陸地棉轉錄因子WRKY29的鑒定及表達分析
2023-12-14 04:43:51劉文豪余渝王旭文田琴吳珂馬麒趙福相孔憲輝
安徽農業科學 2023年23期
劉文豪 余渝 王旭文 田琴 吳珂 馬麒 趙福相 孔憲輝
摘要
[目的]揭示WRKY29基因表達機理,為分子輔助育種創新種質提供基因資源。[方法]以WRKY家族成員中的WRKY29為目標,通過生物信息學方法分析陸地棉WRKY29轉錄因子的理化性質、系統進化關系、亞細胞定位建模和染色體分布定位、基因的組織特異性表達。[結果]生物信息學分析表明,陸地棉WRKY29轉錄因子含有279個氨基酸殘基,理論等電點為6.08,具有典型的WRKY保守結構域,屬于不穩定蛋白。蛋白質二級結構以α螺旋和無規卷曲為主。系統進化關系分析得知WRKY29與同為錦葵目的物種聚類在同一進化分支,亞細胞定位預測WRKY29轉錄因子分布在細胞核,而染色體定位發現WRKY29轉錄因子分布在A02染色體40~48 Mb區域內。WRKY29轉錄因子具有正調控功能,組織表達模式發現在根部具有顯著性表達。[結論]陸地棉轉錄因子WRKY29的鑒定及表達分析為棉花生長發育過程中的抗逆研究提供了重要線索。
關鍵詞 WRKY29;轉錄因子;生物信息學;育種
中圖分類號 S 562? 文獻標識碼 A? 文章編號 0517-6611(2023)23-0085-05
doi:10.3969/j.issn.0517-6611.2023.23.020
Identification and Expression Analysis of Transcription Factor WRKY29 in Gossypium hirsutum L.
LIU Wen-hao, YU Yu, WANG Xu-wen et al
(Cotton Research Institute, Xinjiang Academy of Agricultural and Reclamation Science/Northwest Inland Region Key Laboratory of Cotton Biology and Genetic Breeding, Ministry of Agriculture and Rural Affairs/State Key Laboratory of Cotton Biology, Shihezi, Xinjiang 832000)
Abstract [Objective]In the early stage, our research team identified a WRKY29 gene in upland cotton, and revealed the mechanism of gene expression through analysis, hoping to provide genetic resources for molecular-assisted breeding innovative germplasm.[Method]This study targeted WRKY29, a member of the WRKY family, and analyzed the physicochemical properties, phylogenetic relationships, subcellular location modeling and chromosome distribution location, and tissue specific expression of WRKY29 transcription factors in Gossypium hirsutum L. by bioinformatics methods.[Result]Bioinformatics analysis showed that the upland cotton WRKY29 transcription factor contains 279 amino acid residues, the theoretical isoelectric point is 6.08, and has a typical WRKY conserved domain, which is an unstable protein. The secondary structure of WRKY29 was mainly α-helix and random coil. Phylogenetic analysis showed that WRKY29 and mallow species clustered in the same evolutionary branch. Subcellular localization predicted that WRKY29 was located in the nucleus, chromosome mapping showed that WRKY29 transcription factor was distributed in the 40-48 mb region of A02 chromosome. WRKY29 transcription factor has a positive regulatory function, and the tissue expression pattern was found to be significantly expressed in the roots.[Conclusion]Identification and expression analysis of transcription factor WRKY29 in upland cotton provide important clues for the study of resistance to stress during the growth and development of cotton.
