黃河三角洲第三茬苜蓿主要節肢動物種群動態和群落特征研究
2024-01-02 00:00:00楊美艷宋夢昭高學民王婷鄭方強
山東農業大學學報(自然科學版) 2024年6期

摘要: 于2022 年對山東省黃河三角洲農業高新技術產業示范區第三茬苜蓿田主要節肢動物種類和數量進行了系統調查,分析了主要害蟲及其天敵的種群動態和節肢動物群落特征。第三茬苜蓿上的主要節肢動物有2 綱、11 目、31 科、50 種,其中害蟲29 種,相對多度最高的為薊馬類和煙粉虱;天敵21 種,相對多度最高的為食蚜蠅類和螟蛉盤絨繭蜂。害蟲優勢種有花薊馬、西花薊馬、鱗翅目幼蟲、綠盲蝽、小綠葉蟬和煙粉虱;主要天敵優勢種為煙蚜繭蜂、螟蛉盤絨繭蜂、微小花蝽、三突伊氏蛛和八斑鞘腹蛛。分析了苜蓿主要節肢動物群落特征指數,結果表明,苜蓿的生育期、氣候條件與苜蓿周邊植物上的節肢動物類群是影響種群動態和群落結構特征變化的主要因素。利用最優分割法將第三茬苜蓿主要節肢動物群落的時間格局劃分為3 個時間段:8 月14 日—9 月18 日,9 月19 日—9 月30 日,10 月1 日—10 月31 日。結合生育期和環境條件等因素,分析了這3 個時間段內苜蓿主要節肢動物的發生特點。研究結果為第三茬苜蓿害蟲的防治及天敵的保護利用提供了科學理論依據。
關鍵詞: 第三茬苜蓿;節肢動物;種群動態;群落特征;時間格局
中圖法分類號: S435.4 文獻標識碼: A 文章編號: 1000-2324(2024)06-0925-08
黃河三角洲地區鹽堿地面積大,嚴重制約了其農業的可持續發展。種植既耐鹽堿又有較高經濟價值的作物成為了本地農業可持續發展不可或缺的一部分[1]。苜蓿(Medicago sativa)是豆科植物,具有很好的固氮作用,在我國現代用于牧草和鹽堿地改良已有近80 年的歷史。苜蓿不僅營養豐富,還可優化土壤結構、防止土壤沙漠化和維護生態平衡[2,3]。由于其極高的經濟和生態價值,苜蓿還有“牧草之王”的美譽[4],隨著山東省“糧改飼”項目的推進,開始集中大規模種植苜蓿[5]。而在苜蓿大規模種植的同時,多年連作、粗放管理等問題更為嚴重,也給害蟲的發生創造了有利條件[6]。
近年來,有關苜蓿害蟲及其天敵的物種組成、群落特征以及害蟲的生態防控等方面已有較多報道。周軍等[7]調查了甘肅苜蓿種子田害蟲及天敵的種類,分析了不同類群亞群落的特征,通過時間格局分析明確了苜蓿種子田不同時期害蟲的主要防控對象及防治措施。郭艷春等[8]調查了鄭州黃河灘區紫花苜蓿害蟲的發生動態,主要有棉大造橋蟲(Ascotis selenaria)、盲蝽、甜菜夜蛾(Spodoptera exigua)、蚜蟲、薊馬、斑潛蠅和棉鈴蟲(Helicoverpa armigera)。在寧夏,朱猛蒙等[9]發現苜蓿主要害蟲有蝗蟲類、薊馬類、苜蓿象甲(Hypera postica)、苜蓿斑蚜(Therioaphis trifolii)和苜蓿盲蝽(Adelphocorislineolatus)。