單一及復合喹諾酮類抗生素脅迫下留蘭香萌發生長的響應及耐受性分析
2024-01-17 00:00:00張解和姜佳慧楊怡章凱帆吳彤彤陶海升
農業環境科學學報 2024年12期



關鍵詞:喹諾酮類抗生素;留蘭香;響應;耐受性;隸屬函數
中圖分類號:S682.19;X592;X173 文獻標志碼:A 文章編號:1672-2043(2024)12-2876-13 doi:10.11654/jaes.2024-0537
抗生素可用于臨床上治療相關疾病,也可作為生長促進劑添加至飼料中。喹諾酮類(Quinolones,QNs)抗生素是臨床上使用的第二大類抗菌藥物,同時也是常用獸用抗生素之一,其中環丙沙星(Cipro?floxacin,CIP)、諾氟沙星(Norfloxacin,NOR)和氧氟沙星(Ofloxacin,OFL)是最常用的3種。生物體攝入的QNs抗生素中有高達70%不能被代謝而排出體外[1],并隨醫療及城市生活污水、制藥企業和農牧業廢水等排放至自然水體中。在世界各國的污水廠中,QNs抗生素也是檢出頻率前五的污染物之一。CIP、OFL、NOR 在各國污水廠中的檢出頻率較高,同時也是淡水及海洋環境最常檢出的QNs抗生素[2]。非洲地區廢水中QNs 抗生素的檢測濃度達50 μg·L-1,我國渤海萊州半島、貴州和大連等地檢測到其濃度達μg·L-1數量級,大多數土壤和沉積物環境介質中的含量為μg·kg-1數量級,其中,OFL、CIP 和NOR 在水體和沉積物中最常被檢測到[3-4]。QNs抗生素對生態環境的潛在危害[5]已成為環境領域廣泛關注的熱點問題。
QNs抗生素對植物種子萌發和幼苗生長具有顯著的毒性效應。采用恒溫保濕培養法研究抗生素對油菜(Brassica campestris)萌發的影響時發現,濃度為12.5、25、50、100、200 mg·L-1的NOR處理均抑制了油菜種子萌發,200 mg·L-1處理時種子發芽率和發芽勢相較于對照組均下降了13%,根長抑制率達60%[6]。實驗證明CIP 濃度超過1 mg·L-1 時,黑麥草(Loliumperenne)種子的發芽率低于對照組,發芽勢和發芽指數相對于對照組分別下降50%和30%[7]。類似地,在0.1、0.5、1.0、5.0、10.0、50.0 mg·L-1 OFL脅迫紫花苜蓿(Medicago sativa)種子萌發的毒性試驗中,處理組的發芽率均低于對照組[8]。QNs 抗生素進入農田系統后,也會影響農作物的生長和繁殖。研究表明,0~2mg·L-1 CIP脅迫下,玉米(Zea mays)的H2O2水平和抗氧化酶活性顯著提高,形態生理改變,種子產量降低[9]。2 mg·L-1的OFL處理能夠抑制洋蔥(Allium fistu?losum)的光合作用,破壞葉細胞的超微結構,使葉片黃化[10]。韭菜(Allium tuberosum)在NOR 含量為10mg·kg-1和100 mg·kg-1的土壤中培養時,其葉片中細胞形狀相對于對照組都發生了明顯變化,類囊體在低密度區域膨脹,淀粉粒數量明顯增加,葉片和根中富集的抗生素含量可達0.88 mg·kg-1和0.22 mg·kg-1[11]。