Key words WRKY29;Transcription factor;Bioinformatics;Breeding
基金項目 國家重點研發計劃項目“西北內陸優質機采棉新品種培育-機采優異育種資源的創制”(2017YFD0101601);兵團重點領域創新團隊建設計劃項目“機采棉遺傳育種與高效栽培創新團隊”(2017CB011);棉花生物學國家重點實驗室開放課題“縮節胺拌種對棉花生長發育轉錄組分析”(CB2022A27)。
作者簡介 劉文豪(1992—),男,安徽界首人,助理研究員,碩士,從事棉花育種研究。
*通信作者,研究員,從事棉花育種研究。
收稿日期 2022-07-15
WRKY基因家族是含有WRKY保守結構域的轉錄因子家族,其廣泛存在于植物體中,并且在植物生長發育、逆境脅迫應答和代謝調控等一系列生理生化過程中發揮重要作用[1]。基于全基因組的WRKY基因家族結構和功能分析在許多已測序的物種中得到了廣泛應用[2]。目前已完成擬南芥[3]、水稻[4]、蘿卜[5]、小麥[6]和毛果楊[7]等多個物種的WRKY基因家族分析。棉花是我國重要的經濟作物之一,我國是世界上最大的棉花生產國和消費國,總產和單產均居世界首位[8],2020年全國棉花播種面積為316.99萬hm2。因此,保持我國棉花產業的健康穩定發展,對于促進農業增效,農民增收及農村經濟穩定具有重要意義。但是,近年來棉花病害較為嚴重,危害日益增加,而且缺少抗逆的高產優質棉花主推品種。因此,對抗病、抗逆優良品種的研究至關重要。優質棉的培育是解決問題的根本途徑,分析關鍵基因的調控網絡、差異表達及理化性狀具有重要意義。
有研究表明,WRKY轉錄因子家族成員不僅在植物的生物脅迫和非生物脅迫中起重要作用,還參與碳水化合物的合成、次生代謝產物的合成、植株的衰老和發育[9]。WRKY蛋白能與目標基因啟動子中的W-box(TGACC(A/T))結合,激活或抑制下游基因的表達,調節其應激反應。此外,WRKY蛋白可以與其他轉錄因子相互作用,調節植物的防御反應[10]。Chen等[11]的研究表明,擬南芥Ⅲ類WRKY轉錄因子WRKY46、WRKY54和WRKY70都參與植物甾體激素(BR)調控植物生長和干旱反應。Duan等[12]研究發現,毛白楊PtrWRKY73在植物對生物營養型病原菌的抗性中起著積極作用,但在對壞死營養型病原菌的抗性中起著消極作用。目前很多研究都集中在WRKY基因的功能鑒定上,但是對WRKY基因生物學許多領域的認識有限。如棉花中WRKY基因的系統發育特征、整體表達模式、調控表達的分子機制和途徑等都沒有得到很好的描述[13]。
迄今為止,很多陸地棉WRKY家族的成員被分析研究,但是陸地棉WRKY29轉錄因子尚未被綜合分析,其在陸地棉對生物和非生物脅迫的反應過程中發揮著的作用尚未得知,其涉及的調控網絡及差異表達尚不清楚。筆者以陸地棉標準系TM-1基因組數據庫[14]和轉錄組數據庫PRJNA248163進行WRKY29轉錄因子的序列獲取及差異表達分析,使用多類型生物信息學工具進行理化性質、亞細胞定位、調控網絡等相關的分析,以期為基因功能研究及生物技術培育優質棉花新品種奠定理論基礎。
1 材料與方法
1.1 GhWRKY29蛋白理化性質分析
