紅藍光配比對黃瓜幼苗光合特性的影響
2024-01-02 00:00:00王騰起孫啟穎劉彬彬李清明
山東農業大學學報(自然科學版) 2024年6期


摘要:以‘津優365’黃瓜品種為試材,設置白光(W)、單色紅光(R)、單色藍光(B)以及不同比例的紅藍復合光(9R1B、7R3B、5R5B、3R7B、1R9B)8 個光質處理,研究了紅藍光對黃瓜幼苗穩態光合作用與動態光合作用的影響,以期為提高黃瓜幼苗在不同環境條件下的光合能力提供理論基礎。結果表明,與紅光相比,藍光在提高黃瓜幼苗最大光合能力(Pmax)方面發揮重要作用,其主要通過提高表觀量子利用效率(AQE)、Rubisco 最大羧化效率(Vcmax)、最大電子傳遞速率(Jmax)、磷酸丙糖利用效率(TPU)與氣孔導度(Gs)提高Pmax。在紅藍復合光處理下,AQE、Vcmax、Jmax、TPU與Pmax隨著藍光比例的上升而提高,但1R9B處理與5R5B和3R7B處理之間無明顯差異。此外,適宜比例的藍光(30%-70%)可以提高凈光合速率(A)的誘導速度,降低光合速率升高90%所需時間(T90%A)。1R9B 處理下T90%A 顯著高于7R3B、5R5B 與3R7B處理,說明過量的藍光會降低光合誘導速度。綜上所述,適宜的藍光比例不僅可以提高黃瓜幼苗的最大光合能力,還可以加快光合誘導速度,5R5B與3R7B處理下黃瓜幼苗最大光合能力與動態光合能力最佳。
關鍵詞:黃瓜;紅藍光;穩態光合作用;動態光合作用
中圖法分類號: S642.2 文獻標識碼: A 文章編號: 1000-2324(2024)06-0859-08
黃瓜(Cucumis sativus L.)具有產量高、營養豐富、經濟效益高等優點[1],在世界范圍內被廣泛種植[2,3]。在生產過程中,為了提高黃瓜產量、品質與生長速度,經常采用工廠化育苗的方式培養壯苗[4]。在工廠化育苗中,使用環境條件高度可控的植物工廠進行育苗是工廠化育苗最高級的方式,且在植物工廠中,常使用全人工光源替代自然光提供植物生長所需的光能[5]。與自然光相比,人工光具有更高的可控性,可以根據幼苗生長的需求精準調控光強、光質、光周期等光環境特征,因此有必要深入研究光強、光質、光周期等重要光環境特征對植物幼苗的影響,為提高幼苗生長速度與品質提供理論基礎[6]。作為光環境的重要組成部分,光質會顯著影響植物的生長發育與光合特征[7,8]。其中,紅藍光是維持光合作用正常進行的重要光質,在育苗過程中一般以紅藍復合光作為光源[9,10]。
植物在某一特定環境條件下的光合作用被稱為穩態光合,如植物在飽和光強下的光合速率被稱為最大光合能力,目前已得到廣泛研究[11,12]。有研究表明,紅藍光在維持光合作用正常進行,提高植物光合能力方面發揮更重要的作用[13,14]。與紅光相比,藍光在維持光合作用正常進行方面發揮更重要的作用[15,16]。單色紅光往往會導致紅光綜合癥,表現為光系統發育受阻、氣孔發育不良且氣孔導度降低,從而嚴重降低植物的光合速率[17,18]。在紅光基礎上添加少量藍光可以有效緩解紅光綜合征。有研究發現在紅光基礎上添加藍光可以促進番茄葉綠素合成與電子傳遞,從而提高其光合能力[19]。但也有研究發現,藍光強度過高會對光合作用產生不利影響。如與紅光相比,藍光造成的番茄光系統Ⅱ處的光抑制現象更加嚴重,且在更低的光強下就會發生[16];在紅藍復合光處理下藍光比例超過66%后會抑制生菜生長[20]。綜上所述,適宜強度的藍光可以提高植物的最大光合能力[21]。