韓鳳英等[10]對山東陽谷縣苜蓿主要害蟲種群動態進行了研究,并提出了相應的防治技術,但未對害蟲與天敵的關系進行分析。王建麗等[11]研究發現黑龍江省苜蓿田優勢害蟲有蚜蟲、盲蝽、大青葉蟬(Cicadella viridis)、牛角花齒薊馬(Odontothrips loti)和黃脛小車蝗(Oedaleus infernalis),優勢天敵有龜紋瓢蟲(Propylaea japonica)、蜘蛛和食蚜蠅等。陳佳杰[12]調查河北的苜蓿害蟲發現,害蟲優勢種為棉鈴蟲、苜蓿斑蚜和牛角花齒薊馬。在美國,Randa 等[13]發現美國苜蓿田的主要害蟲有蚜蟲、蝗蟲和苜蓿象甲,其中為害最為嚴重的害蟲是苜蓿象甲;Makenzie 等[14]還研究了苜蓿象甲的生態學和治理技術。有關鹽堿地環境下苜蓿害蟲及天敵種類及種群動態和群落特征的研究則鮮有報道。
苜蓿作為重要的家畜飼料,對安全生產的要求更高,在其害蟲的防治中更加強調生態調控手段[15]。只有明確苜蓿生育期各個時段主要害蟲及其天敵的發生特點,才能制定科學精準的害蟲綠色防控技術。本研究通過對山東省黃河三角洲農業高新技術產業示范區第三茬苜蓿主要節肢動物種群動態和群落特征進行分析,旨在明確苜蓿節肢動物群落的物種組成、多樣性以及主要害蟲與天敵之間的關系,更科學地制定苜蓿害蟲的綠色防控規程。
1 材料與方法
1.1 試驗地概況
本試驗基地選在山東省東營市廣饒縣鹽堿地生物育種與現代農業試驗示范基地(37°17′N,118°37′ E)。苜蓿取樣地面積為1 333.3 m2,周圍種植有小麥(Triticum aestivum)、田菁(Sesbaniacannabina)、馬鈴薯(Solanum tuberosum)、藜麥(Chenopodium quinoa)、玉米(Zea mays)和大豆(Glycine max)等。樣地內未進行有害生物化學防治措施,僅進行常規的肥水管理。
1.2 調查方法
在取樣地采用棋盤式10 點取樣法,每樣點面積為9 m(2 3 m×3 m)。依不同害蟲生活習性和為害特點進行調查,每樣點隨機選取20 個苜蓿枝條。鱗翅目害蟲和各類天敵等采用目測法;薊馬、蚜蟲及具有假死性的節肢動物拍打震落法,拍盤為白搪瓷盤(長×寬=35×24 cm),地面節肢動物采用陷阱法(埋入土中塑料瓶的直徑為10 cm,高為15 cm);飛行能力強且隱蔽性強的節肢動物調查采取掃網法(網口直徑35 cm,桿長90 cm),記錄總數。第三茬苜蓿于8 月初再生,當其長至10 cm 高時開始調查,調查時間為2022 年8 月14 日至10 月31 日,共調查12 次,每7 d 調查1 次,集中在晴天上午完成,若遇到不利天氣等因素,可適當調整調查時間。
1.3 數據分析
(1)種群動態:節肢動物經分類統計后,使用Microsoft Excel繪制主要害蟲及天敵的種群動態曲線,以橫坐標表示調查時間,縱坐標表示調查當日的害蟲及其天敵的總數量。