以擬南芥AtWRKY29(AT4G23550)為探針,從陸地棉標準系TM-1基因組數據庫獲取GhWRKY29基因的cds和蛋白序列,在NCBI網站(https:∥www.ncbi.nlm.nih.gov/)進行比對,SMART網站(http:∥smart.embl.de/)進行結構域的預測分析。使用ProtParam網站(https:∥web.expasy.org/protparam/)分析氨基酸數、分子量、理論等電點、帶正負電荷的氨基酸殘基、分子式、總原子數、不穩定系數、脂肪指數、親疏水性等。
1.2 GhWRKY29蛋白的結構、系統進化關系和序列比對分析
SOPMA網站(https:∥npsa-prabi.ibcp.fr/)輸入氨基酸序列分析GhWRKY29蛋白的二級結構,包括α螺旋、β轉角、延伸鏈和無規卷曲。在線網站SWISS-MODEL(https:∥swissmodel.expasy.org/)分析GhWRKY29蛋白的三級結構。NCBI獲取不同物種的WRKY29蛋白序列和登錄號,使用MEGA 7.0軟件進行系統進化關系分析,用EvolView(http:∥evolgenius.info/)進行系統發育樹的編輯。
1.3 GhWRKY29蛋白亞細胞定位和染色體圖譜繪制
在線網站Predict Protein(https:∥www.predictprotein.org)進行GhWRKY29蛋白的亞細胞定位分析。從陸地棉標準系TM-1基因組數據庫獲取GhWRKY29的基因信息,使用在線網站MG2C(http:∥mg2c.iask.in/mg2c_v2.1/)進行染色體定位的圖譜繪制,chromosome,chromosome id,gene lines,gene id等指標僅變動字體格式,其他均為默認參數。
1.4 GhWRKY29基因的表達模式分析
從NCBI網站下載陸地棉TM-1標準系轉錄組數據包(PRJNA248163),結合
陸地棉轉錄組數據庫,對數據進行矩陣轉換,獲取GhWRKY29
基因在不同組織表達的FPKM值,進一步分析
GhWRKY29基因
的特異表達情況。使用TB-Tools軟件的Super HeatMap
Browser功能進行繪制基因差異表達熱圖,Adobe Illustrator 2019軟件對圖片進行完善。
2 結果與分析
2.1 GhWRKY29蛋白理化性質
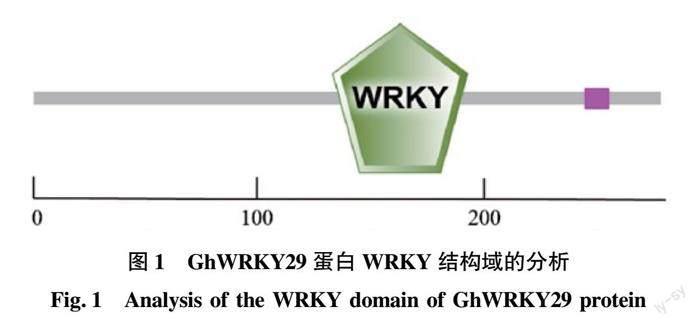
以擬南芥AtWRKY29(AT4G23550)為探針,在陸地棉TM-1標準系基因組數據庫中搜索得到相對應的Gh_A04G1042的蛋白序列和CDS序列。SMART網站結構分析該序列含有分布在第133~193氨基酸區域的WRKY結構域(圖1),該結構域是一個含有60個氨基酸的區域,由一個保守的氨基酸序列WRKYGQK在其N端定義,并帶有一個新穎的鋅指狀基序[15]。根據基因組數據庫注釋信息和NCBI的Blast比對數據確定為GhWRKY29。ProtParam分析結果表明,氨基酸數為279個,分子量為31 899.58,理論等電點為6.08,帶負電荷的氨基酸殘基(Asp+Glu)有41個,帶正電荷的氨基酸殘基(Arg+Lys)有36個。分子式為:C1413H2168N398O434S7,總原子數是4 420,不穩定系數為51.41,是不穩定蛋白。脂肪指數是60.07,親水性的平均值(GRAVY)是-0.814。
2.2 GhWRKY29蛋白的二級結構和三級結構
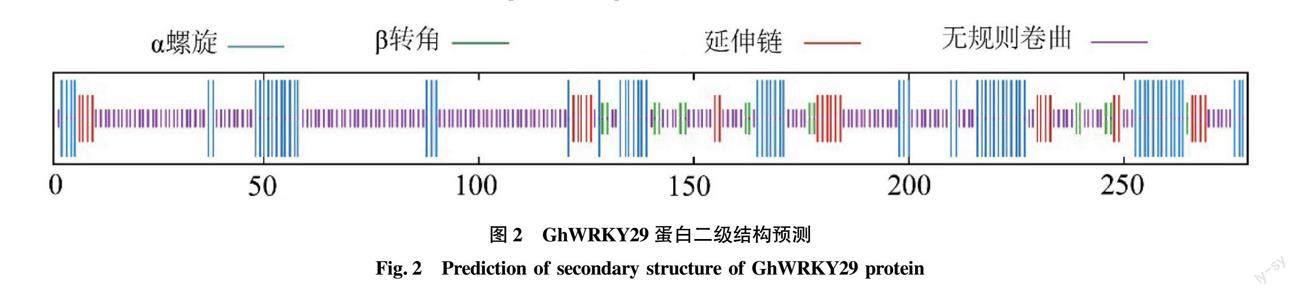
