淺水湖泊沉積物生源要素與浮游藻類(lèi)和沉水植物的關(guān)系
2024-01-01 00:00:00劉佳豪李艷申?yáng)|方喬瑞婷王洪鑄
水生態(tài)學(xué)雜志 2024年4期

DOI:10.15928/j.1674-3075.202305140129
收稿日期:2023-05-14""" ""修回日期:2023-11-10
基金項(xiàng)目:國(guó)家重點(diǎn)研發(fā)計(jì)劃(2021YFC3200103);中國(guó)科學(xué)院水生生物研究所“生態(tài)文明”特色研究所服務(wù)性項(xiàng)目“阻隔湖泊魚(yú)類(lèi)群落機(jī)構(gòu)調(diào)整及生態(tài)系統(tǒng)功能優(yōu)化研究”(Y85Z0511);武漢市政建設(shè)集團(tuán)有限公司科研項(xiàng)目(wszky202014);湖北省重點(diǎn)研發(fā)計(jì)劃項(xiàng)目 (2022BCA072)。
作者簡(jiǎn)介:劉佳豪,1997年生,男,碩士研究生,主要研究方向?yàn)榈鷳B(tài)學(xué)。E-mail: liujh@ihb.ac.cn
通信作者:李艷,1991年生,女,博士,主要研究方向?yàn)榈鷳B(tài)學(xué)。E-mail: liyan@ihb.ac.cn
摘要:為探究長(zhǎng)江中下游淺水湖泊沉積物碳、氮、磷等生源要素的含量及其對(duì)浮游藻類(lèi)和沉水植物的影響,以長(zhǎng)江中下游11個(gè)淺水湖泊為研究對(duì)象,分析2003-2020年夏季沉積物總氮(TNSed)、總磷(TPSed)和有機(jī)碳(TOCSed)含量與浮游藻類(lèi)和沉水植物的關(guān)系。結(jié)果表明:(1)在長(zhǎng)江中下游湖泊中,TPSed在藻型湖泊中最高,為(0.71±0.21)"g/kg,草型湖泊中最低,為(0.52±0.13")g/kg。TNSed和TOCSed則在草型湖泊中最高,分別為(4.64±3.57)g/kg和(55.89±50.28)g/kg,中間態(tài)湖泊最低,分別為T(mén)NSed(1.86±1.26)g/kg,TOCSed(20.00±12.13)g/kg。(2)回歸分析結(jié)果顯示,夏季浮游藻類(lèi)Chl-a含量在TPSed較高的湖泊中更高(P=0.01),沉水植物生物量(BMac)則在TPSed較低的湖泊中更高(P=0.03),而TNSed和TOCSed與Chl-a和BMac的回歸關(guān)系不顯著(Pgt;0.17)。(3)長(zhǎng)江中下游湖泊夏季沉水植物消失的沉積物總磷閾值約為0.87~0.98"g/kg。在長(zhǎng)江中下游湖泊的沉積物中,磷含量對(duì)浮游藻類(lèi)和沉水植物生物量的預(yù)測(cè)力比碳、氮含量更高,進(jìn)一步研究二者定量關(guān)系后可為評(píng)價(jià)水生態(tài)系統(tǒng)沉積物污染狀況及確定水生植被修復(fù)的沉積物條件需求提供科學(xué)依據(jù)。
關(guān)鍵詞:長(zhǎng)江中下游湖泊;沉積物;生源要素;沉水植物;浮游藻類(lèi)
中圖分類(lèi)號(hào):Q178""""""" 文獻(xiàn)標(biāo)志碼:A""""""" 文章編號(hào):1674-3075(2024)04-0019-09
沉積物作為沉水植物等水生生物生長(zhǎng)的基質(zhì),是湖泊生態(tài)系統(tǒng)中的一個(gè)重要組成部分,對(duì)其健康運(yùn)行有重要作用(H?kanson amp; Jansson,1983)。然而近幾十年來(lái),隨著社會(huì)經(jīng)濟(jì)的發(fā)展,大量以碳、氮、磷元素為主的污染物排入湖泊,并通過(guò)水流和自然沉降等作用進(jìn)入沉積物中。由于長(zhǎng)期的積累,沉積物中碳、氮、磷等生源要素的含量要比上覆水中高幾個(gè)數(shù)量級(jí)(Sondergaard et al,2001;秦伯強(qiáng)等,2005)。因此,沉積物營(yíng)養(yǎng)含量對(duì)水中的物質(zhì)循環(huán)及水生生物的生存與發(fā)展等具有極大的影響。
