化學遺傳學調控高血壓大鼠室旁核促腎上腺皮質激素釋放因子神經元對前交感神經元興奮性的影響
2023-12-12 08:18:38馬宏宇郭鑫淇杜梓碩王浩然馬會杰
中國藥理學通報 2023年12期
關鍵詞:高血壓
馬宏宇,郭鑫淇,張 熒,高 璐,杜梓碩,王浩然,馬會杰
(河北醫科大學基礎醫學院生理學教研室,河北 石家莊 050017)
根據“十二五”期間高血壓抽樣調查數據,中國成年人(18歲以上)高血壓(收縮壓≥140 mmHg/舒張壓≥90 mmHg,1 mmHg=0.133 kPa,或者服用高血壓治療藥物超過2周)患病率約為23.2%,即我國約有2.45億高血壓患者[1]。高血壓作為一種常見的心血管疾病,不僅患病率高,還會引起嚴重的心、腦、腎等并發癥,是腦卒中、冠心病等心血管疾病的主要危險因素。下丘腦室旁核(paraventricular nucleus, PVN)作為調節神經內分泌功能和心血管交感緊張性的關鍵腦區[2],含有加壓素能和催產素能大細胞、神經內分泌性小細胞以及調節交感神經活動的前交感神經元[3]。既往研究表明,PVN投射于延髓頭端腹外側區(rostral ventrolateral medulla, RVLM)和脊髓中間外側柱(intermediolateral column, IML)的前交感神經元,在調節交感傳出活動和血壓水平中起重要作用[4]。高血壓發生時PVN前交感神經元過度興奮,然而其過度興奮的原因仍未知。相關研究顯示,自發性高血壓大鼠(spontaneously hypertensive rat, SHR)PVN促腎上腺皮質激素釋放因子(corticotropin-releasing factor, CRF)神經元數量和CRF mRNA水平明顯升高[5],然而CRF神經元過度興奮對前交感神經元興奮性的影響尚不清楚。
化學遺傳學主要是通過對G蛋白偶聯受體(G protein coupled receptor, GPCR)進行改造,使其成為只能被外源性配體特異性激活的受體,即只由設計性藥物激活的設計性受體(designer receptors exclusively activated by designer drugs, DREADDs)[6]。目前大量研究證明,以DREADDs為基礎的化學遺傳學可用于操控各類神經元的興奮性,如谷氨酸能神經元、γ氨基丁酸(γ-aminobutyric acid, GABA)能神經元和多巴胺能神經元[7]。在此基礎上,配合CRF神經元啟動子特異性腺相關病毒可將上述DREADDs序列引入CRF神經元特異性區域并大量表達。典型的DREADDs有Gq、Gi、Gs、Golf和β-arrestin,化合物疊氮平-N-氧化物(clozapine-N-oxide, CNO)作為配體能夠激活上述受體,進而觸發不同的GPCR級聯反應[8]。其中應用最廣泛的是Gq受體偶聯的人M3毒蕈堿型DREADD(hM3Dq)和Gi受體偶聯的人M4毒蕈堿型DREADD(hM4Di)兩種。CNO作為配體與hM3Dq結合后,可通過磷脂酶C(phospholipase C, PLC)系統相關通路引起神經元興奮效應;與hM4Di結合后,可通過腺苷酸環化酶(adenylate cyclase, AC)系統相關通路引起神經元抑制效應。
本研究旨在觀察室旁核CRF蛋白表達水平以及CRF神經元數量,應用化學遺傳學技術調控大鼠PVN CRF神經元興奮性,使用全細胞膜片鉗技術觀察對PVN前交感神經元興奮性的影響,驗證高血壓時CRF神經元通過神經微環路對前交感神經元的調控作用。
1 材料與方法
1.1 動物SPF級正常血壓Wistar Kyoto大鼠和SHR大鼠購自北京維通利華實驗動物技術有限公司,體質量260 g~280 g,動物許可證號:SCXK(京)2016-0006。動物分籠飼養,自由飲食飲水,環境溫度18 ℃~24 ℃,相對濕度45%~55%,每天光照時間12 h,所有實驗均經河北醫科大學實驗動物使用倫理委員會審核批準。實驗過程中所有的動物均按照動物倫理學要求處置。
1.2 主要試劑CRF抗體購自中國武漢愛博泰克公司,β-actin抗體購自中國武漢Proteintech公司,CyTM3結合的羊抗兔抗體購自美國Jackson ImmunoResearch,化學遺傳學腺相關病毒AAV(PHP.eB)-hsyn-DIO-hM3D(Gq)-eGFP和AAV(PHP.eB)-hsyn-DIO-hM4D(Gi)-mCherry購自上海吉凱基因技術有限公司;CRF神經元啟動子特異性腺相關病毒AAV1/2-Rat CRF promoter-Cre購自Genedetect;熒光示蹤劑FluoSpheres購自Invitrogen,DiI購自Sigma;其他生化試劑均為進口分裝或國產分析純。
