環境基質對芽孢桿菌狀態演變與熱抗性影響的研究進展
2023-12-29 08:21:30耿方琳牛洪梅賈凱劉陽泰王翔董慶利
食品與發酵工業 2023年24期
耿方琳,牛洪梅,賈凱,劉陽泰,王翔,董慶利
(上海理工大學 健康科學與工程學院,上海,200093)
芽孢桿菌可形成內生芽孢,芽孢的形成是菌體對環境中營養物質的消耗產生的細胞反應,通常在細胞密度較高、營養匱乏的情況下觸發。營養體出于自我保護的目的發生休眠,形成不具有代謝活力的芽孢,從而獲得了對熱、輻射、干燥、極端pH值及毒性化學物質的高抗性[1]。休眠狀態下的芽孢依然能夠監測周圍環境,并在環境條件適宜時萌發,最終恢復到具有正常代謝能力的營養體狀態,繼續生長。芽孢桿菌屬(Bacillusspp.)常于糧油制品與乳制品中檢出,罐頭制品也是受到芽孢污染的主要食品種類[2]。例如,在乳制品生產中,耐熱芽孢桿菌在熱交換器與濃縮器上附著并形成芽孢[3],能在超高溫滅菌工藝后殘存,并最終進入產品中。可見在食品的生產過程中,芽孢有較大可能抵抗加工工藝中的殺菌處理,并在合適的環境條件中萌發生長,產生腐敗酶或毒素等,導致食品腐敗甚至食源性疾病的發生[4-5]。
自然界中的芽孢桿菌進入食品生產鏈后會面對復雜多樣的環境,所處環境條件與基質成分的不同不僅會在殺菌處理時阻礙或促進殺菌效果,還會對殘存芽孢桿菌的修復與生長產生影響。嗜熱脂肪芽孢桿菌在培養基中的熱抗性較在蒸餾水中明顯更高[6],不同食品中同一菌株對超高壓協同熱處理的失活率也存在差異[7]。因此可能導致芽孢殘存,其在適宜溫度下可以在營養豐富的乳制品中獲得修復并萌發增殖,而在部分pH值較低的食品中,則可能發生進一步的衰亡。
環境基質不僅影響著芽孢桿菌的消長趨勢,還影響著芽孢桿菌菌體的生理狀態演變。芽孢的形成伴隨Spo0A蛋白的磷酸化啟動,至形成具有復雜且織密膜結構的芽孢體結束。此時芽孢內膜的滲透性低、核內水分含量低且吡啶二羧酸(dipicolinic acid,DPA)含量高,使其核內酶與DNA對熱及化學物質具有較強的抗性[8]。而在芽孢萌發時,細胞會發生核內物質泄漏、膜結構通透性增加等一系列變化,營養體恢復代謝與增殖能力,抗性降低。然而,因芽孢形成與萌發時所處基質與環境條件不同,芽孢的形成速度[9]與抗逆性[10],萌發的速度所形成營養體的代謝能力等也都會受到影響。了解不同因素對芽孢桿菌狀態演變的影響,才能更好地根據其抗性的變化在生產中進行控制。
現有工作中對影響芽孢熱殺菌效果的因素研究較多,環境pH和水分活度(water activity,Aw)對不同細菌生理狀態與熱抗性的影響是本領域研究經常考慮的因素,近年來也不乏關于常見食品組分與芽孢桿菌熱失活表現的探討,國內亦有對應的文獻綜述[11];對于芽孢的形成,除環境條件外,二價陽離子對芽孢形成的影響研究較多;而對于芽孢的萌發過程,現有報道更多集中在不同萌發手段的作用效果與作用通路上,少有關于形成條件影響芽孢性質的定量研究。本文整合了現有研究中芽孢在形成、萌發與熱失活3個階段中所處環境基質的不同影響,將環境條件如溫度、pH、Aw與基質中主要的幾種成分(蛋白質、糖類、脂肪、金屬陽離子、磷酸鹽)的作用進行分類討論,旨在補充闡明各環境因素對芽孢桿菌狀態演變及熱抗性的綜合性作用,為芽孢桿菌在食品基質中的進一步研究提供理論參考。