沉積物作為湖泊水中營(yíng)養(yǎng)的主要來(lái)源之一,其營(yíng)養(yǎng)含量也是體現(xiàn)湖泊健康狀況的重要指標(biāo),但目前尚無(wú)以沉積物營(yíng)養(yǎng)含量作為評(píng)價(jià)指標(biāo)的生態(tài)系統(tǒng)健康評(píng)價(jià)標(biāo)準(zhǔn)。雖然也有學(xué)者使用單因子污染指數(shù)評(píng)價(jià)法、綜合污染指數(shù)法及有機(jī)污染指數(shù)法等評(píng)價(jià)方法對(duì)沉積物污染狀況進(jìn)行分級(jí),但現(xiàn)有評(píng)價(jià)方法主要是通過(guò)以未受污染的沉積物指標(biāo)作為基準(zhǔn)值,并在其基礎(chǔ)上進(jìn)行簡(jiǎn)單分級(jí)(王艷平等,2021;馬金玉等,2021)。上述方法未能體現(xiàn)不同類(lèi)型營(yíng)養(yǎng)元素及其含量對(duì)生態(tài)系統(tǒng)影響的差異,因此其評(píng)價(jià)結(jié)果對(duì)湖泊健康管理及水生態(tài)修復(fù)的具體指導(dǎo)意義仍然不足。在湖泊水生態(tài)修復(fù)中甚至參考土壤污染狀況標(biāo)準(zhǔn)(GB 15618-2018)劃定修復(fù)區(qū)(生態(tài)環(huán)境部和國(guó)家市場(chǎng)監(jiān)督管理總局,2018)。導(dǎo)致這一問(wèn)題的主要原因是沉積物營(yíng)養(yǎng)元素種類(lèi)及含量與湖泊健康狀況及其表征指標(biāo)之間的定量關(guān)系不明確。浮游藻類(lèi)和沉水植物作為湖泊生態(tài)系統(tǒng)的主要初級(jí)生產(chǎn)者,是反映湖泊健康狀況的重要因子(Liu et al,2017)。研究表明,長(zhǎng)江流域淺水湖泊存在多穩(wěn)態(tài)現(xiàn)象,包括以浮游藻類(lèi)占優(yōu)的濁水穩(wěn)態(tài)和以沉水植物占優(yōu)的清水穩(wěn)態(tài)等。驅(qū)動(dòng)淺水湖泊穩(wěn)態(tài)轉(zhuǎn)換的關(guān)鍵因子是水體總磷含量,草-藻穩(wěn)態(tài)轉(zhuǎn)換的總磷閾值為0.08~0.12"mg/L,反向轉(zhuǎn)換的閾值為0.04~0.06"mg/L(Wang et al,2014)。然而,沉積物作為沉水植物生長(zhǎng)的基質(zhì)及水中營(yíng)養(yǎng)的重要來(lái)源,其可能通過(guò)各種生物、化學(xué)過(guò)程與浮游藻類(lèi)和沉水植物相互作用,從而影響湖泊生態(tài)系統(tǒng)的穩(wěn)態(tài)轉(zhuǎn)換。已有研究多基于中小尺度實(shí)驗(yàn)研究沉積物理化特征對(duì)沉水植物等的影響(Wetzel,2001;楚建周等,2006)。在沉積物營(yíng)養(yǎng)含量較低時(shí),沉積物能夠通過(guò)滿(mǎn)足沉水植物生長(zhǎng)所需營(yíng)養(yǎng)直接促進(jìn)沉水植物的生長(zhǎng);而當(dāng)沉積物營(yíng)養(yǎng)含量過(guò)高時(shí),沉積物又能通過(guò)內(nèi)源釋放作用間接促進(jìn)浮游藻類(lèi)的增殖,并對(duì)沉水植物形成遮蔽效應(yīng),抑制沉水植物的生長(zhǎng)。此外,沉積物中有機(jī)質(zhì)含量過(guò)高時(shí)易形成底層缺氧狀態(tài),產(chǎn)生NO2-、S2-等有毒物質(zhì),對(duì)沉積物根系造成毒害作用(Barko amp; Smart,1978;Van Wijck et al,1992)。然而,目前自然湖泊中沉積物營(yíng)養(yǎng)含量與浮游藻類(lèi)和沉水植物的定量關(guān)系尚不明確。
長(zhǎng)江中下游是我國(guó)淺水湖泊分布最為集中的地區(qū),分布著數(shù)千個(gè)淺水湖泊,水域面積占全國(guó)湖泊的20%以上(王洪鑄等,2015)。近幾十年來(lái),隨著社會(huì)經(jīng)濟(jì)的發(fā)展,大量生活廢水和工業(yè)廢水排入這些湖泊,湖泊富營(yíng)養(yǎng)化問(wèn)題嚴(yán)重,生物多樣性下降,水生動(dòng)植物資源衰退(袁賽波等,2019)。因此,本研究選擇長(zhǎng)江中下游11個(gè)淺水湖泊為研究對(duì)象,比較分析沉積物碳、氮、磷等生源要素的含量對(duì)浮游藻類(lèi)和沉水植物的影響,為后期確定湖泊生態(tài)系統(tǒng)健康及水生態(tài)修復(fù)等所需的沉積物條件提供科學(xué)依據(jù)。