1.3 蛋白印跡實驗檢測PVN CRF表達使用2%~3%的異氟烷麻醉WKY和SHR大鼠,斷頭取出腦組織,切除小腦、腦干以及前1/3端腦,剩余部分以及腦干部分迅速放入低溫(0~4 ℃)的0.9%生理鹽水中,使用振動切片機(VT1000S, Leica, Germany)切取包含PVN腦區的冠狀面腦片(600 μm),隨即根據圖譜位置取PVN放入凍存管中,在液氮中快速冷凍。提取各組樣本總蛋白,每組上樣量為20 μg,經10% SDS-PAGE凝膠電泳后電轉移至PVDF膜。5%脫脂奶粉室溫封閉2 h,加入兔抗CRF抗體(1 ∶1 000),室溫孵育3 h。TBST洗膜后加羊抗兔IgG(1 ∶5 000),室溫孵育1 h,TBST洗膜,滴加 ECL發光劑,于 ChemiScope S6成像系統中顯影,并對條帶進行定量分析。以目的條帶和β-actin條帶灰度值比值作為最終結果。
1.4 免疫熒光實驗腹腔注射20%的烏拉坦(800 mg·kg-1)麻醉大鼠,依次灌注肝素生理鹽水與4%多聚甲醛-PBS溶液,灌注結束后斷頭取出腦組織,在4%多聚甲醛-PBS溶液中固定1 h。將腦組織放入30%蔗糖-PBS溶液中浸泡脫水24~48 h,加入包埋劑OCT包埋腦組織,根據腦圖譜調整切片的角度,冠狀面切取腦組織切片,切片厚度為25 μm。使用5%牛血清白蛋白室溫封閉1 h,洗去封閉液,加入一抗CRF(1 ∶100),4 ℃孵育過夜。PBS漂洗后加CyTM3結合的羊抗兔IgG(1 ∶500),室溫孵育2 h。使用共聚焦顯微鏡觀察腦片的熒光染色情況,在PVN腦區位置攝取熒光圖像(FV31S, Olympus, Japan),獲取的熒光圖像使用ImageJ軟件統計熒光細胞數量進行分析。
1.5 化學遺傳學病毒注射使用2%~3%的異氟烷麻醉WKY和SHR大鼠,并將大鼠頭部固定于腦立體定位儀。用微精密鉆頭打磨前囟后方顱骨,參照圖譜使用Nanoject II將化學遺傳學病毒微量注入雙側PVN(前囟后1.8 mm,旁開0.5 mm,深度7.8 mm),每側50 nL,注射完成后留針3~5 min。我們將兩種不同的病毒pAAV-hsyn-DIO-hM3D(Gq)-eGFP和AAV1/2-Rat CRF promoter-Cre微量注射入WKY大鼠的PVN腦區。同樣,將pAAV-hsyn-DIO-hM4D(Gi)-mCherry和AAV1/2-Rat CRF promoter-Cre微量注射入SHR的PVN腦區。術后2~3周,兩種病毒同時表達擴增,通過Cre重組酶的作用在WKY大鼠室旁核CRF神經元上表達興奮性hM3Dq-eGFP,SHR大鼠室旁核CRF神經元上表達抑制性hM4Di-mCherry。
1.6 PVN-IML投射神經元的逆行示蹤標記使用2%~3%異氟烷麻醉表達興奮性hM3Dq-eGFP的WKY大鼠和表達抑制性hM4Di-mCherry的SHR大鼠,并暴露其T1-T4脊髓節段。將熒光示蹤劑(FluoSpheres fluorescent microsphere suspension, #F8795, Invitrogen, or DiI, #468495, Sigma, USA)微量注入脊髓IML,每側3個注射位點,每次注射100 nL,注射完成后縫合傷口,待大鼠蘇醒后放回原籠喂養,5~7 d后熒光示蹤劑會被注射部位的神經軸突末梢逆行運輸到向脊髓IML投射的PVN前交感神經元胞體。
1.7 腦片的制備使用2%~3%的異氟烷麻醉WKY和SHR大鼠,斷頭取腦組織,迅速將腦組織置于冰冷的人工腦脊液(artificial cerebral spinal fluid, aCSF)中。aCSF成分如下(單位:mmol·L-1):126.0 NaCl;3.0 KCl;1.2 NaH2PO4;11.0 Glucose;26.0 NaHCO3;1.5 MgCl2;2.4 CaCl2。使用振動切片機制備厚300 μm的包含PVN腦區的冠狀面腦片,腦片于人工腦脊液中34 ℃恒溫孵育1 h,然后移至記錄槽中進行電生理記錄。