(2)群落特征分析:種群優勢度用Berger-Parker 優勢度指數[16] (d)表示,種群生態優勢度以Simpson集中性指數[17(] C)表示,群落多樣性指數分別以Simpson 多樣性指數[17](D)和Shannon-Wiener指數[18(] H')表示,群落均勻度用Pielou均勻度指數[19(] J)計算。以上計算公式也參閱趙志模和郭依泉[20]、馬克平等[21]、丁巖欽[22]、Tokeshi[23]、Magurran[24]的文獻。
(3)時間格局分析:以每次調查時間(T)作為群落實體,以苜蓿重要害蟲種(類)數量(N1)及其天敵種(類)數量(N2)為群落屬性指標,構成T×(N1+N2)矩陣,利用最優分割法分析第3 茬苜蓿節肢動物群落的時間格局。
本文中主要群落特征指數和時間格局分析均利用DPS數據處理系統(V7.05)[25]進行處理。
2 結果與分析
2.1 第三茬苜蓿主要節肢動物物種組成及相對多度
從表1 看出,第三茬苜蓿主要節肢動物有50 種,隸屬于2 綱、11 目、31 科。其中害蟲共有29 種,隸屬于2 綱、7 目、17 科;天敵共有21 種,隸屬于2 綱、8 目、15 科。從相對多度來看,害蟲以刺吸式口器的薊馬科(54.72%)、煙粉虱(19.77%)和葉蟬科(11.93%)數量居多。捕食性類群中,食蚜蠅(0.87%)、蜘蛛類(0.82%)和微小花蝽(0.52%)占比較大。寄生性天敵類群主要為蚜繭蜂類(0.65%)和寄生鱗翅目幼蟲的螟蛉盤絨繭蜂(0.94%),共占1.59%。
2.2 第三茬苜蓿田主要害蟲及天敵的種群動態分析
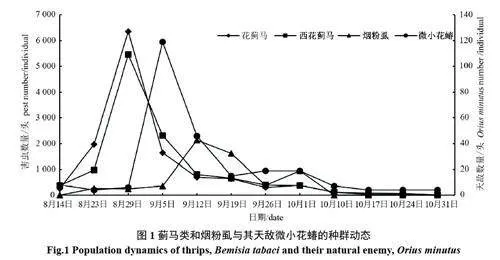
2.2.1 薊馬類和煙粉虱與其天敵微小花蝽種群動態分析 圖1 可知,兩種薊馬的發生動態明顯地呈現前期單峰型增長模式,二者發生量與發生動態較為相似。從第三茬苜蓿田返青以來,薊馬種群數量上升迅速,至8 月29 日達到最大值(花薊馬6 361 頭,西花薊馬5 455 頭),此后隨著微小花蝽種群數量的上升,薊馬的種群數量逐漸下降。煙粉虱在第三茬苜蓿生長的中期多發,在9 月5 日之前種群數量較低且平穩,之后急劇上升,至9 月12 日達到高峰期(2 137 頭),整個9 月中旬持續大量發生,10 月1 日出現1 次小高峰,基本符合煙粉虱偏高溫季節發生重的規律。微小花蝽從8 月23 日到調查結束均有發生,其發生明顯地與薊馬類和煙粉虱相吻合,有較好的跟隨現象,對薊馬和煙粉虱均有良好的控制作用。
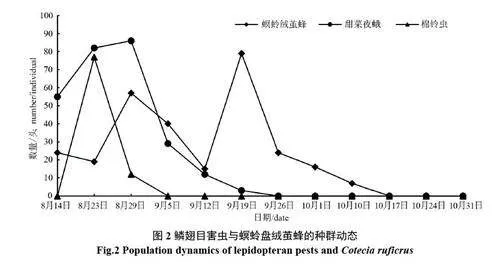