受晝夜節律與云層遮擋影響,自然界中的光強往往處于波動狀態,而光強的波動會影響植物的光合作用[22]。近些年,有研究發現影響植物生長的因素不僅是特定光強下的光合能力,還包括光合作用響應光強波動的能力,即動態光合作用[23]。有研究發現,波動光下限制擬南芥與煙草光合作用的因素主要是氣孔導度,通過加快氣孔開放與關閉可以顯著提高擬南芥與煙草的動態光合能力[24]。提高植物的動態光合能力可以促進波動光條件下植物的生長,有研究發現提高油菜單位葉面積上的氮含量可以加速油菜的光誘導,并提高干物質積累量[25];對比不同品種水稻的氣孔特征與光誘導特性后發現,氣孔密度大且氣孔面積較小的水稻氣孔導度升高更快,從而加速光合誘導,以提高波動光下水稻的生長量[26]。關于光質對植物動態光合作用的影響,不同研究得出了不同的結論,有研究發現紅藍比對番茄穩態光合作用影響較大,但對動態光合作用影響較小[27]。而有研究則發現藍光在提高黃瓜幼苗最大光合能力與氣孔導度的同時,還會加快光合速率和氣孔導度的誘導速度[28]。
本試驗通過對比紅光、藍光以及不同比例的紅藍復合光對黃瓜幼苗穩態光合作用與動態光合作用的影響,將進一步探明紅藍光在調控黃瓜幼苗穩態光合作用與動態光合作用中發揮的作用,為提高動態光環境下黃瓜幼苗的光合能力提供新方法。
1 材料與方法
1.1 試驗材料
以黃瓜(Cucumis sativus L.)品種‘津優365’為試材,采用穴盤育苗的方式獲得供試黃瓜幼苗,育苗方法如下:將種子浸泡在去離子水中使其充分吸水,于28 ℃恒溫培養箱中催芽至種子露白后,點播于72 孔穴盤中,穴盤中填充混合基質(草炭:蛭石:珍珠巖=3∶1∶1)。育苗在人工氣候室中進行,溫度為25±2 ℃,空氣濕度為60%,光照強度為200 μmol·m?2·s?1,光質為白光,光周期為10 h·d?1。待幼苗子葉完全展平時,挑選長勢一致的幼苗移栽到含15 L營養液的黑色不透明塑料盆中,每盆種植16棵黃瓜幼苗。水培時使用二分之一劑量的日本山崎(Yamazaki)營養液(NH4H2PO4濃度為0.5 mmol·L?1, Ca(NO3)2·4H2O濃度為2.0 mmol·L?1,KNO3濃度為3.2 mmol·L?1,MgSO4·7H2O濃度為1.0 mmol·L?1,并添加全劑量的微量元素),并用充氧泵為黃瓜根系供氧。營養液每3 d 更換一次。
1.2 試驗設計
設置白光(W)、單色紅光(R)、90%紅光與10% 藍光(9R1B)、70% 紅光與30% 藍光(7R3B)、50%紅光與50%藍光(5R5B)、30%紅光與70% 藍光(3R7B)、10% 紅光與90% 藍光(1R9B)以及單色藍光(B)8 個處理,光質處理9 d后測定相關指標。紅光波峰為660 nm,藍光波峰為450 nm。環境條件為:溫度25±3 ℃,空氣濕度60%,光強100 μmol·m?2·s?1,光周期14 h·d?1。使用Heliospectra DYNA LED 燈(Heliospectra,Sweden)作為光源。
1.3 試驗方法
(1)光響應曲線:使用LI-6800 光合儀測定光響應曲線,測定時氣體流速為500 μmol·s?1,葉室內空氣濕度為60%,二氧化碳濃度為400 μmol·mol?1,風扇轉速為10 000 rpm,葉片溫度為25 ℃,光源為紅藍復合光,紅藍比為8∶2。測定前,葉片在光強1 500 μmol·m?2·s?1下充分光適應。光強梯度為1 500、1 200、1 000、800、600、400、200、150、100、50、0 μmol·m?2·s?1;每個光強下等待3-5 min。根據直角雙曲線模型擬合光響應曲線[29]。