1"" 材料與方法
1.1"" 研究區(qū)域概況
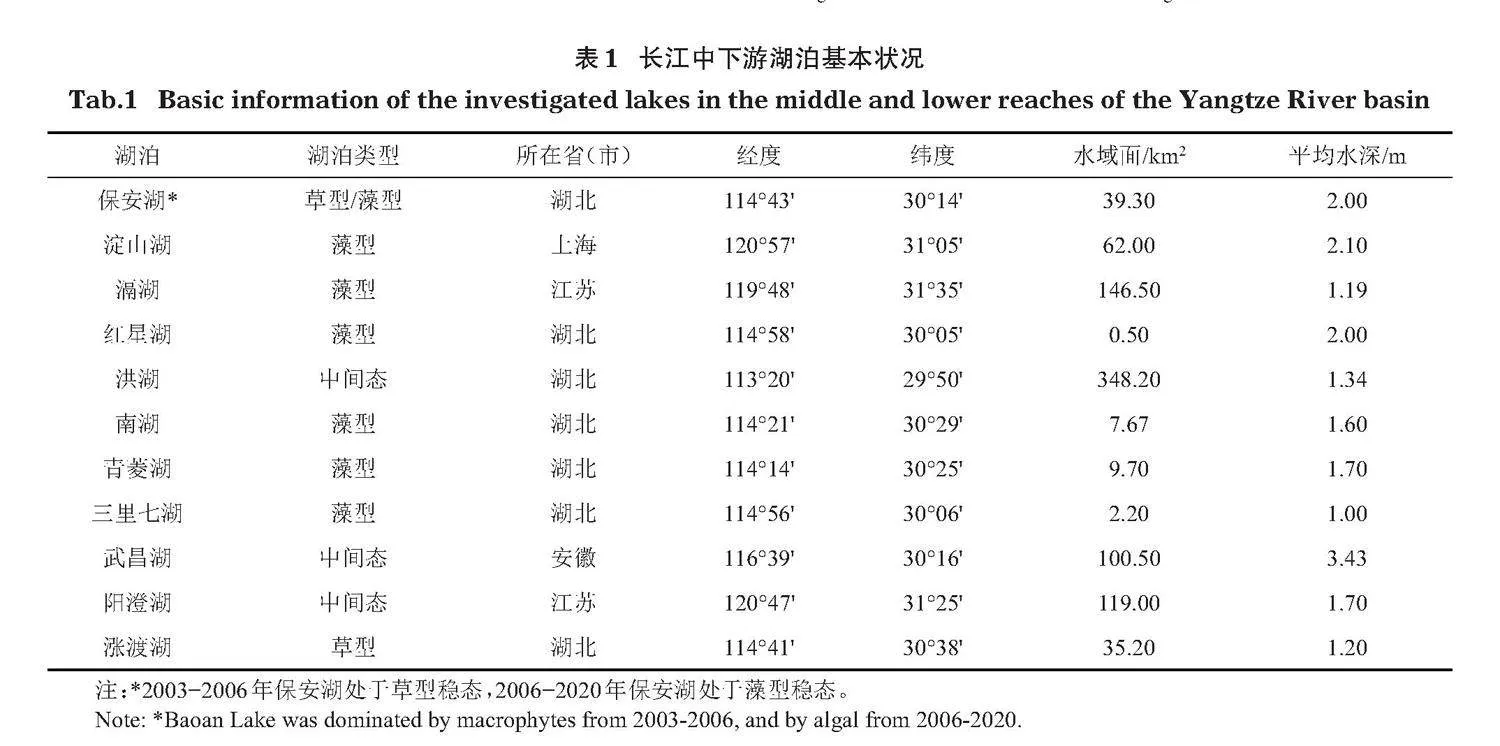
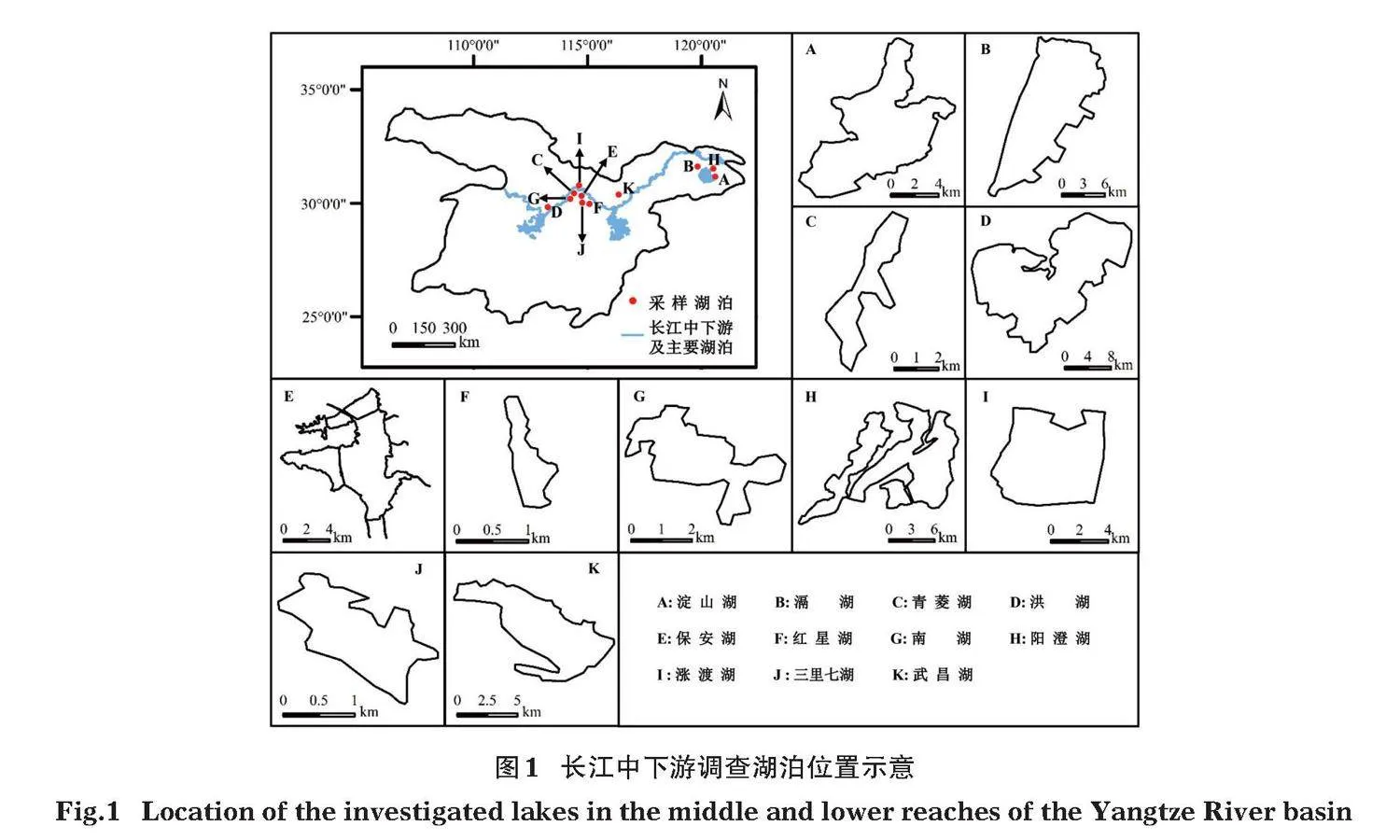
本研究以長(zhǎng)江中下游11個(gè)淺水湖泊(保安湖,淀山湖、滆湖、紅星湖、洪湖、南湖、青菱湖、三里七湖、武昌湖、陽(yáng)澄湖和漲渡湖)為研究對(duì)象(圖1),獲得2003-2020年的調(diào)查數(shù)據(jù)。湖泊基本情況見(jiàn)表1。
按照Bachmann等(2002)的方法,根據(jù)水草和浮游植物干重生物量的比值(BMac/BAlg)將湖泊劃分為3種類(lèi)型:藻型湖泊(BMac/BAlglt;1)、中間態(tài)湖泊(1lt;BMac/BAlglt;100)和草型湖泊(BMac/BAlggt;100)。
1.2"" 樣品采集與分析
1.2.1"" 樣品采集"" 保安湖野外調(diào)查于2003-2012年(除2005年和2008年外)和2020年夏季進(jìn)行,采樣點(diǎn)4~26個(gè)。紅星湖、洪湖、南湖、青菱湖、三里七湖和漲渡湖野外調(diào)查于2003年夏季進(jìn)行,單個(gè)湖泊采樣點(diǎn)3~8個(gè)。淀山湖、滆湖、武昌湖和陽(yáng)澄湖野外調(diào)查于2004年夏季進(jìn)行,單個(gè)湖泊采樣點(diǎn)6~8個(gè)。本研究中相關(guān)數(shù)據(jù)來(lái)源于本團(tuán)隊(duì)歷史調(diào)查數(shù)據(jù)。
用有機(jī)玻璃采水器采集表層(水深0.5"m處)、中層(水深1"m處)、底層水樣(沉積物以上0.1"m處)后混合,裝在1"L的聚乙烯塑料瓶中。沉積物的采集使用彼得森采泥器(UWITEC,A-5310,奧地利)或重力式柱狀采泥器(KC-Denmark,Kajak,丹麥),采集沉積物表層(0~10"cm)的底泥,裝入聚乙烯自封袋中,帶回實(shí)驗(yàn)室。沉水植物使用鐮刀型采草器采集,清洗干凈表面泥沙及底棲動(dòng)物后,吸干表面水分,使用電子秤(SHIMADZU,ELB12K,日本,0.001"g)稱(chēng)量鮮重。沉水植物干重生物量由鮮重生物量乘以0.08獲得(陳洪達(dá)和何楚華,1975)。
1.2.2"" 樣品分析"" 水樣的測(cè)定參照《水和廢水監(jiān)測(cè)分析方法》(國(guó)家環(huán)境保護(hù)總局《水和廢水監(jiān)測(cè)分析方法》編委會(huì),2002)。其中總磷的測(cè)定使用鉬酸銨分光光度法(普析,TU-1810,北京),浮游藻類(lèi)葉綠素a(Chl-a)使用丙酮溶液萃取20~24"h后,在分光光度計(jì)(普析,TU-1810,北京)中讀取665和750"nm處的吸光度值后計(jì)算而來(lái)(黃祥飛等,1999)。浮游藻類(lèi)干重生物量則可由浮游藻類(lèi)葉綠素a含量乘以70獲得(Scheffer,1998)。
沉積物經(jīng)自然風(fēng)干、研磨、過(guò)100目(孔徑0.15"mm)篩后進(jìn)行分析。沉積物的測(cè)定參照《土壤農(nóng)業(yè)化學(xué)分析方法》(魯如坤,1999)。總氮的測(cè)定使用凱氏定氮法(Buchi,K-355,瑞士),總磷的測(cè)定使用高氯酸-硫酸酸溶-鉬銻抗比色法(普析,TU-1810,北京),有機(jī)碳的測(cè)定使用水合熱重鉻酸鉀氧化-比色法(普析,TU-1810,北京)。
1.3"" 數(shù)據(jù)處理
使用Excel 2013、Origin 2022和R 4.2.2對(duì)數(shù)據(jù)進(jìn)行處理與分析,通過(guò)一元線性回歸分析的方法來(lái)分析沉積物總氮、總磷和有機(jī)碳含量與浮游藻類(lèi)和沉水植物的關(guān)系。
2"" 結(jié)果與分析
2.1"" 上覆水和沉積物生源要素含量
