致腐細(xì)菌群體感應(yīng)信號(hào)系統(tǒng)調(diào)控水產(chǎn)品腐敗變質(zhì)的研究進(jìn)展
2023-12-18 08:15:00李錦王迪陳勝軍李春生王悅齊
食品與發(fā)酵工業(yè) 2023年23期
關(guān)鍵詞:信號(hào)
李錦,王迪,陳勝軍,李春生,王悅齊
1(中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所/農(nóng)業(yè)農(nóng)村部水產(chǎn)品加工重點(diǎn)實(shí)驗(yàn)室/國(guó)家水產(chǎn)品加工技術(shù)研發(fā)中心,廣東 廣州,510300)2(上海海洋大學(xué) 食品學(xué)院,上海,201306)3(三亞熱帶水產(chǎn)研究院/海南省深遠(yuǎn)海漁業(yè)資源高效利用與加工重點(diǎn)實(shí)驗(yàn)室,海南 三亞,572018)
海洋因其豐富的資源,被喻為“藍(lán)色糧倉(cāng)”,魚(yú)類(lèi)資源已經(jīng)成為解決世界饑餓以及營(yíng)養(yǎng)缺乏的重要支撐,全世界超過(guò)45億人通過(guò)水產(chǎn)品攝入每日所需的大部分蛋白質(zhì)[1],水產(chǎn)品正逐漸成為人類(lèi)膳食的重要組成部分。2021年全國(guó)水產(chǎn)品總產(chǎn)量6 690.29萬(wàn)t,比2020年增長(zhǎng)2%,漁業(yè)捕撈產(chǎn)量和水產(chǎn)養(yǎng)殖收成均呈現(xiàn)穩(wěn)定增長(zhǎng)狀態(tài)[2]。水產(chǎn)品含有豐富的優(yōu)質(zhì)蛋白,從中攝取的蛋白質(zhì)占人均攝入總動(dòng)物蛋白質(zhì)含量的20%[3];它是人類(lèi)多不飽和脂肪酸的重要來(lái)源,多不飽和脂肪酸在人體健康中具有重要功能:提供熱量、是脂溶性物質(zhì)的載體[4]、保護(hù)心臟健康、清除血管中多余的脂類(lèi)物質(zhì)、調(diào)節(jié)免疫代謝等[5];另外,水產(chǎn)品口感特殊[6]、味道鮮美,廣受消費(fèi)者歡迎。
但水產(chǎn)品因其含水量較高,且富含營(yíng)養(yǎng)物質(zhì),極易發(fā)生腐敗變質(zhì),其中由腐敗引起的損失在我國(guó)每年約800萬(wàn)t,高達(dá)總產(chǎn)量的15%[7],腐敗的水產(chǎn)品體表黏液增多,肌肉變得松散,其品質(zhì)大打折扣[8]。其中細(xì)菌是引發(fā)水產(chǎn)品腐敗變質(zhì)最主要的因素,致腐細(xì)菌可將水產(chǎn)品中的蛋白質(zhì)、脂類(lèi)等大分子逐漸分解成氨基酸、脂肪酸等小分子化合物,并生成胺類(lèi)、硫化氫等有害物質(zhì),同時(shí)產(chǎn)生令人不愉快的氣味。水產(chǎn)品品質(zhì)與安全常用K值、揮發(fā)性鹽基氮 (total volatile basic nitrogen,TVB-N)、生物胺等生化指標(biāo)來(lái)衡量[9]。生物胺作為評(píng)價(jià)水產(chǎn)品品質(zhì)與安全的指標(biāo)之一,水產(chǎn)品中游離氨基酸經(jīng)細(xì)菌產(chǎn)生的氨基酸脫羧酶脫羧作用后生成,包括組胺、腐胺、苯乙胺等8種,生物胺組胺作為對(duì)人類(lèi)危害最大的一種生物胺,過(guò)量攝入會(huì)造成人類(lèi)的食物中毒反應(yīng),包括腹瀉、嘔吐、頭暈等[10]。美國(guó)食品藥品管理局 (Food and Drug Administration,FDA)規(guī)定魚(yú)類(lèi)產(chǎn)品中的組胺限量為50 mg/kg,我國(guó)規(guī)定青皮紅肉魚(yú)類(lèi)檢測(cè)不得高于400 mg/kg,其余限量為200 mg/kg[11]。