QNs抗生素單一及復合脅迫的毒性也不同,CIP、恩諾沙星、左氧氟沙星及其復合物對小麥(Triticum aesti?vum)的短期(20 d)毒性研究發現,高濃度抗生素復合物(100 mg·L-1和300 mg·L-1)相較于單一脅迫,會顯著提高脂質過氧化物含量及抗氧化酶活性,抑制小麥幼苗的生長[12]。可見,QNs抗生素能抑制植物種子的萌發和幼苗生長,且單一及復合抗生素對植物的毒性效應存在差異。QNs抗生素在農村生活污水處理廠進水中的檢出濃度普遍較高(可達65 μg·L-1)[13]。對我國各地區的水、土壤和沉積物中抗生素進行檢測發現,QNs抗生素的檢測濃度可達數十微克每升或每千克[14],且環境中QNs抗生素的存在形式并非某一特定種類,而往往是多種抗生素復合存在,隨著QNs抗生素的大量使用,有必要開展mg·L-1數量級水平的QNs抗生素單一及復合污染的生態毒性研究。
留蘭香(Mentha spicata L.)為隸屬于唇形科薄荷屬的多年生芳香草本植物,廣泛用于室內觀賞和城市綠化,具有極高的藥用、食用及經濟價值[15]。留蘭香原產非洲西北部群島和歐洲南部區域,在我國主要栽培于江蘇、安徽、四川、貴州等地區[16]。留蘭香喜濕潤,其生長初期和中期對水分的需求量較大[17],因此,灌溉用水及土壤中的QNs抗生素會影響其正常生長。污染物進入環境后,不僅能影響植物正常生長,還會嚴重影響芳香草本植物的產量和品質[18-19],因此,評估QNs抗生素的生態毒性,對留蘭香生態種植的順利開展具有指導意義。目前,對留蘭香的生態毒性研究主要圍繞溫度、水分等自然因素,以及鹽脅迫對留蘭香植株生長及品質的影響[20-21],也有研究表明土霉素能夠影響留蘭香莖的生長、降低葉綠素的含量[22],但QNs抗生素單一及復合脅迫對留蘭香種子萌發及幼苗生長的影響及植物耐受情況尚不明確。因此,有必要就抗生素對留蘭香的生態毒性效應做進一步研究。綜上,本文研究典型QNs抗生素CIP、NOR、OFL對留蘭香種子萌發指標和生長特性的影響,旨在為抗生素的生態風險評價提供基礎數據。
1 材料與方法
1.1 材料
試驗所用發育正常的留蘭香(Mentha spicata L.)種子(千粒質量約為0.08 g)由宿遷綠芃種業科技有限公司提供,于2023年10月采自甘肅酒泉,暫存于冰箱中4 ℃低溫保存。CIP、NOR和OFL來自上海麥克林生化科技股份有限公司,純度為99%。
1.2 種子萌發試驗
挑選大小均一且飽滿的留蘭香種子,經75% 乙醇消毒15 min后,用無菌水清洗3~5次去除乙醇,取出并吸干其多余的水分,備用。根據預試驗中留蘭香種子對QNs抗生素的耐受濃度并結合文獻[23-25],配制CIP、NOR、OFL單一及三者等質量復合溶液,濃度為0.1、1、5、10、20 mg·L-1,以無菌蒸餾水為對照(CK)。參考國際種子檢測協會(ISTA)的種子監測標準,采用保溫保濕培育法進行試驗。