上覆水TP含量在藻型湖泊中最高,為(0.18±0.12)mg/L;中間態(tài)湖泊中次之,為(0.06±0.03)mg/L;草型湖泊中最低,為(0.05±0.01)mg/L(圖2)。沉積物數(shù)據(jù)顯示,TNSed和TOCSed含量均表現(xiàn)為草型湖泊最高,分別為(4.64±3.57)和(55.89±50.28)g/kg;藻型湖泊次之,為(3.89±1.66)和(40.11±20.76)g/kg;中間態(tài)湖泊最低(TNSed:lt;1.86±1.26gt;"g/kg;TOCSed:lt;20.00±12.13gt;"g/kg)。TPSed則表現(xiàn)為藻型湖泊最高,為(0.71±0.21)"g/kg,中間態(tài)湖泊次之,(0.59±0.08)"g/kg,草型湖泊最低,為(0.52±0.13)"g/kg(圖2)。
2.2"" 浮游藻類(lèi)和沉水植物狀況
長(zhǎng)江中下游湖泊中,Chl-a含量在藻型湖泊中最高,為(58.92±50.17)μg/L;中間態(tài)湖泊中次之, 為(16.35±8.91)μg/L;草型湖泊中最低,為(5.25±1.57)μg/L(圖3)。BMac則在草型湖泊中最高,為(2"398.53±2 339.05)g/m2,中間態(tài)湖泊次之,為(370.14±387.11)g/m2,藻型湖泊最低(0"g/m2)(圖3)。
2.3"" 碳氮磷含量與葉綠素a和沉水植物生物量的關(guān)系
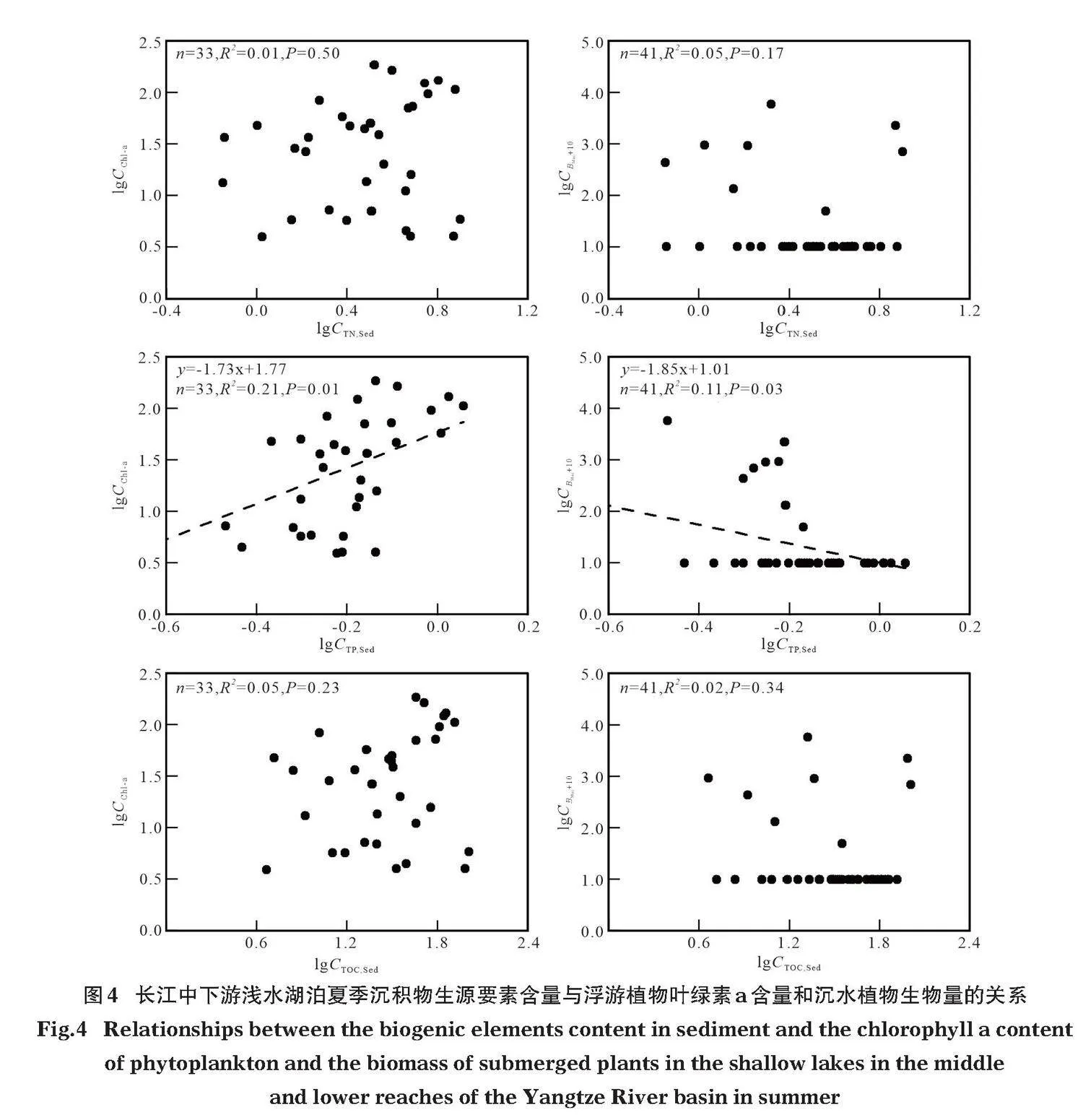
長(zhǎng)江中下游湖泊夏季沉積物總氮、總磷、有機(jī)碳含量與浮游藻類(lèi)葉綠素a含量和沉水植物生物量的回歸分析結(jié)果顯示,浮游藻類(lèi)葉綠素a含量在沉積物總磷含量較高的湖泊中更高(P=0.01),而與總氮和有機(jī)碳的回歸關(guān)系不顯著(Pgt;0.23)(圖4)。沉水植物生物量與沉積物營(yíng)養(yǎng)含量的關(guān)系與浮游藻類(lèi)類(lèi)似,但其生物量在總磷含量較低的湖泊中更高(P=0.03),與總氮和有機(jī)碳的回歸關(guān)系不顯著(Pgt;0.17)(圖4)。
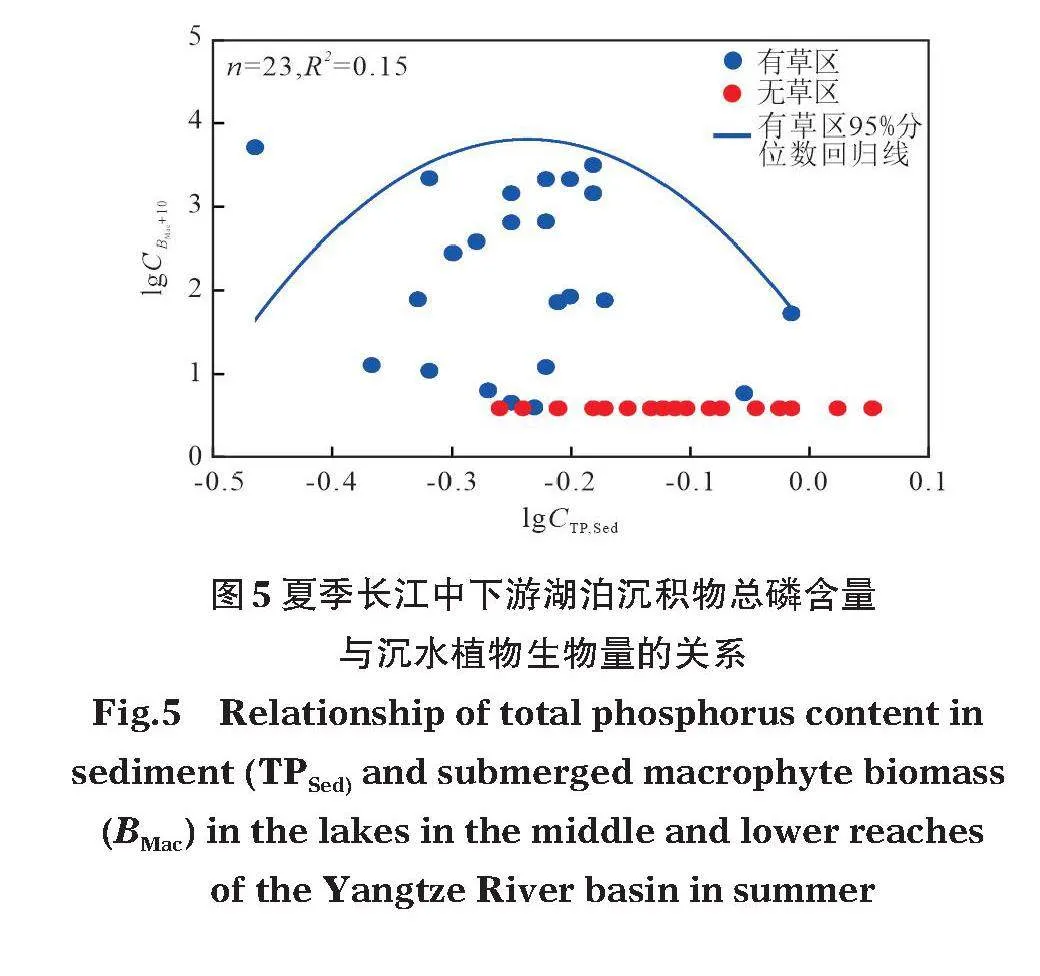
2003-2020年夏季長(zhǎng)江中下游淺水湖泊有草區(qū)和無(wú)草區(qū)沉水植物生物量隨沉積物總磷含量變化的分布圖顯示,有草區(qū)沉積物總磷含量為(0.59±0.14)"g/kg,無(wú)草區(qū)沉積物總磷含量為(0.71±0.20)"g/kg(圖5)。以有草區(qū)沉水植物分布的沉積物總磷含量范圍的95%~100%分位點(diǎn)作為沉水植物消失的沉積物總磷閾值,得到夏季長(zhǎng)江中下游淺水湖泊沉水植物消失的沉積物總磷閾值約為0.87~0.98"g/kg。
3"" 討論
3.1"" 沉積物生源要素與浮游藻類(lèi)和沉水植物的關(guān)系
本次研究表明,相比沉積物氮和碳含量,磷含量對(duì)浮游藻類(lèi)和沉水植物生物量的預(yù)測(cè)力更高。浮游藻類(lèi)葉綠素a含量和沉水植物生物量均與沉積物總磷含量有顯著的回歸關(guān)系,葉綠素a含量在總磷較高的湖泊中更高(P=0.01),沉水植物生物量在總磷較低的湖泊中更高(P=0.03),但二者與總氮和有機(jī)碳含量間的回歸關(guān)系均不顯著(Pgt;0.17)(圖4)。以上結(jié)果表明浮游藻類(lèi)和沉水植物與沉積物磷含量的關(guān)系更密切。
沉積物中碳含量對(duì)浮游藻類(lèi)的生長(zhǎng)沒(méi)有明顯的直接作用,因?yàn)樽匀缓粗袠O少出現(xiàn)碳限制,但可能影響沉水植物的生長(zhǎng)。影響沉水植物生長(zhǎng)的因素包括光照強(qiáng)度、水體氮磷含量、溶氧等(鄢文皓等,2020),而沉積物中有機(jī)碳含量過(guò)高可能通過(guò)分解作用消耗大量氧氣,導(dǎo)致沉積物氧氣含量下降。研究表明,適度的缺氧有利于刺激沉水植物的生長(zhǎng)(Wu et al,2015),但當(dāng)沉積物處于厭氧狀態(tài)時(shí),植物體內(nèi)可能積累乙醇等有毒物質(zhì)。雖然植物自身可通過(guò)薄壁組織的程序性死亡和通氣組織的分化,增加向根部的氧氣輸送量以維持根部的氧化狀態(tài)、使其脫毒,但持續(xù)缺氧可能導(dǎo)致植物爛根、死亡(Brix et al,1987;王文泉和張福鎖,2001;Kozela amp; Regan,2003)。此外,低氧可導(dǎo)致沉積物容重降低,泥-水界面不穩(wěn)定,不利于沉水植被的定植(楊波等,2012)。雖然溶氧等因素能夠影響植物等生存和發(fā)展,但就淺水湖泊而言,由于湖泊水深較淺,風(fēng)力等擾動(dòng)使水體表層與底層之間的交換較為充分,較少出現(xiàn)底層長(zhǎng)時(shí)間缺氧現(xiàn)象。如我們針對(duì)保安湖的長(zhǎng)期監(jiān)測(cè)結(jié)果顯示,其泥-水界面溶氧大多gt;4"mg/L,尚未達(dá)到對(duì)植物產(chǎn)生明顯危害的閾值。因此,沉積物有機(jī)碳含量對(duì)沉水植物沒(méi)有直接影響,難以準(zhǔn)確預(yù)測(cè)沉水植物的生物量。