(2)二氧化碳響應曲線:傳統二氧化碳響應曲線(A-Ci)測定時,環境參數的設置與光響應曲線一致,在測定開始前同樣需進行充分的光適應。二氧化碳濃度梯度為:400、300、200、100、50、400、600、800、1 000、1 200、1 500 μmol·mol?1,每個二氧化碳濃度下等待時間為3-5 min,測定一條傳統二氧化碳響應曲線所需的時間為33-55 min。
除測定傳統二氧化碳響應曲線外,還進行了快速二氧化碳響應曲線(RACiR)的測定[30,31]。測定時設定起始二氧化碳濃度為10 μmol·mol?1,最高二氧化碳濃度為1 500 μmol·mol?1,二氧化碳濃度變化速率為100 μmol·mol?1·min?1,計數間隔為2 s。測定一條快速二氧化碳響應曲線所需時間為16 min,與傳統二氧化碳響應曲線相比節約了50%左右的時間。通過對比兩種測定方法得出的結果,可以提高測量結果的可靠性。使用“msuRACiFit”擬合工具擬合二氧化碳響應曲線[32]。
(3)弱光-強光轉換過程中的光合參數:環境參數與光響應曲線測定一致。使用一步光強變化法評價黃瓜幼苗的動態光合能力[26]。首先,在100 μmol·m?2·s?(1 生長光強)下測定3 min,之后光強升高至1 500 μmol·m?2·s?(1 飽和光強)測定20 min,數據記錄間隔為10 s。可以得到以下參數:黃瓜幼苗在光強為100 μmol·m?2·s?1時的光合速率(A100)和氣孔導度(Gs100)、在光強為1 500 μmol·m?2·s?1時的光合速率(A1500)和氣孔導度(Gs1500)。并計算光合速率在特定時間的誘導狀態(ISt)[26]:
1.4 數據處理
使用Microsoft Excel 2016 (MicrosoftCorporation,USA)計算平均值與標準差;使用SPSS 16.0(IBM,USA)對數據進行數據正態性檢驗與方差齊性檢驗。使用Duncan 法比較不同處理組之間的差異(Plt;0.05),并使用Origin 2021(OriginLab Corporation,USA)繪圖。
2 結果與分析
2.1 紅藍光處理對光響應曲線相關擬合參數的影響
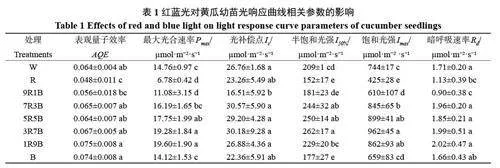
如表1 所示,R 與9R1B 處理下黃瓜幼苗表觀量子效率(AQE)較低,與W處理相比分別降低了24.79% 與12.40%,而1R9B 與B 處理下AQE較高,與W處理相比分別提高了17.22%與16.78%。R 處理顯著降低了Pmax,B 對Pmax 影響較小。紅藍復合光處理下,9R1B處理Pmax最低,而當藍光比例超過50%后,Pmax顯著升高,并在1R9B 處理下達到最高。光補償點(Ic)僅在9R1B 處理下顯著降低,其它處理下無顯著差異。不同光質對半飽和光強(I50%)與飽和光強(Imax)的影響基本相同,與W處理相比,R與B處理降低了I50%與Imax,且R 處理下降幅更大,與W處理相比分別降低了27.25%與42.87%。紅藍復合光處理下,隨著藍光比例的上升,I50%與Imax呈上升趨勢,在3R7B處理下達到最大。不同光質對暗呼吸速率(Rd)的影響較小,僅在R 與9R1B處理下顯著降低。綜上所述,紅光會降低黃瓜幼苗的光合能力、光量子利用效率、半飽和光強、飽和光強與暗呼吸速率,而藍光的作用則相反。