水產(chǎn)品的腐敗不僅嚴(yán)重影響了水產(chǎn)品的質(zhì)量安全,還給水產(chǎn)品加工業(yè)造成了巨大的經(jīng)濟(jì)損失,因此,提高水產(chǎn)品的保鮮技術(shù),減少水產(chǎn)品腐敗是我國(guó)水產(chǎn)品加工行業(yè)亟需解決的問(wèn)題之一。傳統(tǒng)的防腐方法主要是化學(xué)保鮮,包括鹽漬、酸漬、煙熏等,雖然抑制細(xì)菌生長(zhǎng)的效果明顯,殺菌效率高,但一些化學(xué)保鮮手段破壞了水產(chǎn)品品質(zhì),改變了水產(chǎn)品特有的鮮味和軟嫩的口感,同時(shí)產(chǎn)生一定程度的化學(xué)殘留,還會(huì)對(duì)環(huán)境造成污染[12]。物理保鮮技術(shù)也在水產(chǎn)品加工與貯藏中廣泛使用,低溫技術(shù)的應(yīng)用使得水產(chǎn)品腐敗這一研究領(lǐng)域取得突破性進(jìn)展,但其費(fèi)用昂貴,且耐冷腐敗菌會(huì)變?yōu)樘禺愋愿瘮【?同樣會(huì)導(dǎo)致水產(chǎn)品變色、降解和產(chǎn)生異味[13],且魚(yú)與冰表面接觸時(shí)容易造成魚(yú)體機(jī)械損傷,不均勻的低溫分布使得產(chǎn)生的冰晶大小不一,損傷肌原纖維[12];隨著科技的發(fā)展,輻照保藏成為一種新興的水產(chǎn)品保鮮手段,其具有穿透力強(qiáng)、效率高、營(yíng)養(yǎng)物質(zhì)不易流失等優(yōu)點(diǎn),但輻照技術(shù)的安全性一直是人們擔(dān)心的問(wèn)題,水產(chǎn)品在進(jìn)行輻照過(guò)程中也會(huì)產(chǎn)生多種降解物質(zhì),這些物質(zhì)的安全性有待驗(yàn)證[14];超高壓技術(shù)可保持水產(chǎn)品本身的色澤和風(fēng)味,但蛋白質(zhì)會(huì)由于高壓導(dǎo)致肌肉組織更緊密,促進(jìn)蛋白質(zhì)相互作用而發(fā)生變性[15]。
隨著消費(fèi)者日漸增長(zhǎng)對(duì)食品安全以及健康的需求,化學(xué)及一些破壞營(yíng)養(yǎng)成分的物理保鮮方法受到消費(fèi)者的排斥,近年來(lái),從植物中提取天然物質(zhì)用于保障食品的質(zhì)量和安全成為了研究趨勢(shì)。目前,隨著對(duì)細(xì)菌群體感應(yīng)(quorum-sensing,QS)系統(tǒng)研究的不斷深入,研究人員也逐漸關(guān)注到細(xì)菌QS在水產(chǎn)品腐敗中發(fā)揮的作用,對(duì)其進(jìn)行了大量的研究報(bào)道,試圖闡明QS信號(hào)分子在水產(chǎn)品腐敗中的作用機(jī)制,從分子水平上說(shuō)明水產(chǎn)品腐敗的原因,并以此為突破口,從致腐菌株QS角度抑制或減緩水產(chǎn)品腐敗進(jìn)程,為水產(chǎn)保鮮提供新路徑。綜上,有必要總結(jié)闡述致腐細(xì)菌群體感應(yīng)信號(hào)系統(tǒng)調(diào)控水產(chǎn)品腐敗變質(zhì)的研究進(jìn)展,以期對(duì)水產(chǎn)品保鮮工作提供必要研究信息。
1 細(xì)菌群體感應(yīng)現(xiàn)象
細(xì)菌是高度密集的生物群體,QS是一種廣泛存在于細(xì)菌之間的交流機(jī)制(圖1),這種通訊機(jī)制可以調(diào)節(jié)細(xì)菌的一系列重要行為[12],它能使基因的表達(dá)與群體細(xì)胞密度同步[13],以此來(lái)調(diào)節(jié)細(xì)菌在環(huán)境中的適應(yīng)能力,已知的受QS調(diào)控的行為有:生物發(fā)光[14]、質(zhì)粒接合轉(zhuǎn)移[15-16]、毒性基因表達(dá)[17]、色素合成[18-19]、細(xì)菌群游[20]、生物聚集行為[21]等。