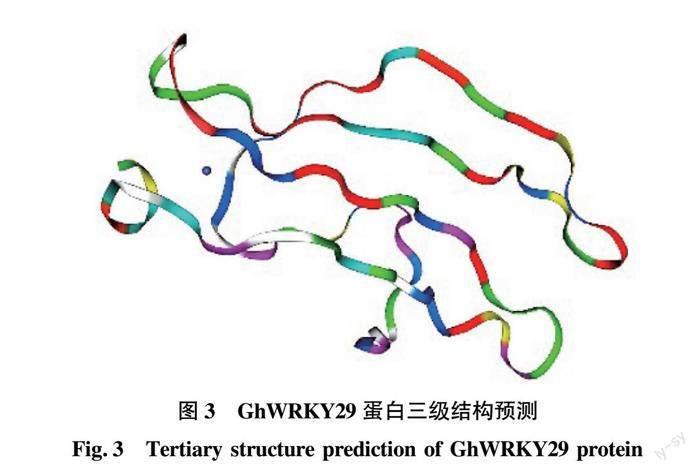
SOPMA分析發現(圖2),GhWRKY29的蛋白質序列含有279個氨基酸,而在這些氨基酸的分布排列中α螺旋68個,占比為24.37%;β轉角15個,占比為5.38%;延伸鏈27個,占比為9.68%;無規卷曲169個,占比為60.57%。基于結構基因組學,把WRKY29的蛋白序列輸入在線網站SWISS-MODEL,通過搜索序列結構模型相同的模板建立骨架,優化模型,得到該蛋白的三級結構的Cartoon模型,而這個模型的氨基酸排列順序與二級結構預測基本相符,進一步驗證了WRKY29蛋白結構的準確性(圖3)。
2.3 GhWRKY29蛋白進化關系及序列結構
從NCBI網站獲取不同物種WRKY29蛋白的氨基酸序列和登錄號,分別是甘草GgWRKY29(QFI57424.1)、擬南芥AtWRKY29(OAO99222.1)、岷江百合LrWRKY29(QRX38926.1)、核桃JrWRKY29(ALU11217.1)、蕪青BrWRKY29(AHB33835.1)、麻風樹JcWRKY29(AGQ04217.1)、煙草NaWRKY29(OIT33212.1)、甜櫻桃PaWRKY29(XP_021820806.1)、木槿HsWRKY29(XP_039015885.1)、榴蓮DzWRKY29(XP_022737617.1)和梨PuWRKY29(KAB2632799.1)。通過系統
發育進化樹對陸地棉WRKY29蛋白進行聚類分析(圖4),結果顯示,該聚類分析分為Group A和Group B兩部分,Group A包括陸地棉GhWRKY29、木槿HsWRKY29和榴蓮DzWRKY29 3個物種,Group B包括甘草GgWRKY29、擬南芥AtWRKY29、岷江百合LrWRKY29、核桃JrWRKY29、蕪青BrWRKY29、麻風樹JcWRKY29、煙草NaWRKY29、甜櫻桃PaWRKY29和梨PuWRKY29 5個物種。
通過DNAMAN軟件對不同物種的蛋白序列進行比對分析,結果表明,陸地棉WRKY29與同屬錦葵科的木槿序列一致性最高,為65.56%;與錦葵目木棉科的榴蓮序列一致性次之,為58.18%;與大戟科的麻風樹序列一致性最低,為15.12%。結構域分析結果顯示,在N端含有WRKYGQK七肽段序列的結構域,而在C端含有1個C2H2類型的鋅指結構
2.4 GhWRKY29蛋白的亞細胞和染色體定位
亞細胞定位是研究基因功能的一種重要技術手段。為準確了解GhWRKY29蛋白在表達調控中所具有的功能,該研究使用GhWRKY29蛋白的氨基酸序列,在網站Plant-mPLoc進行亞細胞定位預測,結果表明,該蛋白定位在細胞核。PredictProtein網站預測結果同樣表明該蛋白定位在細胞核中(圖6A)。從基因組數據庫查詢到該基因位于A02染色體上,依據該基因的序列起始位置和終止位置信息,用MG2C網站預測在染色體上的具體分布情況(圖6B)。這將有助于了解GhWRKY29基因在染色體上的位置和所具有生理功能之間的關系。
2.5 GhWRKY29基因的表達模式
