菌LBM 20027對醬油發酵過程整體揮發性物質的影響,本研究對發酵樣品進行了GC-MS檢測。結果如圖5所示,在接種樣品組中,發現酯類和酸類化合物含量明顯高于空白組,而高濃度的酸類物質有助于濃郁的酯香形成,如乙酸乙酯、己酸乙酯、醋酸、丁酸、異戊酸等,這些酯類化合物賦予醬油強烈的果香和酒香,同時愈創木酚、四乙基愈創木酚、四乙烯基愈創木酚的含量接種LBM 20027的組別高于接種DSM 20339的樣品組,賦予醬油其獨特的醬香特征。但空白組中部分醇類化合物高于接種組,如乙醇、異丁醇、正己醇、3-甲硫基丙醇等。接種嗜鹽四聯球菌可以有效提高包括四乙基愈創木酚在內的各種風味物質含量,且接種LBM 20027的效果更為明顯。

圖5 模擬發酵過程揮發性化合物熱圖Fig.5 Heat map of volatile compounds in simulated fermentation process
3 結論
本研究對高鹽發酵食品的醬醪進行分離篩選,在高鹽環境下篩選出一株具有高四乙基愈創木酚轉化能力的耐鹽乳酸菌LBM 20027,通過對其形態和分子生物學鑒定,發現該菌LBM 20027為嗜鹽四聯球菌。將該菌株LBM 20027與嗜鹽四聯球菌模式菌株DSM 20339相比,LBM 20027的耐酸和耐鹽能力更加出色,不產生組胺,具有更好的四乙基愈創木酚轉化能力和天冬氨酸轉化能力。將菌株應用于高鹽稀態醬油模擬發酵,結果顯示,在模擬體系中菌株LBM 20027也具有更好的四乙基愈創木酚產生能力,上述結果顯示出該菌株改善醬油風味的巨大潛力。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