QS現(xiàn)象最初發(fā)現(xiàn)于一種名為哈維氏弧菌(Vibrioharveyi)的發(fā)光海洋細(xì)菌中,在研究中發(fā)現(xiàn),哈維氏弧菌可以感受自身產(chǎn)生的信號(hào)分子,也可以感受到其他細(xì)菌產(chǎn)生的信號(hào)分子,當(dāng)信號(hào)分子累積到一定程度,便會(huì)誘導(dǎo)哈維氏弧菌產(chǎn)生發(fā)光現(xiàn)象[22]。QS于1994年首次被TAKAYAMA等[23]提出并定義,研究人員如今將QS概述為:隨著細(xì)菌的生長(zhǎng)繁殖,大多數(shù)細(xì)菌可以產(chǎn)生一種誘導(dǎo)性質(zhì)的信號(hào)分子,自誘導(dǎo)物的濃度隨著細(xì)菌種群密度的增加逐漸增多,當(dāng)這種自誘導(dǎo)物信號(hào)分子累積到一定濃度閾值時(shí),便能夠刺激或誘導(dǎo)相關(guān)基因的表達(dá),從而使細(xì)菌展現(xiàn)出與低密度條件時(shí)截然不同的群體行為以及特性,以此來(lái)應(yīng)對(duì)環(huán)境的變化[24-26]。QS系統(tǒng)基本都是由誘導(dǎo)器、信號(hào)合成酶、信號(hào)受體、信號(hào)相應(yīng)調(diào)節(jié)器和調(diào)控基因五部分組成,細(xì)菌的種類(lèi)不同,自誘導(dǎo)因子也有所區(qū)別(圖2)。

a-革蘭氏陰性菌;b-革蘭氏陽(yáng)性菌
革蘭氏陽(yáng)性菌大多分泌自誘導(dǎo)肽類(lèi)信號(hào)分子(autoinducing peptides,AIPs)作為QS系統(tǒng)之間交流的化學(xué)信號(hào)分子[26],雙組分組氨酸激酶(histidine kinase,HK)作為信號(hào)傳感和轉(zhuǎn)導(dǎo)模塊[27],AIP是一種被修飾的寡肽,不能自由地跨細(xì)胞膜擴(kuò)散,需要由雙組分組氨酸激酶介導(dǎo)轉(zhuǎn)運(yùn)過(guò)程[28]。AIP信號(hào)系統(tǒng)是基于磷酸化/去磷酸化級(jí)聯(lián)反應(yīng)的,磷酸化級(jí)聯(lián)反應(yīng)產(chǎn)生的信號(hào)分子激活DNA結(jié)合蛋白,從而影響特定基因的轉(zhuǎn)錄,細(xì)菌將信號(hào)分子分泌到生存環(huán)境中,隨著細(xì)菌的生長(zhǎng)繁殖,信號(hào)分子在環(huán)境中累積的濃度越來(lái)越高,與信號(hào)識(shí)別系統(tǒng)相互識(shí)別,逐漸達(dá)到啟動(dòng)某個(gè)基因的閾值濃度,從而進(jìn)行相應(yīng)的生理調(diào)控[29-30]。
革蘭氏陰性菌QS系統(tǒng)由自誘導(dǎo)合成酶(例:LuxM和 LuxI)和轉(zhuǎn)錄調(diào)節(jié)因子(例:LuxN和 LuxR)2部分組成[31],產(chǎn)生N-乙酰-L-高絲氨酸內(nèi)酯(N-acyl homoserine lactones,AHLs)類(lèi)自誘導(dǎo)因子[32],AHLs類(lèi)自誘導(dǎo)因子由一個(gè)高絲氨酸內(nèi)酯環(huán)和一個(gè)N-酰基側(cè)鏈構(gòu)成(圖3),因?yàn)镹-酰基側(cè)鏈的碳原子個(gè)數(shù)各不相同[34],所以AHLs類(lèi)自誘導(dǎo)因子種類(lèi)豐富。若細(xì)菌通過(guò)LuxI/LuxR系統(tǒng)調(diào)控相關(guān)基因的表達(dá),先由LuxI同源蛋白合成AHLs信號(hào)分子,通過(guò)自由擴(kuò)散將其從細(xì)胞內(nèi)運(yùn)輸?shù)郊?xì)胞外[35],當(dāng)細(xì)胞外的AHLs信號(hào)分子達(dá)到一定的濃度閾值,會(huì)與受體蛋白LuxR結(jié)合形成復(fù)合物,與靶基因的啟動(dòng)子結(jié)合,刺激一系列基因的表達(dá),產(chǎn)生相應(yīng)的腐敗所需毒力因子[36-37]。