2.2 紅藍光處理對二氧化碳響應曲線相關擬合參數的影響
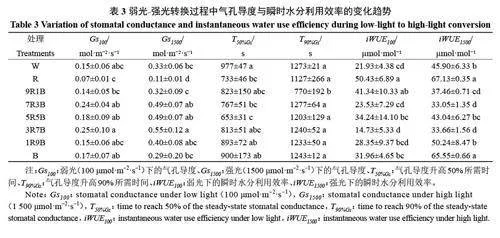
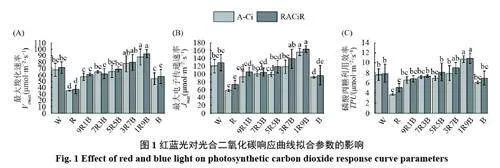
如圖1 所示,與W處理相比,R 處理顯著降低了Vcmax、Jmax與TPU,B處理的作用于R處理相似,但下降幅度要小于R處理。在紅藍復合光處理下,Vcmax、Jmax與TPU隨著藍光比例的上升而上升,在9R1B 處理下較低,但與W處理相比無顯著差異;在1R9B 處理下達到最高,顯著高于W處理。Vcmax、Jmax與TPU在R處理下達到最低,在1R9B處理下達到最高。綜上所述,單色紅光或單色藍光都會降低Vcmax、Jmax與TPU,藍光在提高最大光合能力方面發揮更重要的作用,在紅藍復合光處理下,Vcmax、Jmax與TPU著藍光比例的上升而上升。
2.3 紅藍光處理對弱光-強光轉換過程中黃瓜幼苗氣體交換參數的影響
如表2所示,黃瓜幼苗在弱光(100 μmol·m?2·s?1)下的光合速率(A100)在R處理下較低,與W處理相比下降了20.95%,但差異并未達到顯著水平。與W 處理相比,R 處理下A1500 顯著下降了51.46%,而B處理下A1500與W處理無顯著差異;在紅藍復合光處理下,A1500隨著藍光比例的上升而上升,7R3B、5R5B、3R7B 與1R9B 處理下A1500顯著升高,并在3R7B處理下達到最高。計算光合速率在光強升高后第60 s 時的誘導狀態(IS60s)和第300 s 時的誘導狀態(IS300s)后發現,與W處理相比,IS60s與IS300s在R處理下顯著降低,分別降低了31.04%和37.79%,而在B 處理下變化不明顯。在紅藍復合光處理下,與W處理相比,9R1B處理下IS60s下降不明顯,但IS300s顯著下降,與W處理相比下降了23.59%,整體來看,7R3B 與3R7B 處理下IS60s 與IS600s 較高,1R9B 處理下較低,但都高于9R1B處理。R處理顯著提高了光合速率升高50%需要的時間(T50%A),但對光合速率升高90%所需時間(T90%A)的提升較小。在紅藍復合光處理下,9R1B 與1R9B 處理下T50%A 與T90%A較高,但與W處理相比無顯著差異。綜上所述,紅光會降低黃瓜幼苗的光合速率與光合誘導速度;而適當比例的藍光可以提高強光下黃瓜幼苗的光合速率并加快光合誘導速度。
如表3 所示,弱光下的氣孔導度(Gs100)在3R7B 處理下達到最高,與W處理相比提高了62.29%,顯著高于W處理;在R處理下達到最低,與W處理相比下降了55.55%,顯著低于W處理。在其它處理下,Gs100無顯著差異。強光下的氣孔導度(Gs1500)在R處理下顯著降低,與W處理相比下降了68.49%,在3R7B處理下顯著升高,與W處理相比升高了65.92%。在紅藍復合光下,除9R1B與1R9B處理下Gs1500較低外,其它處理下Gs1500無顯著差異。紅藍復合光下,Gs1500在9R1B處理下達到最低,與B處理無差異,但顯著高于R處理。