1 環境條件對芽孢桿菌狀態演變及熱抗性的影響
眾所周知,溫度是影響細菌生理狀態的重要因素,但當芽孢桿菌處于同樣的溫度下時,環境pH值與Aw則對芽孢桿菌的狀態演變進程與菌體抗逆性影響顯著。
1.1 溫度
溫度對芽孢的形成與后續抗性都有至關重要的作用。在一定的范圍內,提高芽孢形成溫度可使芽孢核內水分含量降低,從而提高芽孢的熱抗性[12],而在超過一定溫度后,所形成的芽孢的抗性逐漸降低。如在一項針對蠟樣芽孢桿菌的研究中,最適溫度下的成孢速率雖然更快,但芽孢的質量與抗性則不如次優溫度下形成的芽孢[13]。然而,在一項關于不同成孢溫度對熱失活影響的建模研究中[14],成孢溫度雖然影響了熱失活速率,但對菌株熱失活的z值并無顯著影響,因此可以推測成孢溫度的影響與芽孢熱失活的機制無關。
前熱處理與萌發時的溫度對萌發能力也有很大的影響。總體來說,芽孢能夠萌發的溫度范圍與菌株的最適生長溫度范圍近似。對于前熱處理而言,適當的熱激活處理能夠極大地刺激營養因子誘導芽孢萌發的效率[15],但同時一項針對梭菌芽孢的研究表明,在亞致死熱處理后,芽孢萌發生長的條件將受到限制,能夠允許芽孢萌發的pH與NaCl濃度范圍會縮小[16]。
1.2 pH
芽孢形成時的基質pH對芽孢形成速度、芽孢終產量與形成芽孢的熱抗性有著顯著性影響。許多研究表明基質酸度的增加阻礙了芽孢的形成,MTIMET等[17]研究顯示,嗜熱脂肪地芽孢桿菌在pH 6.34的液體培養基中形成的芽孢終產量顯著高于在pH 5.83與pH 7.94中的形成的終產量,通過對比D115℃,發現在pH 5.83~7.10環境下形成芽孢的熱抗性顯著高于在pH 7.95下所形成芽孢的熱抗性。BARIL等[9]的研究與上述結論相符,隨基質pH降低,韋氏芽孢桿菌與地衣芽孢桿菌的芽孢終產量顯著減少,且形成最高熱抗性芽孢的產孢pH近于該菌的最適生長pH。基質酸度的增加使形成芽孢抗性下降的現象在其他研究中得到了佐證,對耐熱芽孢桿菌來說,當基質pH上升時,所形成芽孢對Nisin結合熱處理的抗性顯著提高[18]。成孢pH對芽孢熱抗性的影響很有可能是由酸性環境中的質子置換出了與DPA結合的二價陽離子,使得質子化的蛋白質更容易受熱變性而導致的[19],同時,原生質體再水化、芽孢皮層體積減小等形態變化也促使低pH下芽孢抗性降低[20]。值得注意的是,雖然芽孢形成時的基質pH影響了其對熱及其他處理的抗性,但目前所有研究均表明芽孢的熱敏感性獨立于芽孢形成條件的影響。