圖3 酰基高絲氨酸內(nèi)酯(AHLs)的化學(xué)分子結(jié)構(gòu)
除了典型的革蘭氏陽(yáng)性菌和革蘭氏陰性菌中存在的QS系統(tǒng)外,由于天然細(xì)菌群落具有細(xì)菌多樣性和空間結(jié)構(gòu)性,有些細(xì)菌可以同時(shí)利用幾個(gè)QS系統(tǒng),通過(guò)截獲其他QS系統(tǒng)產(chǎn)生的信號(hào)分子,也可以進(jìn)行種間QS信息交流,使自己的相關(guān)基因得到表達(dá)[38],這些系統(tǒng)之間也有著錯(cuò)綜復(fù)雜的聯(lián)系,所以,考慮種間QS信號(hào)如何發(fā)揮作用顯得尤為重要。研究發(fā)現(xiàn),革蘭氏陽(yáng)性菌與革蘭氏陰性菌之間能夠利用呋喃硼酸二酯類(lèi)自誘導(dǎo)物-2(autoinducer-2,AI-2)[39]作為信息交流的信號(hào)分子,其產(chǎn)生依賴(lài)于LuxS蛋白[40],信號(hào)分子由4,5 -二羥基- 2,3 -戊二酮(4,5-dihydroxy-2,3-pentanedione,DPD)經(jīng)LuxS酶催化合成,然后被LuxPQ蛋白跨膜轉(zhuǎn)運(yùn)到轉(zhuǎn)錄調(diào)節(jié)因子[31]。研究普遍認(rèn)為,許多革蘭氏陽(yáng)性菌與革蘭氏陰性菌均能夠產(chǎn)生AI-2,這些QS信號(hào)分子是通用的,所以,它們也被稱(chēng)為L(zhǎng)uxS/AI-2型信息系統(tǒng)[24]。此外,一些革蘭氏陰性菌中尤其是海洋細(xì)菌會(huì)產(chǎn)生環(huán)二肽類(lèi)(diketopiperazine,DKPs)自誘導(dǎo)因子,DKPs通過(guò)激活或抑制LuxR型蛋白而調(diào)節(jié)LuxR型QS[41]。銅綠假單胞菌(Pseudomonasaeruginosa)能夠產(chǎn)生喹諾酮類(lèi)信號(hào)分子[42]、擴(kuò)散信號(hào)因子 (diffusible signaling factor,DSF)[43]等,并且刺激胞外多聚物(exo-polysaccharide,EPS)和其他毒力因子的產(chǎn)生[44]。
有的細(xì)菌存在QS現(xiàn)象,但是細(xì)菌自身并不產(chǎn)生引起QS的信號(hào)分子,這些細(xì)菌可以利用其他細(xì)菌分泌的QS信號(hào)分子來(lái)增強(qiáng)自身的集體行為,這種現(xiàn)象被稱(chēng)為竊聽(tīng)現(xiàn)象(圖4)。群體細(xì)菌共同分享信號(hào)分子可以保持群體菌落的穩(wěn)定,有一些細(xì)菌會(huì)因?yàn)楦`聽(tīng)行為充分利用了QS信號(hào)分子而變成菌群中的優(yōu)勢(shì)菌種,波羅的海希瓦氏菌(Shewanellabaltica)作為水產(chǎn)品中常見(jiàn)的致腐菌株,它可以竊聽(tīng)其他細(xì)菌產(chǎn)生的AHLs信號(hào)分子,使自身成為水產(chǎn)品中的優(yōu)勢(shì)致腐菌株,同時(shí)增強(qiáng)自身致腐能力[45]。細(xì)菌的竊聽(tīng)現(xiàn)象不止存在于不同菌種之間,同時(shí)存在于同種屬之間,像腐敗希瓦氏菌和波羅的海希瓦氏菌之間也存在竊聽(tīng)現(xiàn)象[45]。