計算光強升高后氣孔導度升高50%(T50%Gs)與90%所需時間(T90%Gs)后發現,與W處理相比,R、7R3B與5R5B處理下T50%Gs顯著降低,與W處理相比分別下降了24.91%、21.50%與33.11%,其它處理下T50%Gs與白光相比無顯著差異。T90%Gs僅在9R1B處理下顯著降低,與W處理相比下降了39.53%。弱光下的瞬時水分利用效率(iWUE100)在R處理與9R1B 處理下較高,與W處理相比分別提高了129.94%與88.48%,差異達到顯著水平;在3R7B處理下較低,但與W處理的差異并未達到顯著水平;在其他處理下iWUE100與W處理無顯著差異。強光下的瞬時水分利用效率(iWUE1500)在R與B處理下顯著高于W處理,與W處理相比分別提高了46.25%與42.82%,在9R1B、7R3B與3R7B處理下顯著低于W處理,分別降低了18.38%、28.00%與26.67%。綜上所述,紅光會降低氣孔導度,但會提高氣孔導度的上升速度及瞬時水分利用效率。紅藍復合光處理下,藍光比例上升在提高氣孔導度的同時并未顯著提高氣孔導度的上升速度。
3 討論
不同光質對黃瓜幼苗的穩態光合作用產生了顯著影響。整體來看,單色紅光處理下黃瓜幼苗穩態光合能力較差,主要通過降低表觀量子效率(AQE)、最大光合速率(Pmax)、最大羧化速率(Vcmax)、最大電子傳遞速率(Jmax)以及磷酸丙糖利用效率(TPU)顯著降低了黃瓜幼苗的最大光合能力(表1,圖1)。AQE 是通過計算光響應曲線在弱光階段的斜率得出的,AQE較低代表植物利用光能的能力較弱,而Pmax與Imax的降低則說明植物的光合潛能以及抵抗強光的能力降低,導致光合速率在低光強下達到最高,無法利用更多的光能,這一般是由于光合機構發育不良導致的[33]。Vcmax是在二氧化碳響應曲線初始階段,即低二氧化碳濃度下限制光合能力提高的主要因素。此時葉綠體羧化位點處的二氧化碳濃度較低,限制光合能力上升的原因主要是卡爾文循環關鍵酶Rubisco 的羧化活性[34]。而隨著二氧化碳濃度的升高,限制因素開始轉變為羧化反應底物RuBP再生速率的限制,RuBP 的再生又要依靠光合電子傳遞過程中產生的ATP 與NADPH,所以最大電子傳遞速率Jmax在一定程度上代表RuBP 的再生速率,提高Jmax是提高光合速率的重要手段[35]。在二氧化碳響應曲線的最后一個階段,即二氧化碳濃度過高后,限制光合速率上升的主要因素轉變為光合產物的利用效率,即TPU。TPU高代表植物可以將光合產物快速運離羧化位點,防止光合產物過分積累抑制光合作用[32]。單色紅光下AQE、Pmax、Vcmax、Jmax與TPU顯著下降(表1,圖1),說明單色紅光通過降低光能利用率、Rubisco 活性、電子傳遞以及磷酸丙糖利用效率降低黃瓜幼苗的最大光合能力。而藍光在促進光合機構發育,提高黃瓜最大光合能力方面發揮更重要的作用,這與藍光在弱光下促進黃瓜葉片葉綠體與光系統發育從而提高黃瓜幼苗光合能力的結果相同[36]。同時Miao 等也發現與紅光相比藍光可以提高黃瓜光系統活性、氣孔導度、Vcmax與Jmax從而提高最大光合能力[37]。在紅藍復合光下,隨著藍光比例的上升,AQE、Pmax、Imax、Vcmax、Jmax與TPU呈上升趨勢,證明了藍光在提高黃瓜幼苗最大光合能力方面發揮重要作用,且藍光比例達到90%后并未降低黃瓜幼苗的最大光合能力,說明在紅藍復合光下,藍光比例過高并不會對黃瓜幼苗的最大光合能力產生不利影響,與之前研究得出的紅藍復合光下提高藍光比例可以提高黃瓜幼苗最大光合能力的結論相同[38]。綜上所述,與紅光相比藍光在提高黃瓜幼苗最大光合能力方面發揮更大的作用,但紅光完全缺失同樣會降低最大光合能力,在藍光基礎上添加少量(10%)紅光即可最大限度地提高黃瓜幼苗的最大光合能力,進一步證明了紅藍光可以協同提高植物的最大光合能力。