2.2.2 鱗翅目害蟲與螟蛉盤絨繭蜂種群動態分析 鱗翅目害蟲(主要是甜菜夜蛾和棉鈴蟲)的發生動態明顯地呈前期單峰型(圖2),主要發生在第三茬苜蓿分枝期,此時葉片幼嫩,為害造成的“開天窗”和“缺刻”相對嚴重。甜菜夜蛾和棉鈴蟲自8 月14 日苜蓿返青后,數量均呈上升趨勢,其中,甜菜夜蛾在8 月29 日達到最高峰,棉鈴蟲在8 月23日達到最高峰。達到高峰之后二者數量均開始下降,主要是由于螟蛉盤絨繭蜂的控制作用所致。螟蛉盤絨繭蜂分別在8月29日(61頭)與9月5日、9月19日數量(79頭)較大,形成兩個高峰期,特別是第一個高峰期明顯地與鱗翅目幼蟲發生相吻合,對甜菜夜蛾和棉鈴蟲均具有良好的控制作用。
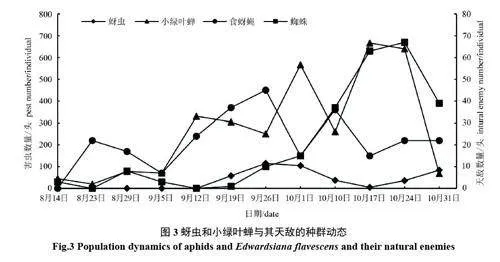
2.2.3 蚜蟲和小綠葉蟬與其天敵的種群動態分析 8 月中旬到9 月上旬的高溫季節,蚜蟲數量很少。至9 月中旬后隨著氣溫略降,蚜蟲數量逐漸增多,至9 月26 日達到高峰,此后由于天敵食蚜蠅、蚜繭蜂等天敵的控制作用以及10 月3 日的暴雨,蚜量急劇下降,10 月17 日后豌豆蚜數量又明顯回升,至調查結束數量達到第2 個高峰。小綠葉蟬種群數量在9 月5 日急劇上升,分別在10 月1 日、10 月中下旬出現兩次高峰,10 月10 日出現一個低谷是由于蜘蛛的自然控制作用和10 月3 日的暴雨所致。小綠葉蟬在發生動態上總體上呈現后期多發型(10 月份)。食蚜蠅類種群數量出現兩次高峰,第1 次在8 月23 日,主要與三葉草彩斑蚜發生相關;第2 次主要與豌豆蚜發生相吻合,說明食蚜蠅對蚜蟲有良好的跟隨效應。蜘蛛類則與蚜蟲類、小綠葉蟬的跟隨關系顯著(圖3)。
2.3 第三茬苜蓿主要節肢動物優勢度與優勢種
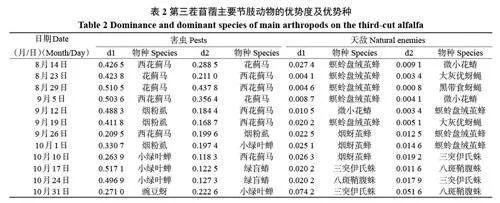
依據不同時間段物種的優勢度大小判定優勢種(表2)。苜蓿害蟲的優勢種主要為西花薊馬、花薊馬、小綠葉蟬、煙粉虱和綠盲蝽。自8 月中旬至9 月初,花薊馬和西花薊馬保持種群最大優勢度,是主要害蟲優勢種,花薊馬在8 月29 日達到優勢度最大值0.510 5,西花薊馬在9 月5 日達到優勢度最大值0.503 6。9 月中旬至10 月初煙粉虱的數量上升,成為優勢種,在9 月12 日達到最大優勢度0.488 3。10 月上旬至10 月底小綠葉蟬、綠盲蝽數量上升,成為優勢種,其中小綠葉蟬在10月17日達到最大優勢度0.517 1。
自然天敵的主要優勢種有螟蛉盤絨繭蜂、微小花蝽、煙蚜繭蜂、食蚜蠅類和蜘蛛類,8 月中旬至9 月中旬主要優勢天敵是鱗翅目幼蟲寄生蜂螟蛉盤絨繭蜂,在8 月14 日達到最大值0.024 7;9 月12 日微小花蝽優勢度較高,食蚜蠅類也有較高的優勢度。9 月中旬至10 月初煙蚜繭蜂與大灰優蚜蠅、螟蛉盤絨繭蜂交替成為優勢種。10 月中下旬氣溫逐漸降低,蜘蛛類天敵成為優勢種。
2.4 第三茬苜蓿主要節肢動物群落特征分析
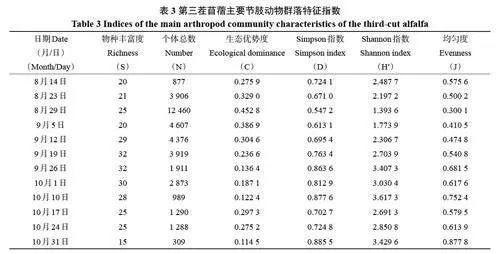