圖4 細(xì)菌QS系統(tǒng)竊聽(tīng)現(xiàn)象
2 群體感應(yīng)系統(tǒng)與水產(chǎn)品腐敗
2.1 QS對(duì)水產(chǎn)品腐敗指標(biāo)的影響
大多數(shù)水產(chǎn)品的腐敗都是由致腐細(xì)菌的生長(zhǎng)繁殖和代謝活動(dòng)引起的,不同的貯存溫度和加工方式會(huì)導(dǎo)致不同的腐敗菌生長(zhǎng),希瓦氏菌、假單胞菌、肉食桿菌屬、索絲菌屬、發(fā)光桿菌屬、不動(dòng)桿菌、氣單胞菌等均被報(bào)道作為不同水產(chǎn)品中的優(yōu)勢(shì)腐敗菌株[46]。多項(xiàng)研究已證明許多參與水產(chǎn)品腐敗的菌株能夠分泌QS信號(hào)分子,這些信號(hào)分子在水產(chǎn)品腐敗過(guò)程中不斷積累并發(fā)揮形成生物膜、降解蛋白質(zhì)和脂質(zhì)等作用[47]。
ZHU等[48]采用生物報(bào)告實(shí)驗(yàn)、薄層層析和氣相色譜-質(zhì)譜聯(lián)用技術(shù)對(duì)冷藏凡納濱對(duì)蝦腐敗過(guò)程中的QS信號(hào)分子進(jìn)行了檢測(cè),結(jié)果顯示,存在3種類(lèi)型的自誘導(dǎo)因子,AHLs、AI-2和DKPs均可能參與冷藏凡納濱對(duì)蝦的腐敗變質(zhì);研究發(fā)現(xiàn),外源AHLs提高了凡納濱對(duì)蝦中致腐細(xì)菌的生長(zhǎng)速率和種群占比,外源性4,5-二羥基-2,3-戊二酮通過(guò)抑制競(jìng)爭(zhēng)菌的生長(zhǎng)提高了蝦肉特定腐敗菌的占比。GU等[49]從4 ℃貯藏的大黃魚(yú)中分離出102株腐敗菌,其中60株為希瓦氏菌,鑒定為大黃魚(yú)的特異性腐敗菌,從中檢測(cè)出4種DKPs分子,通過(guò)添加這4種合成的DKPs,以TVB-N值作為腐敗潛力的評(píng)價(jià)指標(biāo),結(jié)果表明,4種DKPs均對(duì)希瓦氏菌的致腐能力具有促進(jìn)作用。羅未[50]測(cè)定了貯藏期間鯖魚(yú)組胺呈累計(jì)上升趨勢(shì),并檢測(cè)出一共含有3種DKPs信號(hào)分子,相關(guān)性分析結(jié)果顯示,腐敗指標(biāo)生物胺的積累與DKPs信號(hào)分子變化趨勢(shì)顯著相關(guān)。
此外,QS系統(tǒng)可以調(diào)控水產(chǎn)品致腐菌株致腐基因的表達(dá)。RpoS是QS系統(tǒng)中一種重要的調(diào)節(jié)因子,參與許多病原體毒力因子的表達(dá),LIU等[51]研究發(fā)現(xiàn),RpoS通過(guò)調(diào)節(jié)胞外蛋白酶和滅菌三文魚(yú)汁中TVB-N的產(chǎn)生來(lái)促進(jìn)熒光假單胞菌的腐敗,假單胞菌存在QS現(xiàn)象,其能夠產(chǎn)生AHLs信號(hào)分子,通過(guò)構(gòu)建了RpoS的缺失突變菌株以探索其在熒光假單胞菌中的功能,AHLs的產(chǎn)生和5個(gè) AHLs 相關(guān)基因的轉(zhuǎn)錄水平與野生型菌株相比,RpoS突變體明顯降低,所以QS通過(guò)刺激致腐基因的表達(dá),加速了水產(chǎn)品的腐敗變質(zhì)。WANG等[52]運(yùn)用全基因組測(cè)序分析方法揭示了大菱鲆的特定腐敗菌熒光假單胞菌PF08的QS系統(tǒng)與其致腐性的關(guān)系,證明了QS系統(tǒng)誘導(dǎo)蛋白酶、生物膜、硫及生物胺的生成等致腐相關(guān)基因表達(dá),闡明了QS系統(tǒng)在調(diào)控相關(guān)腐敗基因時(shí)發(fā)揮的關(guān)鍵作用。