自然界中的光強度往往處于不斷的變化中,因此植物在某一特定光強度下的光合能力很難與生長量對應,因此有必要研究光合作用對光強變化的響應[39,40]。本研究通過研究弱光-強光轉換過程中光合參數的變化趨勢發現:弱光下的光合速率與氣孔導度變化較小,僅在單色紅光下小幅度下降;在紅藍復合光下,藍光比例的上升提高了強光下的光合速率與氣孔導度,但藍光比例過高(90%)會引起光合速率與氣孔導度發生不明顯的下降,紅光則顯著降低了強光下的光合速率與氣孔導度,但瞬時水分利用效率最高(表2-3)。以上結果說明紅光對最大光合能力的負面影響更大,但可以提高水分利用效率,而藍光可以促進氣孔開放從而提高植物的光合能力,但氣孔導度的升高會導致水分利用效率的降低,這與之前的結論相似[38]。藍光比例過高會影響氣孔開放與光合作用,這與之前在大豆幼苗上得出的結論一致,主要是由于藍光的光量子利用效率低于紅光,從而影響光合電子傳遞過程導致的[41]。對比光合速率在不同時間的誘導狀態以及光合速率和氣孔導度到達特定誘導狀態所需時間后發現:單色紅光顯著降低了弱光-強光轉換過程中光合速率的誘導速度,而單色藍光下光合誘導速度與白光相似;在紅藍復合光處理下,9R1B與1R9B降低了光合速率的誘導速度,其它復合光處理下誘導速度較快(表2)。這說明雖然藍光可以加快光合速率的誘導速度,但藍光比例太高會對光合誘導速度產生不利影響,證明相比于最大光合能力,黃瓜幼苗的動態光合能力更容易受到高比例藍光的抑制。對比氣孔導度的誘導速度后發現,在紅藍復合光下藍光比例的上升并沒有提高氣孔導度的誘導速度;雖然9R1B處理可以提高氣孔的誘導速度(表3),但這很可能是由于最大氣孔導度降低導致的。這與Li 等得出的藍光可以提高黑暗-強光轉換過程中黃瓜幼苗氣孔開放速度的結果不同[28],可能是由于測定方法不同,以及初始氣孔導度不同導致的,說明植物的初始氣孔導度很可能會影響之后的光誘導進程。
4 結論
單色紅光通過降低AQE、Vcmax、Jmax、TPU與Gs降低黃瓜幼苗的最大光合能力,但對弱光下的光合速率影響較小,同時可以通過降低最大氣孔導度加快弱光-強光轉換過程中氣孔導度的誘導速度。藍光不僅可以提高黃瓜幼苗的最大光合能力,還會提高弱光-強光轉換過程中光合速率的上升速度。藍光比例過高對黃瓜幼苗最大光合能力的影響較小,但會降低弱光-強光轉換過程中光合速率的誘導速度,藍光比例在50%-70%之間可以有效提高黃瓜幼苗的最大光合能力與動態光合能力。
參考文獻
[1] 王 新. 淺析北方蔬菜營養價值與栽培模式[J]. 農業
開發與裝備,2022(04):177-179.
[2] 李艾潼,張晨陽,王思惠,等. 黃瓜國內外分級標準對
比及發展現狀分析[J]. 食品安全導刊,2020(09):
24-26.
[3] 李懷智. 我國黃瓜栽培的現狀及其發展趨勢[J]. 蔬
菜,2003(08):3-4.
[4] 魏云鵬. 目前我國黃瓜栽培茬口及播種時間[J]. 吉
林蔬菜,2004(04):39.
[5] 楊 凡,蔡毓新,田昭然,等. 黃瓜雙斷根嫁接工廠化
育苗關鍵技術[J]. 中國瓜菜,2020,33(05):90-92.