1.8 腦片膜片鉗電生理學記錄全細胞膜片鉗所用的玻璃微電極尖端阻抗為3~6 MΩ,記錄電極內電極內液的組分(單位:mmol·L-1)140 K-gluconate,2 KCl,3 MgCl2,10 HEPES, 5 Phosphocreatine,2 K-ATP,0.2 Na-GTP,用KOH調整pH值為7.4,滲透壓為290~300 mOsmol·L-1,制備好的腦片標本以恒溫恒速(34 ℃,3 mL·min-1)持續灌流。在正置顯微鏡(OlympusBX51WI)下,準確定位第三腦室雙側具有熒光標記的PVN前交感神經元,經過全細胞電壓鉗記錄,保持電位為-60 mV,用人工腦脊液持續灌流,記錄自發興奮性突觸后電流(spontaneous excitatory postsynaptic currents,sEPSCs),穩定記錄9 min后,灌流液中加入CNO (10 μmol·L-1) 灌流6 min,然后沖洗恢復至基礎狀態。全細胞電流鉗記錄PVN前交感神經元的膜電位和電流誘發的動作電位,通過記錄電極給予系列躍階電流(0 pA-200 pA,增幅20 pA,時程500 ms)刺激,記錄在每個躍階電流細胞膜電位和動作電位的變化,待波形穩定后,灌流液中加入CNO (10 μmol·L-1) 灌流6 min,然后沖洗恢復至基礎狀態。
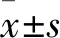
2 結果
2.1 SHR室旁核CRF蛋白表達水平升高相比于WKY大鼠,SHR室旁核CRF蛋白表達水平明顯升高(P<0.01, Fig 1A,B),說明高血壓時PVN CRF神經元合成CRF增多。

Fig 1 The protein expression level of CRF was
2.2 SHR PVN CRF陽性神經元和脊髓投射的PVN前交感神經元數量增加通過免疫熒光實驗,我們觀察到相比于WKY大鼠,SHR室旁核的CRF神經元數量明顯增加 (P<0.05, Fig 2A,B)。同時,SHR向脊髓投射的熒光標記的PVN前交感神經元數量增多,且分布于CRF陽性神經元周圍 (P<0.05, Fig 2A,C)。
2.3 PVN CRF神經元化學遺傳學DREADDs的表達及PVN前交感神經元逆行投射熒光示蹤劑的標記預先將CRF啟動子特異性Cre依賴的AAV與化學遺傳學AAV混合微量注入大鼠PVN,14 d后,通過將熒光示蹤劑注射到IML,逆行示蹤PVN前交感神經元。最后在WKY大鼠PVN可見表達hM3Dq-eGFP的CRF神經元(綠色)以及含有DiI的前交感神經元(紅色)(Fig 3A,B,D)。同樣,在SHR大鼠PVN可見表達hM4Di-mCherry的CRF神經元(紅色)以及含有FluoroSpheres的前交感神經元(綠色)(Fig 3A,C,E)。

Fig 2 Numbers of CRF-expressing neurons and presympathetic neurons increased in PVN of

Fig 3 PVN CRF neurons expressed with chemogenetic DREADDs and PVN presympathetic neurons retrogradely labeled with fluorescent tracer in WKY rats and SHR
2.4 CNO抑制SHR大鼠PVN CRF神經元可降低前交感神經元sEPSCs的頻率應用全細胞電壓鉗記錄大鼠腦片PVN前交感神經元的sEPSCs,觀察灌流液中給予CNO作用于抑制性hM4Di對sEPSCs的影響。與WKY大鼠相比,SHR大鼠PVN前交感神經元sEPSCs的頻率更高(P<0.01, Fig 4A-C),但幅度并無明顯差異。在PVN CRF神經元提前表達hM4Di-mCherry的SHR大鼠腦片,用含CNO(10 μmol·L-1)的人工腦脊液灌流腦片可引起PVN前交感神經元sEPSCs頻率明顯降低(P<0.01, Fig 4B,C),但不影響其幅度(Fig 4B,D)。
然而,對于在PVN CRF神經元提前表達hM3Dq-eGFP的WKY大鼠腦片,用含CNO的人工腦脊液灌流腦片對PVN前交感神經元sEPSCs的頻率和幅度均無明顯影響(Fig 4A, C,D)。以上數據表明,SHR大鼠PVN前交感神經元過度激活,通過化學遺傳學技術抑制CRF神經元興奮性能夠有效抑制SHR大鼠PVN前交感神經元的過度激活。