基質pH還能夠影響芽孢萌發能力以及萌發后菌體的代謝活力及抗性。對于在食品加工中受到熱處理的芽孢來說,基質pH作用于菌體,并能影響其修復萌發能力。TRUNET等[21]使用雙重染色的方法觀察不同萌發階段的芽孢,發現熱處理后的芽孢在pH 7.4環境中的萌發比例比在pH 5.5中更高,說明酸性條件降低了該菌芽孢的修復萌發能力。類似地,MTIMET等[17]對嗜熱脂肪地芽孢桿菌在不同pH與溫度下的修復進行動力學研究,結果表明最適pH值為7.75,且在最適范圍外,隨pH降低,菌體熱抗性下降。基質中有機酸或高濃度鹽的存在,以協同作用的方式加強了芽孢的熱失活效果,對熱損傷芽孢的生存有顯著影響[22]。對多株芽孢桿菌進行研究發現,若熱處理后的殘余芽孢處于亞致死pH環境中,長期來看,其數量幾乎全部出現明顯下降,但失活速率受到菌株性質與亞致死條件影響[23]。另一項研究也印證了這一點:嗜熱脂肪地芽孢桿菌芽孢在不同pH與溫度組合下進行熱處理并在非最適條件下保存,隨保存時間延長,樣品均發生后續失活,逐漸喪失在瓊脂平板上修復生長的能力,其間在中性條件下進行熱處理的芽孢對不利保存條件的抗性更強[24]。利用流式細胞儀及核酸染色技術,可以發現在酸性條件下進行熱處理后,芽孢中敏感亞群的比例更高,導致后續對不利保存條件的抗性降低[25]。與溫度的作用類似,低pH也對芽孢萌發的其他條件造成了限制,縮小了芽孢桿菌能夠生長的溫度范圍[26]。關于低pH影響芽孢萌發的原因,CIARCIAGLINI等[27]提出,酸性環境可以改變氨基酸側鏈的電離狀態從而改變電荷分布、氫鍵與蛋白質結構,許多蛋白質在pH<5時展開,從而顯著降低了芽孢的萌發能力。
1.3 Aw
所處基質的Aw對芽孢的形成、萌發及其抗逆性均有作用。芽孢形成的時間與終產量受到Aw的影響,對于枯草芽孢桿菌來說,在Aw為0.993的培養條件下,4 d內芽孢濃度即可達到9×109spore/mL,而在Aw0.950下,經17天培養后芽孢濃度也僅有3×107spore/mL[28];韋氏芽孢桿菌與地衣芽孢桿菌芽孢濃度也在培養基質Aw上升至0.996后,出現了多個數量級的增加[9]。而Aw對芽孢萌發的作用則體現在,造成Aw變化的保濕劑分子與芽孢內膜間的作用能夠阻礙芽孢核內DPA的泄漏,從而使其對激活萌發與失活的熱與高壓處理具有了更高的抗性,抑制了此情況下的萌發[29]。SAMAPUNDO[30]測定了芽孢在不同Aw、pH與溫度組合下的失活與萌發生長情況,證明當Aw降低時,熱激活誘導的芽孢萌發延滯期總體增長,所需萌發溫度要求變高。降低Aw也阻礙了芽孢的熱失活效果,當Aw由0.99降至0.92時,熱與高壓協同誘導的蠟樣芽孢桿菌芽孢萌發與失活數量均降低幾個數量級[31];LEGUERIAL等[32]報道了使用蔗糖作為保濕劑降低Aw時,隨蔗糖含量增加,蠟樣芽孢桿菌芽孢熱抗性提高,相似的結果在對嗜熱脂肪地芽孢桿菌、解淀粉芽孢桿菌,以及芽孢桿菌營養體的研究中也被證明;在生物膜中的芽孢熱抗性往往也高于浮游芽孢[33],且濕法培養的生物膜中芽孢雖生成速率不如在干法生物膜中快,但由于培養環境Aw更高,其熱抗性也更強[34]。而對于射頻加熱處理,由于Aw越高介電損耗系數就越高,樣品所吸收的射頻能量也增多,但溫度的上升與樣品的熱性能也有關,因此隨所處基質Aw的升高,射頻加熱滅菌效果先增后減[35]。