利用陸地棉轉錄組數據庫PRJNA248163對GhWRKY29基因進行表達模式分析(圖7),轉錄組檢測組織分別為根、莖、葉。根據FPKM值分析GhWRKY29基因的表達模式,圖7a為對照植株,圖7b是該基因在根、莖、葉中的表達情況。GhWRKY29基因在陸地棉植株的根、莖、葉的表達模式存在顯著差異,在根中的表達量最高,其次是葉片,在莖中的表達量相對最低。陸地棉根系具有吸收養分、水分及固定地上部分的功能,而且還具有向土壤輸入有機質和感知根部周邊環境變化的作用。GhWRKY29基因在根部具有高表達量,推測其可能在根系響應逆境脅迫的調控網絡中具有重要功能。
3 討論
植物在長期進化過程中形成了生長發育和逆境響應的調控網絡,轉錄因子家族在該過程中起到重要作用[16]。WRKY轉錄因子家族參與干旱、鹽漬、高溫、冷凍、枯黃萎病和蟲害等多種環境脅迫的響應。Gu等[17]研究發現,棉花GhWRKY27通過減少葉綠素含量及提高衰老相關基因的過表達促進了葉片的衰老;Wang等[18]研究表明,棉花GhWRKY33介導植物對干旱脅迫的反應,并參與ABA信號通路;Chu等[19]推測GhWRKY41通過調節活性氧(ROS)清除和抗氧化劑基因的表達來增強植物對逆境的耐受性。陸地棉WRKY家族的部分成員被分析研究,而WRKY29基因的綜合研究報道較少。早期的系統發育研究主要基于生物的表型特征,通過表型比較來研究物種之間的進化關系。然而,利用表型特征進行系統發育分析存在很大的局限性。該研究以生物信息學方法從基因組數據庫獲得GhWRKY29轉錄因子的cds序列和蛋白序列,經過Blast比對、數據庫篩選及SMART網站結構分析確定了基因身份。生物信息學分析結果表明,GhWRKY29的蛋白質序列含有279個氨基酸,屬于不穩定蛋白。對該蛋白的二級序列和三級結構預測有助于了解蛋白質的生物功能。從NCBI獲取其他11個物種的WRKY蛋白序列及登錄號與GhWRKY29蛋白進行聚類分析,得知陸地棉GhWRKY29蛋白與同為錦葵目的木槿HsWRKY29和榴蓮DzWRKY29在同一進化分支,而非錦葵目的其余9個物種的WRKY29蛋白聚類在Group B的其他分支。在Group A中發現,陸地棉GhWRKY29蛋白與同為錦葵科的木槿HsWRKY29親緣關系更近,與木棉科榴蓮的親緣關系相對較遠,說明物種的進化具有保守性,這些分析增強了對自然選擇在不同物種WRKY29進化過程中所起作用的了解。亞細胞定位預測GhWRKY29定位在細胞核,參與核基因的轉錄調控。由染色體位置圖譜可知,GhWRKY29基因分布在基因組A亞組的第2條染色體的40~48 Mb,有助于了解GhWRKY29基因在染色體上的位置和生理功能之間的關系,也可用于判斷其在遺傳圖譜上的相對距離。GhWRKY29基因的表達模式分析了其在根、莖、葉中的差異表達,結果表現為根部的表達量>葉部的表達量>莖部的表達量。轉錄因子和植物激素在控制根發育的根基因調控網絡方面起著關鍵的調節作用[20]。GhWRKY29基因在根部大量表達可能涉及蛋白質運動和基因表達調控的機制。前人研究發現,擬南芥細胞核AtWRKY29與AtBAG7相互作用有助于應對脅迫及增加錯誤折疊蛋白質的積累,最終維持細胞的穩態[21]。WRKY29在小麥中的異位表達導致矮小和抽穗早,增強了對鐮刀菌赤霉病的抗病性[22]。該研究結果為進一步GhWRKY29基因克隆提供了必要的條件,為后續研究GhWRKY29基因在生物和非生物脅迫應答過程中的功能奠定了基礎。
參考文獻
[1] 伍林濤,杜才富,張敏琴,等.WRKY轉錄因子的結構及其在植物抗逆境脅迫中的功能[J].分子植物育種,2013,11(4):634-638.