2.2 QS對(duì)生物膜生成的影響
水產(chǎn)品的腐敗與致腐細(xì)菌生物膜的生成密不可分。生物膜通常被定義為附著在物體表面并包被在胞外聚合物基質(zhì)中的高密度細(xì)菌團(tuán)[53],生物膜細(xì)胞相對(duì)于單個(gè)細(xì)胞具有其獨(dú)特的性質(zhì),包括對(duì)抗菌藥物的耐受性增強(qiáng)、對(duì)外界環(huán)境的附著能力增強(qiáng)等,使得其難以從食品所處環(huán)境中去除[8]。由于生物膜的頑強(qiáng)性,導(dǎo)致其總是成為食源性疾病和食品腐敗變質(zhì)的潛在關(guān)鍵因素,生物膜通常含有高濃度的細(xì)胞,是QS細(xì)胞密度依賴(lài)的基因表達(dá)調(diào)控的重要組成部分[54-55]。細(xì)菌的QS現(xiàn)象有助于生物膜的形成、流動(dòng)和物質(zhì)的分泌以及提高生物膜抗菌治療的能力[56-58]。AHLs影響食品腐敗細(xì)菌和食源性致病菌對(duì)細(xì)胞外聚合物的初始附著和產(chǎn)生,是對(duì)生物膜形成影響最多的QS系統(tǒng)[59]。PARSEK等[13]研究表明,QS通過(guò)減少生物膜的主要胞外多糖之一的合成和誘導(dǎo)表面活性劑分子(鼠李糖脂)的合成,來(lái)促進(jìn)銅綠假單胞菌生物被膜的分散。XIANG等[60]研究發(fā)現(xiàn),熒光假單胞菌的QS現(xiàn)象調(diào)控與生物膜密切相關(guān)的胞外凝集素的產(chǎn)生,影響其基因表達(dá),c-di-GMP是該水產(chǎn)品致腐菌株生物膜形成的關(guān)鍵調(diào)控因子,QS促進(jìn)了該基因的表達(dá),另外,由生物被膜表面蛋白表達(dá)的轉(zhuǎn)錄譜分析發(fā)現(xiàn),大多數(shù)基因受到QS的調(diào)控。
2.3 QS相關(guān)基因?qū)λa(chǎn)品腐敗的影響
一個(gè)完整的QS系統(tǒng)離不開(kāi)相關(guān)信號(hào)受體,為了通過(guò)抑制QS現(xiàn)象來(lái)抑制水產(chǎn)品的腐敗,研究人員構(gòu)建了相關(guān)接收QS信號(hào)的基因缺陷型菌株,將控制分泌QS信號(hào)受體的基因敲除,通過(guò)造成信號(hào)分子受體缺陷而無(wú)法被識(shí)別,進(jìn)而使得QS系統(tǒng)的表達(dá)受阻。曾名湧教授團(tuán)隊(duì)通過(guò)同源重組技術(shù)構(gòu)建了希瓦氏菌LuxR受體的缺陷菌株,希瓦氏菌LuxR基因敲除后,與野生型希瓦氏菌相比較,其生長(zhǎng)沒(méi)有受到明顯影響,但是其致腐、產(chǎn)硫、QS竊聽(tīng)和感受外源信號(hào)分子的能力顯著降低,減緩了凡納濱對(duì)蝦的腐敗[61-62];朱耀磊等[63]用基因敲除構(gòu)建了QS基因LuxRI缺失型菌株,與野生型相比,即食海參的蜂房哈夫尼菌H4(HafniaalveiH4)失去分泌AHLs的能力,其生物膜的形成和泳動(dòng)能力也明顯減弱,研究結(jié)果表明QS相關(guān)基因?qū)λa(chǎn)品腐敗具有重要調(diào)控作用。
3 通過(guò)調(diào)控QS抑制水產(chǎn)品腐敗
如圖5所示,理論上的抑制QS途徑包括抑制信號(hào)分子的生成、降解信號(hào)分子、阻止信號(hào)分子擴(kuò)散、產(chǎn)生信號(hào)分子受體的競(jìng)爭(zhēng)性受體以及產(chǎn)生與信號(hào)分子類(lèi)似的信號(hào)分子去競(jìng)爭(zhēng)受體等,可以達(dá)到上述目的化學(xué)物質(zhì)被稱(chēng)為QS抑制劑(quorum-sensing inhibition,QSI)。QSI被認(rèn)為是阻止腐敗菌造成水產(chǎn)品腐敗的有效替代手段,它能夠阻斷細(xì)胞間QS信號(hào)分子交流并抑制特定基因表達(dá)[56],通過(guò)阻斷信號(hào)分子的產(chǎn)生、傳播和與受體結(jié)合等來(lái)達(dá)到抑制腐敗的產(chǎn)生[65]。