Aw對芽孢桿菌狀態演變及抗性的作用效果還受不同保濕劑間差異的影響。對耐熱芽孢桿菌來說,在添加NaCl的培養基中形成的芽孢,在熱處理后所降低的數量更少,而添加甘油雖然同樣降低了Aw,卻未對熱失活效果造成顯著影響[18];然而,JACOBSEN等[36]的研究結果與上述不同,無論使用甘油、山梨醇還是NaCl作為保濕劑,Aw的變化都不對芽孢的熱抗性造成影響。不同保濕劑對芽孢萌發的抑制效果也有所差異,一項對枯草芽孢桿菌、蠟樣芽孢桿菌與巨大芽孢桿菌的萌發動力學研究[37]發現,幾種保濕劑按對萌發的抑制與對熱失活的阻礙效果由強到弱排序都為蔗糖>海藻糖>甘油,但對巨大芽孢桿菌來說,或由于蔗糖被糖苷酶分解為大量葡萄糖,添加蔗糖反而刺激并加速了其芽孢的萌發。
保濕劑抑制芽孢萌發的作用機理因刺激芽孢萌發的方式不同而存在差異(圖1)[1]。有研究表明,低Aw對營養萌發的影響主要作用于早期萌發啟動(commitment)階段,保濕劑的加入延后了芽孢開始泄漏DPA的時間,其中,蔗糖的加入還增加了此段過程的DPA泄漏量,但甘油卻未引起該現象的發生,這可能是由于芽孢內膜對二者的滲透性不同造成的[38]。通常來說,營養萌發中萌發受體與萌發劑的結合需要水分的參與,而蔗糖的加入使芽孢膜結構對水分的滲透性變弱,使萌發受體不易結合,進而阻礙了芽孢的萌發。而對于外源2,6吡啶二羧酸鈣(Ca2+-DPA)與十二烷胺等萌發劑誘導的非營養萌發來說,當環境中保濕劑濃度較高,則萌發劑的溶解度降低,因此不利于芽孢的萌發。此外,某些萌發因子與保濕劑間的相互作用也會影響萌發進程。高壓對芽孢萌發的誘導作用分為兩種,在相對較低壓下,壓力處理促進營養萌發受體結合,而較高壓下則促進Ca2+-DPA泄漏通道的開啟[39-40]。其中,蔗糖對高壓誘導萌發的抑制作用比甘油顯著,但在較低壓(150 MPa)下其對壓力誘導的營養萌發作用不顯著。
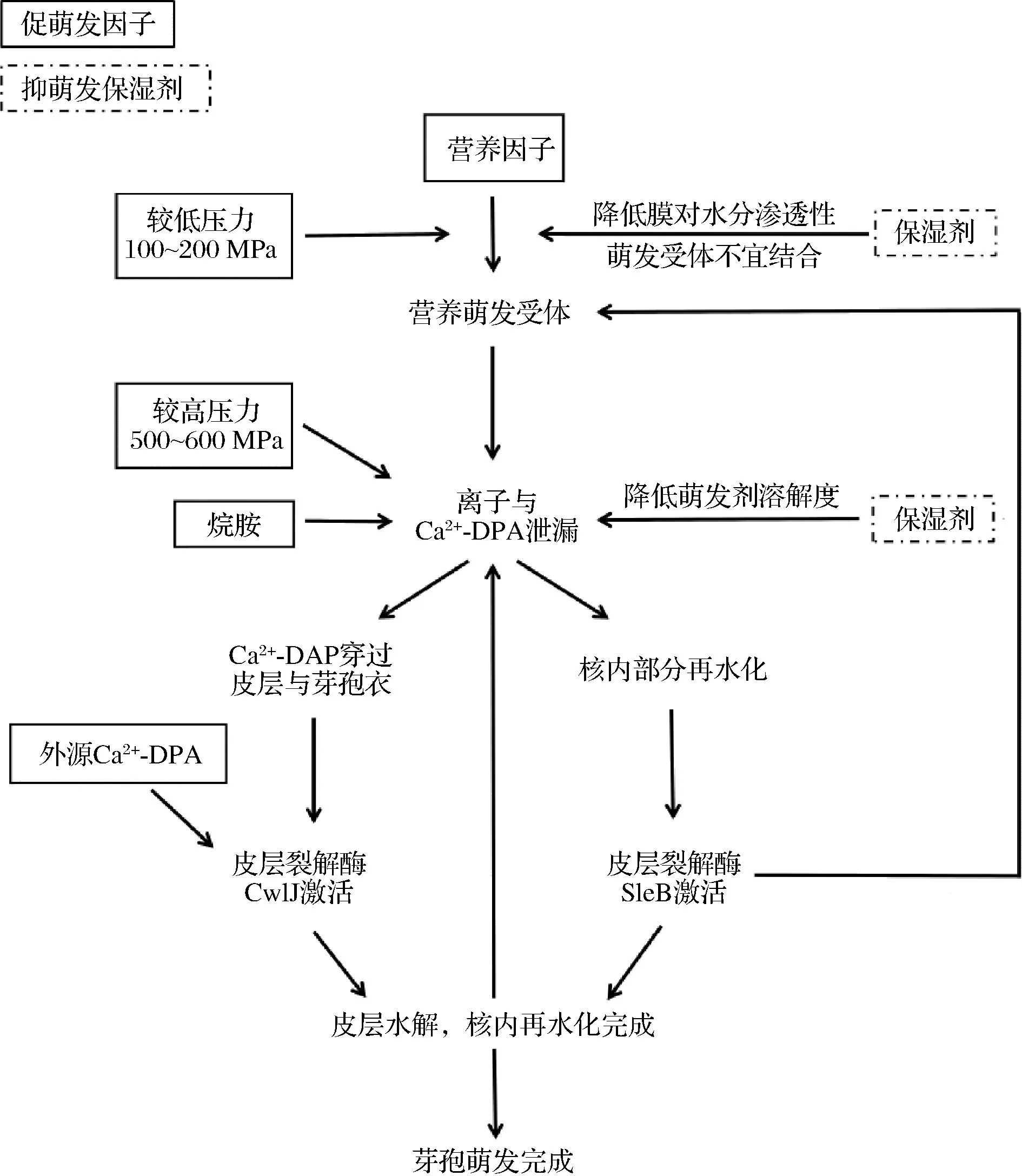
圖1 萌發過程中觸發因素與保濕劑作用示意圖[1]Fig.1 The role of germination trigger factors and humectants during germination
對影響芽孢桿菌生理性質的環境基質進行了總結,如表1所示。

表1 環境基質對芽孢桿菌生理性質的影響Table 1 The effects of ambient matrix on the physiological properties of Bacillus spp.