[2] 任媛,趙玉潔,張心慧,等.石榴WRKY基因家族全基因組鑒定與表達分析[J].西北植物學報,2020,40(2):218-231.
[3] WANG Q S,WANG M H,ZHANG X Z,et al.WRKY gene family evolution in Arabidopsis thaliana[J].Genetica,2011,139(8):973-983.
[4] RAMAMOORTHY R,JIANG S Y,KUMAR N,et al.A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments[J].Plant Cell Physiol,2008,49(6):865-879.
[5] KARANJA B K,FAN L X,XU L,et al.Genome-wide characterization of the WRKY gene family in radish(Raphanus sativus L.)reveals its critical functions under different abiotic stresses[J].Plant Cell Rep,2017,36(11):1757-1773.
[6] GUPTA S,MISHRA V K,KUMARI S,et al.Deciphering genome-wide WRKY gene family of Triticum aestivum L.and their functional role in response to abiotic stress[J].Genes Genomics,2019,41(1):79-94.
[7] HE H S,DONG Q,SHAO Y H,et al.Genome-wide survey and characterization of the WRKY gene family in Populus trichocarpa[J].Plant Cell Rep,2012,31(7):1199-1217.
[8] 喻樹迅.我國棉花生產現狀與發展趨勢[J].中國工程科學,2013,15(4):9-13.
[9] JIANG J J,MA S H,YE N H,et al.WRKY transcription factors in plant responses to stresses[J].J Integr Plant Biol,2017,59(2):86-101.
[10] 張凡,尹俊龍,郭瑛琪,等.WRKY轉錄因子的研究進展[J].生物技術通報,2018,34(1):40-48.
[11] CHEN J N,NOLAN T M,YE H X,et al.Arabidopsis WRKY46,WRKY54,and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought responses[J].Plant Cell,2017,29(6):1425-1439.
[12] DUAN Y J,JIANG Y Z,YE S L,et al.PtrWRKY73,a salicylic acid-inducible poplar WRKY transcription factor,is involved in disease resistance in Arabidopsis thaliana[J].Plant Cell Rep,2015,34(5):831-841.
[13] DOU L L,ZHANG X H,PANG C Y,et al.Genome-wide analysis of the WRKY gene family in cotton[J].Mol Genet Genomics,2014,289(6):1103-1121.
[14] ZHANG T Z,HU Y,JIANG W K,et al.Sequencing of allotetraploid cotton(Gossypium hirsutum L.acc.TM-1)provides a resource for fiber improvement[J].Nat Biotechnol,2015,33(5):531-537.
[15] 田云,盧向陽,彭麗莎,等.植物WRKY轉錄因子結構特點及其生物學功能[J].遺傳,2006,28(12):1607-1612.
[16] 卜華虎,王曉清,任志強,等.植物WRKY轉錄因子家族基因研究進展[J].山西農業科學,2020,48(7):1158-1163.
[17] GU L J,DOU L L,GUO Y N,et al.The WRKY transcription factor GhWRKY27 coordinates the senescence regulatory pathway in upland cotton(Gossypium hirsutum L.)[J].BMC Plant Biol,2019,19(1):1-14.
[18] WANG N N,XU S W,SUN Y L,et al.The cotton WRKY transcription factor(GhWRKY33)reduces transgenic Arabidopsis resistance to drought stress[J].Sci Rep,2019,9(1):1-13.
[19] CHU X Q,WANG C,CHEN X B,et al.The cotton WRKY gene GhWRKY41 positively regulates salt and drought stress tolerance in transgenic Nicotiana benthamiana[J].PLoS One,2015,10(11):1-21.
[20] PETRICKA J J,WINTER C M,BENFEY P N.Control of Arabidopsis root development[J].Annu Rev Plant Biol,2012,63:563-590.
[21] LI Y R,WILLIAMS B,DICKMAN M.Arabidopsis B-cell lymphoma2(Bcl-2)-associated athanogene 7(BAG7)-mediated heat tolerance requires translocation,sumoylation and binding to WRKY29[J].New Phytol,2017,214(2):695-705.
[22] SAROWAR S,ALAM S T,MAKANDAR R,et al.Targeting the pattern-triggered immunity pathway to enhance resistance to Fusarium graminearum[J].Mol Plant Pathol,2019,20(5):626-640.