圖5 抑制細(xì)菌QS現(xiàn)象的途徑
3.1 植物提取物
研究發(fā)現(xiàn),一些植物提取物具有延緩食品腐敗、防止生物膜形成、降解生物膜等[27,66]功能,具有成為QSI的潛力(表1),研究報(bào)道,其中發(fā)揮主要抑制作用的功能成分包括酚類(lèi)、醌類(lèi)、黃酮類(lèi)、生物堿類(lèi)、萜類(lèi)、聚乙炔類(lèi)等[75-76]。檸檬油能夠干擾AHLs介導(dǎo)的銅綠假單胞菌和嗜水氣單胞菌(Aeromonashydrophila)QS毒力因子的產(chǎn)生,影響其生物膜的形成[67],從而減緩了致腐細(xì)菌的群聚和生長(zhǎng),能夠延長(zhǎng)水產(chǎn)品的貨架期;LI等[66]研究表明,肉桂醛顯著抑制熒光假單胞菌的QS相關(guān)因子,在不干擾AHLs信號(hào)分子的基礎(chǔ)上,可與LuxR型蛋白結(jié)合,使熒光假單胞菌QS依賴(lài)表型發(fā)生改變,從而影響熒光假單胞菌胞外蛋白酶的產(chǎn)生、游動(dòng)和聚集運(yùn)動(dòng)以及生物膜的形成,并通過(guò)測(cè)定大菱鲆4 ℃貯藏期間TVB-N值的變化,證明了肉桂醛可以抑制大菱鲆貯藏期間TVB-N值的上升,有效抑制了大菱鲆的腐敗進(jìn)程;HUSAIN FOHAD等[69]研究稱(chēng),薄荷油強(qiáng)烈干擾了AHLs調(diào)控的銅綠假單胞菌和嗜水氣單胞菌毒力因子和生物被膜的形成,使得AHLs介導(dǎo)的紫色桿菌素產(chǎn)量呈劑量依賴(lài)性降低,致腐菌株的生長(zhǎng)均不受影響,分子對(duì)接結(jié)果證明薄荷醇具有QSI的特性;ZHU等[68]研究指出,綠茶多酚類(lèi)物質(zhì)可引起大黃魚(yú)中波羅的海希瓦氏菌QS系統(tǒng)AI-2分子的降解,同時(shí)降低了DKPs的活性,并且腐敗指標(biāo)三甲胺的生成也隨之減少;VASAVI等[70]研究了從番石榴中提取的黃酮組分對(duì)銅綠假單胞菌的菌落群體運(yùn)動(dòng)和生物膜的形成的影響,采用紫色桿菌CV026生物傳感器法測(cè)定番石榴葉黃酮的抗QS活性,通過(guò)評(píng)估紫色桿菌素產(chǎn)生的反應(yīng),結(jié)果表明,黃酮組分通過(guò)干擾AHLs信號(hào)的反應(yīng)來(lái)抑制QS,但并不抑制細(xì)菌的生長(zhǎng)和AHLs的合成;HUSAIN FOHAD等[71]的實(shí)驗(yàn)結(jié)果表明,芒果葉的甲醇提取物可以使得AHLs信號(hào)分子含量顯著降低,細(xì)菌的生長(zhǎng)卻不受影響,掃描電鏡結(jié)果顯示,還破壞了銅綠假單胞菌胞外多糖的合成,從而影響其生物膜的形成,除了生物膜的生成受到影響外,芒果葉的甲醇提取物對(duì)受QS調(diào)控的多個(gè)性狀均表現(xiàn)出了抑制作用,表明其通過(guò)QS干擾可能具有多靶點(diǎn)的廣譜活性;從柚子皮中篩選到的QS抑制活性物質(zhì)鑒定為柚皮苷類(lèi)化合物,計(jì)算機(jī)模擬顯示,柚皮苷類(lèi)化合物與QS受體LasR/LuxR結(jié)合穩(wěn)定,使得信號(hào)分子無(wú)法與受體結(jié)合,從而達(dá)到了抑制細(xì)菌的QS現(xiàn)象的目的[67]。
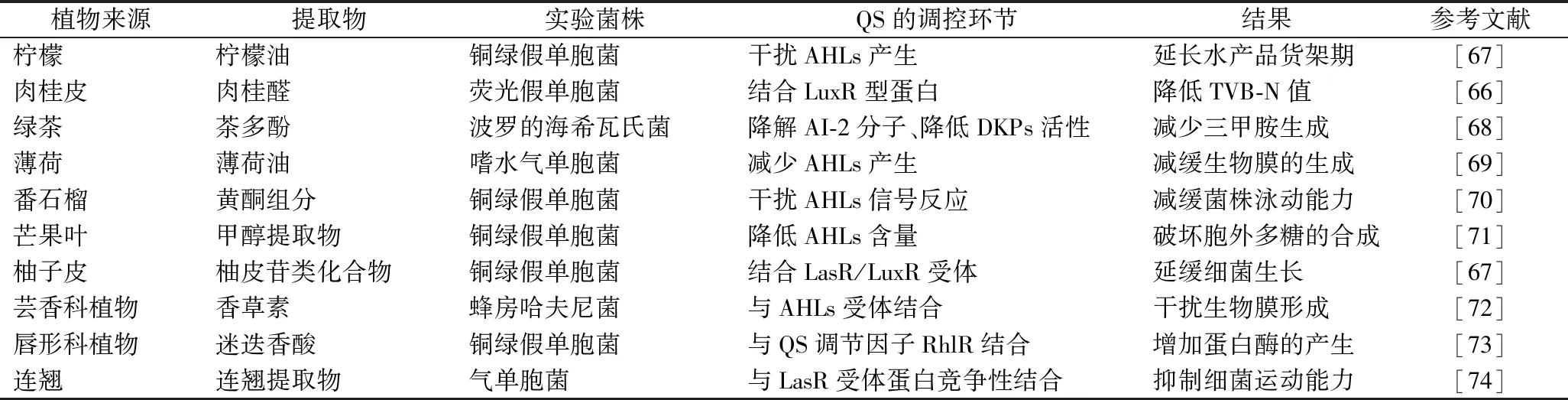