2 基質成分對芽孢桿菌狀態演變及熱抗性的影響
關于不同基質成分對芽孢桿菌生理狀態的影響,已有研究各有側重。蛋白質、脂肪、糖類等分別對芽孢對熱等因素的抗逆性有影響,而芽孢的形成則在營養匱乏的條件下發生,因此少見基質營養成分對芽孢形成影響的研究。部分二價金屬陽離子能顯著促進芽孢的形成,并增強所形成芽孢的抗性。磷酸鹽的存在能影響芽孢結構的穩定性,從而影響熱失活效果。
2.1 營養成分
基質蛋白質含量影響著芽孢對熱及堿處理的抗性。STOECKEL等[41]等將16株芽孢桿菌產生的芽孢在牛奶與磷酸鹽緩沖液中于不同溫度下進行處理,結果顯示牛奶對芽孢的失活具有阻礙作用。另有研究顯示脫脂牛奶濃縮物與脫脂牛奶間的芽孢熱抗性具有明顯差異,當干物質含量上升至30%時,芽孢在脫脂牛奶濃縮物中的濕熱抗性顯著增加[42]。乳清蛋白對此的作用機理仍需進一步研究,目前可能的推測為蛋白質受到高溫加熱時變性凝固,高濃度蛋白形成凝膠,緊密包裹在芽孢周圍,一方面減緩了熱的傳遞速度,另一方面蛋白質變性后與芽孢壁緊密結合,增加了芽孢壁層的厚度,使其熱抗性進一步增強[43],還有研究表明隨干物質濃度升高,而帶來的Aw降低也可能是芽孢熱抗性增加的原因[44]。基質蛋白質含量對堿處理造成的芽孢失活也有顯著影響,這是因為堿性物質通過溶解芽孢壁上的蛋白質造成菌體結構破壞與失活,但這種使蛋白質變性的行為是不定向的,因此當基質中蛋白質含量增加時,菌體所受到的堿性物質攻擊相對減少,因此抗性有所增加。在WEDEL等[45]的研究中,牛奶殘留物質的存在顯著阻礙了其中六株芽孢桿菌的NaOH失活效果。然而,基質中蛋白質含量的增加對酸造成的芽孢失活并無明顯影響。
糖類與芽孢的萌發和熱抗性有關。其對萌發的作用主要是因為糖類物質可作為碳源被微生物利用,同時部分糖類能夠作為萌發因子誘導營養萌發[46]。而糖類對芽孢熱抗性的影響多是通過改變Aw與細胞內外滲透壓來進行的,已在1.3節中進行敘述,此處不另加討論。
關于脂肪在熱處理時對細菌的保護作用,現有研究存在爭議。部分研究提出,基質中的脂肪能夠降低聯合熱處理對非產芽孢細菌的損傷[47-48],并且,牛奶的脂肪含量越高,其中的蠟樣芽孢桿菌對微波熱處理及紫外殺菌處理的抗性就越強[49]。但除確認電阻加熱中基質脂肪通過改變電導率使殺菌效果下降外,脂肪在熱處理中保護菌體的機制尚不明確。脂肪是否除干擾Aw之外,還通過別的途徑對芽孢起到保護作用,這一問題仍需進行深入研究。
2.2 金屬陽離子
金屬陽離子對芽孢的形成與形成芽孢的熱抗性作用顯著。芽孢的高熱抗性來源于芽孢核內的低水分含量、金屬陽離子與DPA累積導致的礦化作用以及小分子可溶性酸蛋白的高含量,在芽孢的形成過程中,需要環境基質提供Ca2+與核內DPA進行螯合,形成Ca2+-DPA儲存在芽孢核內,以使芽孢核內的生命大分子物質形成穩定凝膠,提高其熱抗性[50],且Ca2+也是芽孢形成中相關蛋白酶與蛋白水解酶重要的輔因子。在田晶晶等[51]的研究中,在凝結芽孢桿菌芽孢形成過程中添加碳酸鈣,發現芽孢終產量顯著增加,且形成芽孢對熱、胃酸及膽鹽的耐受能力均提高,同時碳酸鈣對pH的調整作用還大幅提前了芽孢化啟動的時間。Mn2+等離子作用于芽孢,能夠使芽孢發生礦化作用,從而促進芽孢化進程并提高所產芽孢的熱抗性[52],但Mn2+含量過高也會導致芽孢的熱抗性降低[53]。另有研究表明,在培養基中加入Mg2+使菌體生長的營養體數量增加[54],也在一定程度上使形成的芽孢數量上升。WELLS-BENNIK等[55]研究8種培養基質(分別添加了Ca2+、K+、Mg2+與Mn2+)對18株嗜熱脂肪地芽孢桿菌芽孢形成與熱抗性的影響,發現在添加全部4種離子的瓊脂上形成的芽孢熱抗性最強,早前對枯草芽孢桿菌的研究結果與其一致[56]。這些研究都說明了不同的金屬陽離子對芽孢形成具有促進作用。