第三茬苜蓿節肢動物群落特征指數見表3。物種豐富度在9 月中下旬達到最大值,是因為苜蓿田周圍玉米等作物臨近收獲,一些薊馬類、葉蟬類、食蚜蠅、蜘蛛類等節肢動物遷移到苜蓿田的結果。群落個體總數量在8 月下旬至和9 月中旬均高,主要是薊馬類和煙粉虱優勢度很高所致,此時物種豐富度較低,生態優勢度較高,群落多樣性指數(D和H')較低。9 月中旬之后,隨著氣溫下降,主要害蟲煙粉虱、薊馬類的優勢度降低以及多種天敵遷移到苜蓿田繁殖,物種豐富度較高,生態優勢度明顯下降,從而群落多樣性和均勻度明顯提高。至10 月下旬,隨著氣溫降低,苜蓿臨近收獲,節肢動物總體種類和數量明顯下降,生態優勢度降至最小,群落多樣性和均勻度均達最高。
2.5 第三茬苜蓿主要節肢動物群落時間格局分析
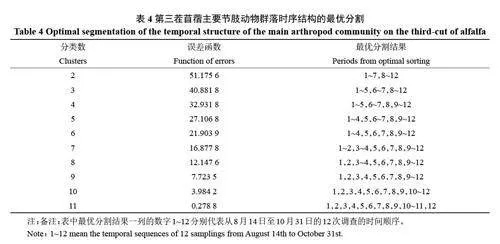
根據表4 可知,誤差函數值在最優分割數為3 之前下降幅度較大,之后幅度變緩。因此,將苜蓿害蟲及其天敵群落的發展階段分為3 個時段最佳。
第1 時段(1~5 次)8 月14 日至9 月18 日,第3茬苜蓿處于分枝期,葉片生長發育快,鱗翅目幼蟲(棉鈴蟲、甜菜夜蛾等)發生為害重,薊馬類害蟲種群數量也較多。天敵主要有寄生鱗翅目幼蟲的螟蛉盤絨繭蜂及捕食薊馬的微小花蝽,由于薊馬類種群數量甚高,導致群落豐富度與群落多樣性都較低。
第2 時段(6~7 次)9 月19 日至9 月30 日,這段時間隨著氣溫下降,雨水也少,苜蓿進入現蕾期,加上天敵作用,鱗翅目幼蟲、煙粉虱和西花薊馬數量均明顯減少。小綠葉蟬和蚜蟲類數量明顯上升,天敵煙蚜繭蜂、大灰優蚜蠅和微小花蝽數量明顯增多。群落多樣性和均勻度明顯提高。
第3 時段(8~12 次)10 月1 日至10 月31 日,此時氣溫下降快,苜蓿進入開花期,加上苜蓿田周圍玉米等作物臨近收獲,一些植食性蝽和小綠葉蟬遷移到苜蓿田導致種群數量略有增加。10 月中旬以后蚜蟲種群數量明顯上升,其天敵蚜繭蜂、食蚜蠅、微小花蝽和蜘蛛類數量增加。再加上薊馬類和煙粉虱的數量顯著減少,群落多樣性和均勻度明顯提高。
3 討論
3.1 第三茬苜蓿主要害蟲與天敵的跟隨效應
8 月初至9 月上旬,高溫少雨(2022 年整個8 月份降雨偏少)導致適合這個季節發生的害蟲薊馬、煙粉虱發生量大,為害重,故在9 月中旬之前,刺吸類害蟲薊馬類和煙粉虱的相對多度和優勢度一直很高(表2);食葉鱗翅目害蟲的相對多度也較高,主要是甜菜夜蛾的幼蟲發生量較大。這些害蟲直接影響葉片的光合作用和第三茬苜蓿的早期生長發育。所以,自8 月14 日至9 月中旬,微小花蝽與薊馬和粉虱的跟隨關系、螟蛉盤絨繭蜂與鱗翅目幼蟲的跟隨關系都較緊密。9 月中旬之后,隨著氣溫下降,適合較低溫度環境發生的蚜蟲種群上升,同時,其天敵食蚜蠅數量也隨之上升。總之,第三茬苜蓿上主要害蟲與相應的天敵跟隨效應十分明顯,且自然控制作用較強,但由于害蟲的數量很大,這些天敵有時難以將這些害蟲完全控制住。因此,在防治這些害蟲時,可通過使用一些低毒或微毒生理選擇性殺蟲劑或生物殺蟲劑來防治,盡量保護自然天敵,讓天敵的自然控制作用發揮得更好。