表1 植物源提取物調(diào)控QS及其抑制水產(chǎn)品腐敗作用
3.2 植物提取物
群體淬滅是指通過(guò)使用生物或者非生物因素降解信號(hào)分子從而干擾QS,阻斷細(xì)胞間的通訊交流,信號(hào)降解可以通過(guò)改變菌落生存環(huán)境的化學(xué)特性來(lái)實(shí)現(xiàn),如改變其pH;也可以是來(lái)自細(xì)菌或動(dòng)物產(chǎn)生的酶的作用,已知AHLs降解酶的2種主要類(lèi)型是乳糖酶和酰化酶[77],乳糖酶水解AHLs的HSL環(huán)進(jìn)而生成相應(yīng)的酰基胡桃苷,酰化酶切割A(yù)HLs的酰胺鍵生成相應(yīng)的脂肪酸和高絲氨酸內(nèi)酯[78]。JIE等[61]文章顯示,來(lái)自芽孢桿菌屬(Bacillussp.)新型高絲氨酸內(nèi)酯酶(AHLs內(nèi)酯酶)通過(guò)水解AHLs的內(nèi)酯鍵來(lái)降低AHLs的活性,淬滅了QS信號(hào)分子,顯著降低了水產(chǎn)品優(yōu)勢(shì)腐敗菌的致腐能力。
4 結(jié)論與展望
伴隨人們?nèi)找嬖鲩L(zhǎng)的水產(chǎn)品攝入需求,減少水產(chǎn)品腐敗變質(zhì)成為水產(chǎn)品加工領(lǐng)域重點(diǎn)研究方向之一。本文從致腐細(xì)菌引發(fā)水產(chǎn)品中腐敗變質(zhì)的角度出發(fā),詳細(xì)介紹了菌株QS系統(tǒng)的分類(lèi)及其相關(guān)的作用機(jī)制,闡述了已開(kāi)展的研究中如何從細(xì)菌QS環(huán)節(jié)抑制水產(chǎn)品腐敗現(xiàn)象,并對(duì)相關(guān)QSI使用情況和作用效果的研究進(jìn)展進(jìn)行了總結(jié)。
從QS角度出發(fā)研究細(xì)菌引發(fā)的水產(chǎn)品腐敗變質(zhì)尚處于初步階段,研究人員仍舊需要對(duì)細(xì)菌QS系統(tǒng)及其產(chǎn)生何種QS信號(hào)分子、不同QS信號(hào)分子如何調(diào)控細(xì)菌致腐能力做更深一步的研究;其次,綠色來(lái)源的QSI作為延緩水產(chǎn)品腐敗變質(zhì)、保障水產(chǎn)品品質(zhì)安全的重要新型抑菌保鮮劑,其作用機(jī)理、提取的規(guī)模化制備以及使用穩(wěn)定性等方面都需要開(kāi)展更深一步的研究。目前,對(duì)于水產(chǎn)品致腐菌株QSI的研究仍停留在實(shí)驗(yàn)環(huán)境中,如何將理論實(shí)驗(yàn)結(jié)果運(yùn)用到實(shí)際生產(chǎn)中同樣面臨挑戰(zhàn)。但伴隨系統(tǒng)生物學(xué)、食品組學(xué)、生物信息學(xué)、合成生物學(xué)等領(lǐng)域的發(fā)展,相信未來(lái)可以更好地闡明基于細(xì)菌QS系統(tǒng)的水產(chǎn)品腐敗變質(zhì)機(jī)制,為水產(chǎn)品品質(zhì)與安全控制提供新的理論基礎(chǔ)與技術(shù)支撐。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國(guó)生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(hào)(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(hào)(2018年2期)2018-04-18 12:18:10
鐵道通信信號(hào)(2016年11期)2016-06-01 12:11:32
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
中國(guó)病理生理雜志(2015年8期)2015-12-21 12:38:06