2.3 磷酸鹽
關于磷酸鹽對芽孢熱抗性的影響,現有研究結果各不相同。STOECKEL等[41]對比16株芽孢桿菌芽孢在磷酸鹽緩沖液與牛奶中的熱抗性,其中嗜熱脂肪地芽孢桿菌在牛奶中的熱抗性更高;但在其另一項研究中[57],一株蠟樣芽孢桿菌芽孢在磷酸鹽緩沖液與嬰幼兒配方奶粉中對95 ℃熱處理抗性表現相同,與此一致的還有DONG等[58]的研究,解淀粉芽孢桿菌芽孢在磷酸鹽緩沖液、低脂與全脂牛奶中,在110 ℃熱處理下熱抗性無顯著差異。許多研究都提及了磷酸鹽對熱抗性與菌體修復的不利影響,如KUMAR等[59]探究了磷酸鹽對芽孢熱抗性影響的作用機理:在中性條件下,芽孢熱抗性隨基質中磷酸鹽含量的增加而降低,對芽孢失活過程中泄漏的Ca2+進行檢測,以判斷芽孢核內Ca2+-DPA的泄漏程度,發現隨磷酸鹽含量增加,Ca2+泄漏比例上升,說明磷酸鹽是通過螯合并帶離芽孢核內二價陽離子使芽孢熱抗性降低的。
3 結論與展望
本文綜述了部分環境條件與基質成分對芽孢桿菌狀態演變及熱抗性的影響。在菌體面臨的環境條件中,溫度、pH與Aw是最具有代表性的,也是對芽孢形成、萌發以及其熱抗性都具有顯著影響的因素。隨pH與Aw降低,或在不適宜的溫度下,芽孢形成速度減緩、產量降低、抗性變弱,萌發能力也受到影響。在食品基質的常見成分中,蛋白質、糖類與脂肪含量經常被進行組合研究,探究它們對菌體熱失活時的保護作用,蛋白質還能夠在堿性環境下保護菌體。基質中的金屬離子通過加強芽孢核的礦化及合成Ca2+-DPA以促進芽孢的形成,并使產生的芽孢對熱抗性增加;相反的,磷酸鹽則使Ca2+-DPA發生泄漏,降低了芽孢的抗性。
關于環境基質對產芽孢細菌消長的影響,現有研究多集中在其對熱失活的作用,還未系統建立產芽孢細菌在不同食品加工與貯存過程中的數量與狀態連續變化的理解。基于此,本文從以下方面做出展望:其一,需要進一步研究環境基質因素在殺菌處理時對芽孢產生作用的細胞與分子機制,明確各因素在不同殺菌處理方式中是通過何種通路、在何種程度上對芽孢起到保護作用的;其二,應該針對殘存芽孢在不同食品基質及保存條件下的狀態分化,推進針對不同食品的熱殺菌參數及貨架期研究;其三,將芽孢誘導萌發后再殺滅是當前的研究熱點,不同環境基質中芽孢的萌發能力與后續抗性有差異,可以根據這一點進行適當的萌發誘導,更高效地對細菌進行殺滅;其四,建議建立不同培養條件與基質中描述芽孢桿菌狀態及菌數變化的動態模型,以更好地理解其消長趨勢和對食品質量進行控制。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中老年保健(2021年12期)2021-08-24 03:30:40
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
當代陜西(2021年2期)2021-03-29 07:41:24
中國生殖健康(2020年6期)2020-02-01 06:28:50
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年11期)2019-01-07 01:28:02