[6] 李 光. 人工光源對黃瓜幼苗生長發育的影響[J]. 上
海蔬菜,2020(03):55-56.
[7] 何偉明,劉龐源,武占會,等. 基于不同溫度的不同光
質處理對黃瓜幼苗生長的影響[J]. 北方園藝,2017
(09):30-33.
[8] Jeong HW, Lee HR, Kim HM, et al. Using light
quality for growth control of cucumber seedlings in
closed-type plant production system[J]. Plants, 2020,
9(5):639.
[9] 譚杰揮,劉厚誠. 植物工廠蔬菜育苗光調控技術研
究進展[J]. 農業工程技術,2022,42(01):24-29.
[10] 汪 穎,楊新琴,徐 沛,等. 不同紅藍光配比LED光源
對蔬菜育苗的影響[J]. 浙江農業科學,2020,61(04):
649-653.
[11] 葉子飄. 光合作用對光和CO2響應模型的研究進展
[J]. 植物生態學報,2010,34(06):727-740.
[12] 葉子飄,于 強. 光合作用對胞間和大氣CO2響應曲
線的比較[J]. 生態學雜志,2009,28(11):2233-2238.
[13] Gon?alves VD, Fagundes-Klen MR, Trigueros DEG,
et al. Combination of Light Emitting Diodes (LEDs)
for photostimulation of carotenoids and chlorophylls
synthesis in Tetradesmus sp[J].Algal Research, 2019,
43:101649.
[14] Liu J, Van Iersel MW. Photosynthetic physiology of
blue, green, and red light: light intensity effects and
underlying mechanisms[J]. Frontiers in Plant
Science, 2021,12:619987.
[15] 張 歡,徐志剛,崔 瑾,等. 光質對番茄和萵苣幼苗生
長及葉綠體超微結構的影響[J]. 應用生態學報,
2010,21(04):959-965.
[16] 謝 鑫,王俊玲,段立肖,等. 光質對番茄葉片葉綠素
熒光動力學參數的影響[J]. 河北農業大學學報,
2013,36(06):48-51+88.
[17] Miao Y, Chen Q, Qu M, et al. Blue light alleviates 'red
light syndrome' by regulating chloroplast ultrastructure,
photosynthetic traits and nutrient accumulation in
cucumber plants[J]. Scientia Horticulturae, 2019, 257:
108680.
[18] Chen XL, Li YL, Wang LC, et al. Red and blue
wavelengths affect the morphology, energy use
efficiency and nutritional content of lettuce (Lactuca
sativa L.)[J].Scientific Reports, 2021(1),11:8374.
[19] 楊俊偉,鮑恩財,張珂嘉,等. 不同紅藍光比例對番茄
幼苗葉片結構及光合特性的影響[J]. 西北農業學
報,2018,27(05):716-726.
[20] Clavijo-Herrera J, Van Santen E, Gómez C. Growth,
water-use efficiency, stomatal conductance, and
nitrogen uptake of two lettuce cultivars grown under
different percentages of blue and red light[J].
Horticulturae, 2018,4(3):16-29.
[21] Hogewoning SW, Wientjes E, Douwstra P, et al.
Photosynthetic quantum yield dynamics: from
photosystems to leaves[J].Plant Cell, 2012,24(5):1921-
1935.
[22] Matsuda R, Ito H, Fujiwara K. Effects of artificially
reproduced fluctuations in sunlight spectral distribution
on the net photosynthetic rate of cucumber leaves[J].
Frontiers in Plant Science, 2021,12:675810.
[23] Qu M, Hamdani S, Li W, et al. Rapid stomatal response
to fluctuating light: an under-explored mechanism to
improve drought tolerance in rice[J]. Function Plant
Biology, 2016,43(8):727-738.
[24] Sakoda K, Yamori W, Groszmann M, et al. Stomatal,
mesophyll conductance, and biochemical limitations to
photosynthesis during induction[J]. Plant Physiology,
2021,185(1):146-160.