2.5 CNO調控PVN CRF神經元興奮性對前交感神經元電流誘發的動作電位無影響應用全細胞電流鉗記錄WKY和SHR大鼠PVN前交感神經元電流誘發的動作電位 (Fig 5A)。與WKY大鼠相比,SHR大鼠PVN前交感神經元的膜電位更加去極化(P<0.01, Fig 5B-D)。同時,SHR大鼠PVN前交感神經元電流誘發動作電位的頻率和放電數量也明顯高于WKY大鼠(P<0.01, Fig 5B-F)。但是在PVN CRF神經元提前表達hM3Dq-eGFP的WKY大鼠或表達hM4Di-mCherry的SHR大鼠腦片,CNO(10 μmol·L-1)灌流給藥對PVN前交感神經元電流誘發的動作電位均無明顯影響(Fig 5B-F)。以上數據表明,通過化學遺傳學技術調控CRF神經元興奮性對WKY和SHR大鼠電流誘發的動作電位無明顯影響。

Fig 4 Effect of chemogenetic manipulation of CRF neurons on sEPSCs of presympathetic neurons in PVN in WKY rats and

Fig 5 Effect of chemogenetic manipulation of CRF neurons on current-evoked firing of PVN presympathetic neurons in WKY rats and
3 討論
高血壓發病率居高不下,患病人群更趨于年輕化,因此闡明高血壓發生的潛在神經機制尤為迫切。既往研究表明,前交感神經元過度興奮引起交感傳出活動增強[9]是高血壓發生的重要神經機制。我們的研究結果顯示,SHR PVN前交感神經元sEPSCs頻率以及電流誘發放電活動明顯高于WKY,表明,PVN前交感神經元過度興奮在高血壓發生時起到重要作用。研究表明PVN作為調節神經內分泌功能和心血管交感緊張性的關鍵腦區[2],其CRF神經元數量和CRF mRNA水平在高血壓患者[10]和SHR[5]中均明顯升高。因此,我們推測SHR大鼠PVN前交感神經元興奮性的變化與CRF神經元過度激活和內分泌活動增強密切相關。
存在于突觸前囊泡中的興奮性神經遞質以量子級釋放并作用于突觸后膜相關受體,引發的突觸后膜內向電流即為sEPSCs,反映的是由突觸前興奮性神經遞質釋放變化而引發的突觸后神經元興奮性變化。電流誘發動作電位是向神經元注入電流而誘發的動作電位,反映神經元的可興奮性。我們通過化學遺傳學技術抑制自發性高血壓大鼠PVN CRF神經元,可有效降低前交感神經元sEPSCs的頻率,但對電流誘發的動作電位無明顯影響。因此,我們認為在高血壓發生時CRF神經元興奮性增強是前交感神經元過度興奮的主要原因之一。但PVN CRF神經元激活對高血壓大鼠PVN前交感神經元sEPSCs和電流誘發動作電位的反應性差異,有待于進一步研究。
此外,前交感神經元的過度激活與興奮性谷氨酸能傳入增強、抑制性GABA能傳入減弱密切相關[11-12]。而這兩類中間神經元上也有CRF受體(CRFR)分布,CRF作為腦內重要神經調質可調節其興奮性[13]。PVN CRF神經元可通過釋放CRF,作用于CRFR發揮神經內分泌功能[14-15]。兩種類型的CRF受體,即CRFR1和CRFR2,均在PVN高表達,并與自主神經活動的調節密切相關[16]。高血壓時室旁核內源性CRF增多,通過CRFR1增強谷氨酸能神經元的興奮性,進而引起增強前交感神經元的興奮性[5]。然而,抑制SHR大鼠CRF神經元改善前交感神經元過度興奮后,是否可改善增強的交感傳出活動與升高的血壓,仍需進一步探究。
綜上所述,自發性高血壓大鼠PVN CRF神經元與前交感神經元之間存在神經微環路,PVN CRF神經元興奮性增強是前交感神經元過度興奮的重要原因之一,本研究為高血壓的防治提供了實驗依據。
猜你喜歡
青春期健康(2022年19期)2022-10-12 16:46:40
西部醫學(2021年10期)2021-10-28 08:25:50
中老年保健(2021年5期)2021-08-24 07:07:16
中老年保健(2021年11期)2021-08-22 03:14:10
今日農業(2020年19期)2020-12-14 14:16:52
中國生殖健康(2020年6期)2020-02-01 06:29:06
中國生殖健康(2018年6期)2018-11-06 07:09:44
基層中醫藥(2018年4期)2018-08-29 01:25:58
基層中醫藥(2018年6期)2018-08-29 01:20:14
中國繼續醫學教育(2015年2期)2016-01-06 01:36:28