3.2 關于第三茬苜蓿主要害蟲與天敵的優勢種判斷
本研究中優勢種的判斷主要基于害蟲及天敵的種群數量,從不同時間的調查結果來看,在9 月之前,苜蓿的優勢種害蟲有西花薊馬、花薊馬、煙粉虱,對應的主要天敵則是微小花蝽;10 月的害蟲優勢種是小綠葉蟬、綠盲蝽和豌豆蚜,對應的天敵優勢種是蜘蛛類和煙蚜繭蜂,蜘蛛類能捕食多種害蟲。其實,在第三茬苜蓿分枝期,食葉類鱗翅目害蟲數量較大,對苜蓿的為害更大,盡管在數量上無法和許多刺吸類害蟲相比,但從危害性看,也應是優勢種。在鱗翅目害蟲發生的同時,其寄生性天敵螟蛉盤絨繭蜂跟隨現象明顯,發生量較大,從8 月中旬到10 月初一直是優勢種。所以,對優勢種的判定不能僅以物種個體數量來判斷,還應根據其在生態系統中的作用等方面來判斷,以提高優勢種判定的客觀性[20,22],這樣才能更好地去控制害蟲。
3.3 第三茬苜蓿主要節肢動物群落的時間格局對苜蓿害蟲防治的意義
有序樣本最優分割是在不打破時間順序的基礎上,對節肢動物群落時間格局進行劃分的方法[20,25]。研究昆蟲群落的時間格局,對于掌握各時期內害蟲及天敵的發生特點、防治或保護具有重要理論和實踐意義[26]。
第1 時段(8 月14 日至9 月18 日),此時段苜蓿處于分枝期,主要害蟲為薊馬類和鱗翅目類,自然天敵(微小花蝽、螟蛉盤絨繭蜂等)也很豐富,有很好的自然控制作用。防治這些害蟲盡量用生物殺蟲劑(防治薊馬類可用乙基多殺霉素,防治鱗翅目幼蟲用昆蟲病毒、蘇云金芽孢桿菌等)和選擇性微毒或低毒級化學殺蟲劑(如氯蟲苯甲酰胺或滅幼脲類)以保護利用天敵。對鱗翅目害蟲(甜菜夜蛾、棉鈴蟲等)還可以用頻振殺蟲燈或性誘劑誘殺成蟲。Tharp 等指出,對苜蓿害蟲防治要用環境更友好、目標更明確的替代性控制方法,如使用昆蟲生長調節劑氟酰脲[27]。
第2 時段(9 月19 日至9 月30 日),這段時間苜蓿進入現蕾期,主要害蟲是煙粉虱、小綠葉蟬、薊馬類和豌豆蚜。這些害蟲盡管有豐富的天敵,但難以被控制住。因此,可考慮用選擇性的新煙堿類殺蟲劑進行防治。
第3 個時段(10 月1 日至10 月31 日),除了小綠葉蟬和蚜蟲類發生量較高外,其他類害蟲數量均較低,天敵主要是蜘蛛類、食蚜蠅類和微小花蝽。考慮到這一時段害蟲數量較低和有較豐富的天敵,可以不用化學防治措施,讓天敵充分發揮自然控制作用,也避免了苜蓿采收期殺蟲劑的殘留。
參考文獻
[1] 韓 瑩. 加快推進苜蓿產業發展的建議[J]. 江西農
業, 2020,(18):140-144.
[2] 陳 頎,張林源. 苜蓿的開發與利用[J]. 科技創新導
報,2009,(7):111-112.
[3] 王文信. 中國苜蓿種植業發展的對策[J]. 北京農學
院學報,2022,37(1):117-120.