[25] Liu J, Zhang J, Estavillo GM, et al. Leaf N content
regulates the speed of photosynthetic induction under
fluctuating light among canola genotypes (Brassica
napus L.) [J]. Physiologia Plantarum, 2021, 172(4):
1844-1852.
[26] Xiong Z, Xiong D, Cai D, et al. Effect of stomatal
morphology on leaf photosynthetic induction under
fluctuating light across diploid and tetraploid rice[J].
Environmental and Experimental Botany, 2022, 194:
104757.
[27] Zhang Y, Kaiser E, Zhang Y, et al. Red/blue light
ratio strongly affects steady-state photosynthesis, but
hardly affects photosynthetic induction in tomato
(Solanum lycopersicum) [J]. Physiologia Plantarum,
2019,167(2):144-158.
[28] Li X, Zhao S, Lin A, et al. Effect of different ratios of
red and blue light on maximum stomatal conductance
and response rate of cucumber seedling leaves[J].
Agronomy, 2023,13(7):1941-1954.
[29] Lobo FDA, Barros MP, Dalmagro HJ, et al. Fitting net
photosynthetic light-response curves with Microsoft
Excel - a critical look at the models[J].Photosynthetica,
2013,51(3):445-456.
[30] Stinziano JR, Morgan PB, Lynch DJ, et al. The rapid
A-Ci response: photosynthesis in the phenomic era[J].
Plant Cell and Environment, 2017,40(8):1256-1262.
[31] Saathoff AJ, Welles J. Gas exchange measurements
in the unsteady state[J]. Plant Cell and Environment,
2021,44(11):3509-3523.
[32] Gregory LM, McClain AM, Kramer DM, et al. The
triose phosphate utilization limitation of photosynthetic
rate: Out of global models but important for leaf
models[J]. Plant Cell and Environment, 2021, 44(10):
3223-3226.
[33] 劉福霞,劉乃森. 不同光質對黃瓜幼苗生理生化特
性的影響[J]. 湖北農業科學,2016,55(07):1722-1725.
[34] Sun JL, Sui XL, Huang HY, et al. Low light stress
down-regulated rubisco gene expression and
photosynthetic capacity during cucumber (Cucumis
sativus L.) leaf development[J].Journal of Integrative
Agriculture, 2014,13(5):997-1007.
[35] Raines CA. Improving plant productivity by re-tuning
the regeneration of RuBP in the Calvin-Benson-
Bassham cycle[J]. New Phytologist, 2022, 236(2):
350-356.
[36] Wang XY, Xu XM, Cui J. The importance of blue light
for leaf area expansion, development of photosynthetic
apparatus, and chloroplast ultrastructure of Cucumis
sativus grown under weak light[J]. Photosynthetica,
2015,53(2):213-222.
[37] Miao YX, Wang XZ, Gao LH, et al. Blue light is
more essential than red light for maintaining the
activities of photosystem Ⅱ and Ⅰ and photosynthetic
electron transport capacity in cucumber leaves[J].
Journal of Integrative Agriculture, 2016,15(1):87-100.
[38] Hogewoning SW, Trouwborst G, Maljaars H, et al.
Blue light dose-responses of leaf photosynthesis,
morphology, and chemical composition of Cucumis
sativus grown under different combinations of red
and blue light[J]. Journal of Experimental Botany,
2010,61(11):3107-3117.
[39] Zhang Y, Kaiser E, Marcelis LFM, et al. Salt stress and
fluctuating light have separate effects on photosynthetic
acclimation, but interactively affect biomass[J]. Plant
Cell and Environment, 2020,43(9):2192-2206.
[40] Xu J, Guo Z, Jiang X, et al. Light regulation of
horticultural crop nutrient uptake and utilization[J].
Horticultural Plant Journal, 2021,7(5):367-379.
[41] Fang L, Ma Z, Wang Q, et al. Plant growth and
photosynthetic characteristics of soybean seedlings
under different led lighting quality conditions[J].Journal
of Plant Growth Regulation, 2020,40(2):668-678.