氮是影響浮游藻類(lèi)和沉水植物的生長(zhǎng)繁殖的影響因子之一,但由于氮循環(huán)在湖泊生態(tài)系統(tǒng)中是開(kāi)放式循環(huán),沉積物或水中的氮含量難以成為浮游藻類(lèi)和沉水植物生長(zhǎng)的最關(guān)鍵因子。當(dāng)湖泊中氮含量較低時(shí),沉積物和水中許多藻類(lèi)和其他微生物能夠通過(guò)生物固氮作用將空氣中的氮?dú)廪D(zhuǎn)化為氨氮從而獲取氮源,而當(dāng)湖泊中氮的含量過(guò)高時(shí),還可能通過(guò)反硝化等作用離開(kāi)(曾巾等,2007)。Schindler等(2008)進(jìn)行了氮磷添加實(shí)驗(yàn),結(jié)果表明控氮不能控制浮游藻類(lèi)的生長(zhǎng)繁殖,還會(huì)促進(jìn)固氮藍(lán)藻的生長(zhǎng)繁殖。李艷等(2021)在長(zhǎng)江中下游4個(gè)池塘中進(jìn)行的氮磷添加實(shí)驗(yàn),也呈現(xiàn)出類(lèi)似的結(jié)果。因此,沉積物中的氮很少直接影響浮游藻類(lèi)的生長(zhǎng),對(duì)浮游藻類(lèi)生物量的預(yù)測(cè)能力較弱。就沉水植物而言,在一定范圍內(nèi)沉積物氮含量的增加有助于促進(jìn)植物的生長(zhǎng),但當(dāng)?shù)窟^(guò)高時(shí),可能會(huì)對(duì)植物的生長(zhǎng)造成不利影響。水體中氮含量,尤其是氨氮增加時(shí)能夠?qū)е鲁了参矬w內(nèi)游離氨基酸積累等生理脅迫,植物自身能夠通過(guò)調(diào)整代謝活動(dòng)進(jìn)行解毒,但解毒過(guò)程消耗大量碳水化合物,因此不利于其生物量的積累。然而有研究表明,水中氨氮含量小于5"mg/L時(shí)對(duì)植物生長(zhǎng)的影響較小(Cao et al,2004;Yu et al,2015),這一閾值高于許多長(zhǎng)江中下游湖泊中氮水平。以上分析表明,沉積物中氮含量雖然能夠在一定程度上影響浮游藻類(lèi)和沉水植物的生長(zhǎng),但卻難以定量預(yù)測(cè)其生物量。
沉積物磷主要通過(guò)為生物提供營(yíng)養(yǎng)元素影響其生長(zhǎng),在一定范圍內(nèi),沉積物總磷含量的升高能夠促進(jìn)浮游藻類(lèi)和沉水植物生物量的增加(Carr,1998),而沉積物磷含量過(guò)高時(shí)雖然對(duì)生物沒(méi)有直接的毒害,但可能通過(guò)內(nèi)源釋放提高或維持水中的營(yíng)養(yǎng)含量,從而影響生物的生長(zhǎng)(Xie et al,2003)。關(guān)于水中磷含量與浮游藻類(lèi)之間定量關(guān)系的研究已有很多,許多學(xué)者也建立了通過(guò)水中磷含量預(yù)測(cè)藻類(lèi)生物量的定量模型(錢(qián)天鳴等,2002;陳永川等,2010)。在沉積物磷含量較高的富營(yíng)養(yǎng)湖泊中沉水植物難以生存的最根本原因便是藻類(lèi)過(guò)多導(dǎo)致的水下光照不足(Sayer et al, 2010)。王英才(2010)研究發(fā)現(xiàn),草型湖泊中Chl-a通常低于10 μg/L,而藻型湖泊中通常高于36 μg/L。浮游藻類(lèi)過(guò)量增殖后會(huì)對(duì)沉水植物形成遮蔽作用,不利于植物的生存與發(fā)展(Le Bagousse-Pinguet et al,2012;Jackson,2003)。由于湖泊中的磷為封閉型循環(huán),進(jìn)入湖泊的磷元素通過(guò)多種方式在上覆水、生物及沉積物間遷移轉(zhuǎn)化,最終沉降進(jìn)入沉積物中,這可能是導(dǎo)致其相比碳和氮對(duì)浮游藻類(lèi)和沉水植物預(yù)測(cè)力更高的重要原因之一。