[4] 李競前,閆奎友,柳珍英,等. 我國優質高產苜蓿發展
狀況及對策建議[J]. 中國飼料,2021,(11):95-98.
[5] 張進紅,劉名江,吳 波,等. 50 個紫花苜蓿品種在黃
河三角洲鹽堿地區的生產性能評價[J]. 農學學報,
2019,9(2):64-71.
[6] 熊 兵. 紫花苜蓿的病蟲害防治及飼用價值分析[J].
中國畜禽種業, 2022, 18(5):51-52.
[7] 周 軍. 景泰苜蓿種子田節肢動物群落結構及多樣
性研究[D]. 甘肅農業大學碩士論文,2005.
[8] 郭艷春,高永革. 鄭州黃河灘區紫花苜蓿昆蟲發生
動態調查[J]. 河南農業科學,2010,(9):99-101.
[9] 朱猛蒙,李東寧,張 蓉,等. 不同種植年限苜蓿草地
昆蟲種群及群落的發生規律[J]. 寧夏農林科技,
2014,55(1):48-53.
[10] 韓鳳英,徐金強,秦 旭,等. 山東苜蓿地主要害蟲種
群的消長規律[J]. 湖北農業科學,2014,53(9):2058-
2060.
[11] 王建麗,申忠寶,潘多鋒,等. 黑龍江蘭西縣苜蓿害蟲
及天敵種類調查初報[J]. 黑龍江農業科學,2014,(5):
67-69.
[12] 陳佳杰. 河北滄州地區苜蓿田昆蟲群落動態及主要
害蟲防控技術[D]. 河北農業大學碩士學位論文,
2020.
[13] Randa J, Shiri N. Wyoming producer priorities and
perceptions of alfalfa insect pests[J]. Journal of
Integrated Pest Management, 2017, 8: 1-5.
[14] Makenzie E, Pellissier Z N, Randa J. Ecology and
management of the alfalfa weevil (Coleoptera:
Curculionidae) in Western United States alfalfa[J].
Journal of Integrated Pest Management, 2017, 8: 1-7.
[15] 楊偉光,柴 華,黃新育,等. 黑龍江省齊齊哈爾地區
紫花苜蓿主要蟲害及其發生規律[J]. 飼料博覽,
2013,(12):26-30.
[16] Berger W H, Parker F L. Diversity of planktonic
Foraminifera in deep sea sediments[J]. Science,
1970, 168(3937): 1345-1347.
[17] Simpson E H. Measuerment of diversity[J]. Nature,
1949, 163(688): 631-648.
[18] Shannon C E, Weaver W. The mathematical theory
of communication[M]. Urbana: University of Illinios
Press, 1949.
[19] Pielou E C. Ecological diversity[M]. Newyork:
Wiley-interscience, 1975.
[20] 趙志模,郭依泉. 群落生態學原理與方法[M]. 重慶:
科學技術文獻出版社重慶分社,1990.
[21] 馬克平,劉玉明. 生物群落多樣性的測度方法Ⅰα 多
樣性的測度方法( 下) [J]. 生物多樣性, 1994, (4):
231-239.
[22] 丁巖欽. 昆蟲數學生態學[M]. 北京:科學出版社,
1994.
[23] Tokeshi M. Species abundance patterns and community
structure[J]. Advances in Ecological Research, 1993, 24:
111-186.
[24] Magurran A E. 張峰主譯. 生物多樣性測度[M]. 北
京:科學出版社,2011.
[25] 唐啟義.DPS 數據處理系統:實驗設計、統計分析
及數據挖掘[M]. 北京:科學出版社,2010.
[26] 鄭方強,張曉華,曲誠懷,等. 紅富士蘋果園害蟲與天
敵群落的定量分析[J]. 應用生態學報,2009,20(4):
851-856.
[27] Tharp C I, Blodgett S L, Kephart K D. Susceptibility
of alfalfa insect pests and predators to mustang max,
warrior and rimon, 2005[J]. Arthropod Management
Tests, 2006, 31: 1-2.