3.2"" 沉水植物生長(zhǎng)的沉積物總磷條件需求
目前關(guān)于淺水湖泊沉水植物消失的閾值研究多集中于上覆水中總磷含量對(duì)沉水植物生長(zhǎng)的影響。Jeppesen等(1990)通過(guò)對(duì)丹麥多個(gè)湖泊的比較發(fā)現(xiàn),淺水湖泊由以沉水植物占優(yōu)的清水穩(wěn)態(tài)向以浮游藻類(lèi)占優(yōu)的濁水穩(wěn)態(tài)轉(zhuǎn)換的上覆水總磷閾值為0.050~0.125"mg/L。Wang等(2014)通過(guò)對(duì)長(zhǎng)江中下游多個(gè)湖泊的比較發(fā)現(xiàn),上覆水總磷含量是驅(qū)動(dòng)淺水湖泊穩(wěn)態(tài)轉(zhuǎn)換的關(guān)鍵因子,清水穩(wěn)態(tài)向濁水穩(wěn)態(tài)轉(zhuǎn)換的總磷閾值為0.08~0.12"mg/L,反向轉(zhuǎn)換的閾值為0.04~0.06"mg/L。劉昔等(2023)通過(guò)對(duì)洪湖長(zhǎng)期監(jiān)測(cè)數(shù)據(jù)的分析發(fā)現(xiàn),洪湖清水穩(wěn)態(tài)向濁水穩(wěn)態(tài)轉(zhuǎn)換的總磷閾值為0.092"mg/L,濁水穩(wěn)態(tài)向清水穩(wěn)態(tài)轉(zhuǎn)換的閾值為0.051"mg/L。這些研究結(jié)果表明,在淺水湖泊中沉水植物消失的上覆水總磷閾值,約為0.08~0.12"mg/L。沉積物磷含量雖然也是影響沉水植物的重要因素,但目前關(guān)于淺水湖泊中沉水植物消失的沉積物總磷閾值的定量研究較少。從本研究長(zhǎng)江中下游11個(gè)湖泊的數(shù)據(jù)來(lái)看,沉水植物主要存在的沉積物磷含量為0.34~0.68"g/kg(圖4)。此外,我們進(jìn)一步分析了2003-2020年夏季長(zhǎng)江中下游淺水湖泊有草區(qū)和無(wú)草區(qū)沉積物總磷含量與沉水植物生物量之間的關(guān)系,得到沉水植物消失的沉積物總磷閾值約為0.87~0.98"g/kg(圖5)。此前,關(guān)于沉水植物生長(zhǎng)的沉積物條件閾值的研究多為小尺度模擬實(shí)驗(yàn)。例如在國(guó)外溪流中開(kāi)展的營(yíng)養(yǎng)添加實(shí)驗(yàn)結(jié)果表明,沉積物磷含量在0.04~0.95"g/kg范圍內(nèi)促進(jìn)水生植物的生長(zhǎng),超過(guò)此范圍,開(kāi)始出現(xiàn)抑制植物生長(zhǎng)的現(xiàn)象(Carr,1998),這與本研究獲取的結(jié)果類(lèi)似。而國(guó)內(nèi)部分中宇宙實(shí)驗(yàn)的研究結(jié)果表明,當(dāng)沉積物總磷含量高于1.00"g/kg時(shí),沉水植物生長(zhǎng)狀況相對(duì)較差(謝貽發(fā)等,2007;王立志等,2010;葛緒廣等,2014),這一結(jié)果略高于本研究確定的閾值。這可能是因?yàn)樵谧匀缓粗校统练e物外,許多其他環(huán)境因子也會(huì)對(duì)沉水植物的生長(zhǎng)產(chǎn)生不利影響,尤其是風(fēng)浪、魚(yú)類(lèi)等。因此關(guān)于沉水植物生存所需的沉積物條件閾值還需更多研究加以確定,尤其是多湖比較等大尺度研究。
目前湖泊生態(tài)修復(fù)工程常采用控源截污、生態(tài)疏浚、沉水植物群落重建等方法,其中沉水植物群落重建是生態(tài)修復(fù)工程中常用手段之一,這是因?yàn)槌了参镒鳛楹瓷鷳B(tài)系統(tǒng)的重要組成部分,能夠吸收水體營(yíng)養(yǎng)元素,維持湖泊清水穩(wěn)態(tài),還具有一定的觀賞價(jià)值。研究沉水植物生存所需的沉積物條件閾值能夠?yàn)楹瓷鷳B(tài)修復(fù)工程中選擇沉水植物修復(fù)區(qū)提供參考。此外確定該閾值還能夠?yàn)樯鷳B(tài)疏浚工程中確定生態(tài)疏浚的面積及深度提供依據(jù)。因此,研究沉水植物生存所需的沉積物條件閾值在生態(tài)修復(fù)工程中具有重要的指導(dǎo)意義。
參考文獻(xiàn)
陳洪達(dá), 何楚華, 1975. 武昌東湖水生維管束植物的生物量及其在漁業(yè)上的合理利用問(wèn)題[J]. 水生生物學(xué)集刊, 5(3): 410-420.
陳永川, 張德剛, 湯利, 2010. 滇池水體磷的時(shí)空變化與藻類(lèi)生長(zhǎng)的關(guān)系[J]. 生態(tài)環(huán)境學(xué)報(bào), 19(6):1363-1368.
楚建周, 王圣瑞, 金相燦, 等, 2006. 底質(zhì)營(yíng)養(yǎng)狀況對(duì)黑藻生長(zhǎng)及光合作用的影響[J]. 生態(tài)環(huán)境, 15(4): 702-707.
國(guó)家環(huán)境保護(hù)總局《水和廢水監(jiān)測(cè)分析方法》編委會(huì), 2002. 水和廢水監(jiān)測(cè)分析方法[M]. 4版.北京: 中國(guó)環(huán)境科學(xué)出版社: 243-257.
葛緒廣, 王國(guó)祥, 陳成忠, 等, 2014. 苦草生長(zhǎng)對(duì)沉積物中磷遷移轉(zhuǎn)化的影響[J]. 生態(tài)學(xué)報(bào), 34(20): 5802-5811.
黃祥飛, 陳偉民, 蔡啟銘, 1999. 中國(guó)生態(tài)系統(tǒng)研究網(wǎng)絡(luò)觀測(cè)與分析標(biāo)準(zhǔn)方法:湖泊生態(tài)調(diào)查觀測(cè)與分析[M]. 北京: 中國(guó)標(biāo)準(zhǔn)出版社: 77-80.
魯如坤, 1999. 土壤農(nóng)業(yè)化學(xué)分析方法[M]. 北京: 中國(guó)農(nóng)業(yè)科學(xué)技術(shù)出版社.
劉昔, 厲恩華, 徐杰, 等, 2023. 洪湖濕地生態(tài)系統(tǒng)演變及穩(wěn)態(tài)轉(zhuǎn)換關(guān)鍵驅(qū)動(dòng)因子閾值研究[J]. 湖泊科學(xué), 35(3): 934-940.
李艷, 王洪鑄, 馬碩楠, 等, 2021. 不同外源營(yíng)養(yǎng)負(fù)荷對(duì)浮游藻類(lèi)群落結(jié)構(gòu)特征的影響[J]. 水生生物學(xué)報(bào), 45(6): 1291-1298.
馬金玉, 王文才, 羅千里, 等, 2021. 黃大湖沉積物營(yíng)養(yǎng)鹽分布及來(lái)源解析[J]. 環(huán)境工程技術(shù)學(xué)報(bào), 11(4): 678-685.
秦伯強(qiáng), 高光, 胡維平, 等, 2005. 淺水湖泊生態(tài)系統(tǒng)恢復(fù)的理論與實(shí)踐思考[J]. 湖泊科學(xué), 17(1): 9-16.
錢(qián)天鳴, 陳超, 程詠, 2002. 杭州西湖水體生態(tài)環(huán)境參數(shù)的相互關(guān)系[J].中國(guó)環(huán)境監(jiān)測(cè), 18(3): 41-44.
王洪鑄, 王海軍, 劉學(xué)勤, 等, 2015. 實(shí)施環(huán)境-水文-生態(tài)-經(jīng)濟(jì)協(xié)同管理戰(zhàn)略, 保護(hù)和修復(fù)長(zhǎng)江湖泊群生態(tài)環(huán)境[J]. 長(zhǎng)江流域資源與環(huán)境, 24(3): 353-357.
王立志, 王國(guó)祥, 葛緒廣, 等, 2010.底質(zhì)營(yíng)養(yǎng)鹽負(fù)荷對(duì)輪葉黑藻生長(zhǎng)和光合熒光特性的影響[J]. 生態(tài)學(xué)報(bào), 30(2): 473-480.
王文泉, 張福鎖, 2001. 高等植物厭氧適應(yīng)的生理及分子機(jī)制[J]. 植物生理學(xué)通訊, 37(1): 63-70.
王英才, 2010. 湖泊生態(tài)系統(tǒng)穩(wěn)態(tài)轉(zhuǎn)換過(guò)程及階段劃分研究[D]. 北京: 中國(guó)科學(xué)院研究生院:82-97.
王艷平, 徐偉偉, 韓超, 等, 2021. 巢湖沉積物氮磷分布及污染評(píng)價(jià)[J]. 環(huán)境科學(xué), 42(2): 699-711.
謝貽發(fā), 李傳紅, 劉正文, 等, 2007. 基質(zhì)條件對(duì)苦草(Vallisneria natans)生長(zhǎng)和形態(tài)特征的影響[J]. 農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào), 26(4): 1269-1272.
楊波, 王保棟, 韋欽勝, 等, 2012. 低氧環(huán)境對(duì)沉積物中生源要素生物地球化學(xué)循環(huán)的影響[J]. 海洋科學(xué), 36(5): 124-129.
袁賽波, 張曉可, 劉學(xué)勤, 等, 2019. 長(zhǎng)江中下游湖泊水生植被的生態(tài)水位管理策略[J]. 水生生物學(xué)報(bào), 43(增刊1): 104-109.
鄢文皓, 王會(huì)會(huì), 李前正, 等, 2020. 影響沉水植物恢復(fù)的環(huán)境閾值研究進(jìn)展[J].生態(tài)科學(xué), 39(5): 240-247.
生態(tài)環(huán)境部, 國(guó)家市場(chǎng)監(jiān)督管理總局, 2018. 土壤環(huán)境質(zhì)量 農(nóng)用地土壤污染風(fēng)險(xiǎn)管控標(biāo)準(zhǔn): GB 15618—2018[S]. 北京: 中國(guó)標(biāo)準(zhǔn)出版社.
曾巾, 楊柳燕, 肖琳, 等, 2007. 湖泊氮素生物地球化學(xué)循環(huán)及微生物的作用[J]. 湖泊科學(xué), 19(4): 382-389.
Bachmann R W, Horsburgh C A, Hoyer M V, et al, 2002. Relations between trophic state indicators and plant biomass in Florida lakes [J]. Hydrobiologia, 470(1): 219-234.
Barko J W, Smart R M, 1978. The growth and biomass distribution of two emergent freshwater plants, Cyperus esculentus and Scirpus validus, on different sediments [J]. Aquatic Botany, 5: 109-117.
Brix H, 1987. Treatment of wastewater in the rhizosphere of wetland plants:the root-zone method [J]. Water Science and Technology, 19(1/2): 107-118.
Cao T, Ni L Y, Xie P, 2004. Acute biochemical responses of a submersed macrophyte, Potamogeton crispus L., to high ammonium in an aquarium experiment [J]. Journal of Freshwater Ecology, 19(2): 279-284.
Carr G M, 1998. The growth and biomass distribution of two emergent freshwater and sediment phosphorus and nitrogen in a Canadian prairie river [J]. Freshwater Biology, 39: 525-536.
H?kanson L, Jansson M, 1983.Principles of Lake Sedimentology[M]. Berlin, Heidelberg: Springer Berlin Heidelberg.
Jackson L J, 2003. Macrophyte-dominated and turbid states of shallow lakes: evidence from Alberta Lakes [J]. Ecosystems, 6(3): 213-223.
Jeppesen E, Jensen J P, Kristensen P, et al, 1990. Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes 2: threshold levels, long-term stability and conclusions [J]. Hydrobiologia, 200/201(1): 219-227.
Kozela C, Regan S, 2003. How plants make tubes [J]. Trends in Plant Science, 8(4): 159-164.
Le Bagousse-Pinguet Y L, Liancourt P, Gross N, et al, 2012. Indirect facilitation promotes macrophyte survival and growth in freshwater ecosystems threatened by eutrophication [J]. Journal of Ecology, 100(2): 530-538.
Liu X Q, Yang Z D, Yuan S B, et al, 2017. A novel methodology for the assessment of water level requirements in shallow lakes [J]. Ecological Engineering, 102: 31-38.
Sayer C D, Davidson T A, Jones J I, 2010. Seasonal dynamics of macrophytes and phytoplankton in shallow lakes: a eutrophication-driven pathway from plants to plankton? [J].Freshwater Biology, 55(3): 500-513.
Scheffer M, 1998. Ecology of Shallow Lakes [M]. Dordrecht: Kluwer Academic Publishers: 76-118.
Schindler D W, Hecky R E, Findlay D L, et al, 2008. Eutrophication of lakes cannot be controlled by reducing nitrogen input: results of a 37-year whole-ecosystem experiment [J]. Proceedings of the National Academy of Sciences of the United States of America, 105(32): 11254-11258.
Sondergaard M, Jensen P J, Jeppesen E, 2001. Retention and internal loading of phosphorus in shallow, eutrophic lakes [J]. The Scientific World Journal, 1: 427-442.
van Wijck C V, de Groot C-J,Crillas P, 1992. The effect of anaerobic sediment on the growth of Potamogeton pectinatus L.: the role of organic matter, sulphide and ferrous iron[J]. Aquatic Botany, 44(1):31-49.
Wang H J, Wang H Z, Liang X M, et al, 2014. Total phosphorus thresholds for regime shifts are nearly equal in subtropical and temperate shallow lakes with moderate depths and areas [J]. Freshwater Biology, 59(8): 1659-1671.
Wetzel R G, 2001. Limnology: lake and river ecosystems" [M]. 3rd Ed. San Diego: Academic Press: 527-575.
Wu J, Dai Y R, Rui S Y, et al, 2015. Acclimation of Hydrilla verticillata to sediment anoxia in vegetation restoration in eutrophic waters [J]. Ecotoxicology, 24(10): 2181-2189.
Xie L Q, Xie P, Tang H J, 2003. Enhancement of dissolved phosphorus release from sediment to lake water by Microcystis blooms: an enclosure experiment in a hyper-eutrophic, subtropical Chinese Lake [J]. Environmental Pollution, 122(3): 391-399.
Yu Q, Wang H Z, Li Y, et al, 2015. Effects of high nitrogen concentrations on the growth of submersed macrophytes at moderate phosphorus concentrations [J]. Water Research, 83: 385-395.
(責(zé)任編輯"" 鄭金秀)
Relationship of Biogenic Substances in Sediments with Phytoplankton
and Submerged Macrophytes in Shallow Lakes
LIU Jia‐hao1,2, LI Yan2,SHEN Dong‐fang2,3, QIAO Rui‐ting2,3, WANG Hong‐zhu2
(1. College of Fisheries and Life Science, Dalian Ocean University, Dalian"" 116023, P. R. China;
2. State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology,
Chinese Academy of Sciences, Wuhan"" 430072, P. R. China;
3. University of Chinese Academy of Sciences, Beijing"" 100049, P. R. China)
Abstract:In this study, we explored the effects of biogenic substances in sediments on phytoplankton and submerged macrophytes in shallow lakes in the middle and lower reaches of the Yangtze River basin. The study was based on summer investigations of water quality, biogenic substances in sediments and submerged macrophytes in 11 shallow lakes from 2003 to 2020 (Baoan Lake during the summers of 2003-2012, 2020; Hongxing, Honghu, Nanhu, Qingling, Sanliqi and Zhangdu Lakes in the summer of 2003; Dianshan, Gehu, Wuchang and Yangcheng Lakes in the summer of 2004). The relationship between sediment nutrient content and algal or macrophyte dominance was analyzed using univariate linear regression. Results show that: (1) The total phosphorus content in sediment (TPSed) was highest in algal-dominated lakes (0.71±0.21) g/kg and lowest in macrophyte-dominated lakes (0.52±0.13) g/kg. Total nitrogen content (TNSed) and total organic carbon content (TOCSed) in sediment were highest in macrophyte-dominated lakes [TNSed: (4.64±3.57) g/kg; TOCSed: (55.89±50.28) g/kg] and lowest in intermediate lakes [TNSed: (1.86±1.26) g/kg; TOCSed: (20.00±12.13) g/kg]. (2) Regression analysis shows that phytoplankton chlorophyll-a content (Chl-a) increased and submerged macrophytes biomass (BMac) decreased significantly at high TPSed (Plt;0.05). However, TNSed and TOCSed had little effect on Chl-a and BMac (Pgt;0.17). (3) TPSed thresholds for the disappearance of submerged macrophytes in summer was 0.87-0.98 g/kg in the lakes of the middle and lower reaches of the Yangtze River basin. Our results demonstrate that TPSed is a better predictor of Chl-a and BMac than TNSed and TOCSed in shallow lakes of the middle and lower reaches of the Yangtze River basin. More quantitative analysis of these relationships will provide a scientific basis for assessing sediment pollution in shallow lakes and determining sediment conditions necessary for restoring and maintaining the submerged macrophyte community.
Key words:lakes in the middle and lower reaches of the Yangtze River; sediment; nutrient content; submerged macrophytes