取墊有2層濾紙的直徑9 cm培養皿,用鑷子將50粒留蘭香種子均勻地排列在培養皿中,分別加入5 mL不同濃度的CIP、NOR、OFL及復合溶液,每個處理重復3次,將培養皿置于恒溫培養箱,設置溫度為(25±1)℃、光周期為12 h光照/12 h黑暗,光照強度為2 000 lx。發芽期間每隔2 d更換一次濾紙、沖洗培養皿,隨機改變留蘭香種子在培養皿中的位置,并添加抗生素溶液保持濕潤[26-28],及時剔除霉變的種子,觀察并記錄其發芽情況(以胚根長于種子1/2作為萌發標準)。CIP、NOR和OFL在培養過程中可能發生光降解,考慮實際抗生素濃度變化對種子萌發的影響,用250 mL錐形瓶盛裝試驗濃度的抗生素溶液置于培養箱中,培養中添加的溶液為同時放入培養箱內錐形瓶中的抗生素溶液。
1.3 分析方法
1.3.1 種子萌發指標的測定
在留蘭香萌發試驗中,將種子放入人工培養箱后的24 h為試驗第1天,以連續3 d種子發芽數不再增加視為發芽結束(本試驗為發芽14 d結束),根據每天發芽的種子數,計算種子的發芽百分比。計算培養結束時的日平均發芽率、平均發芽時間、發芽指數和活力指數等。日發芽種子數達到最高峰時作為發芽勢計算標準[29],CIP和NOR處理時以種子萌發的第4天、OFL和復合處理時以第5天的種子發芽百分比為發芽勢(GP)。
式中:n 指全部正常發芽的種子數;N 指供試種子總數;Gt 指第t 天發芽的種子數;Dt 指相應的發芽天數;RL 指胚根長;Gd 指總發芽天數。
脅迫指數為正值,表示處理組指標值小于對照組,該指標被抑制,反之表示對該指標起促進作用,其絕對值大小反映影響程度。可塑性指數反映植物對生境的適應能力,數值越大,表明適應能力越強[30-31]。
1.3.2 幼苗形態指標的測定
培養完成后取出萌發后的幼苗,用濾紙吸干其表面水分,用直尺測定胚根長和胚芽長,稱量其鮮質量,計算胚根長、胚芽長和鮮質量的抑制率。構建CIP、NOR、OFL和復合物的濃度與種子胚根長、胚芽長和鮮質量的抑制率的回歸方程,當指標的抑制率為50%時,對應的抗生素濃度即為該種抗生素對胚根長、胚芽長和鮮質量的半數抑制濃度(IC50)。
胚根長(RL)、胚芽長(GL)、鮮質量(FW)抑制率=(對照組-處理組)/對照組×100% (9)
1.3.3 留蘭香對QNs抗生素耐受性的評價
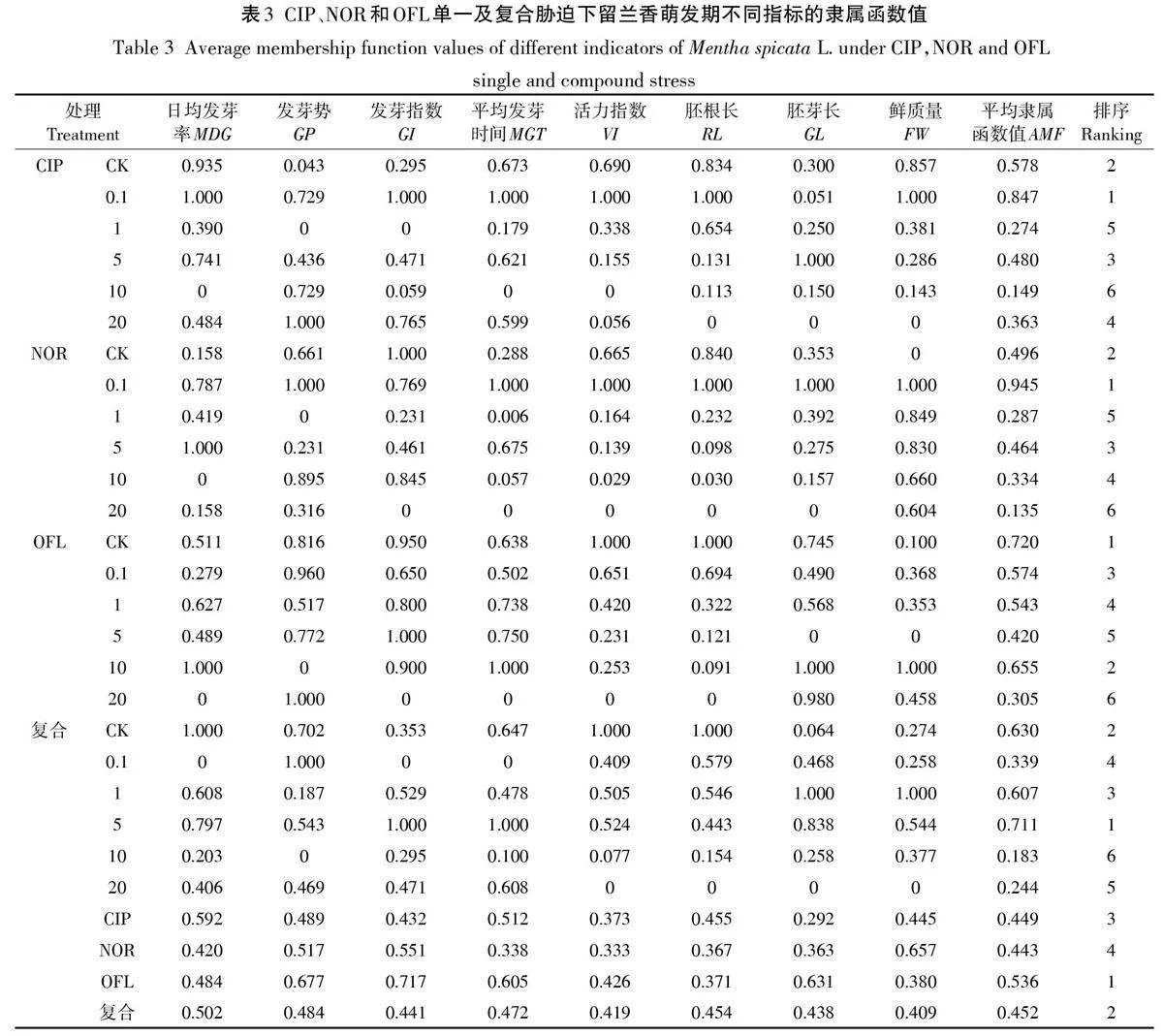
根據模糊數學原理,利用隸屬函數法對留蘭香試驗種子萌發和幼苗形態指標進行計算并排序,綜合評價QNs抗生素脅迫下種子的耐受性情況[26]。利用公式(10)和公式(11)[32-33]分別計算留蘭香各指標(發芽勢、日平均發芽率、平均發芽時間、發芽指數、活力指數、胚根長、胚芽長和鮮質量)在CIP、NOR和OFL單一及復合脅迫下的具體隸屬函數值μ(Xij),根據測定指標的隸屬函數值,計算該濃度下所有指數的平均隸屬函數值(AMF)。AMF 值越大表示種子在萌發期對QNs抗生素的綜合耐受性越強,反之更弱。按灰色系統理論要求[34],將留蘭香種子對抗生素的綜合耐受性和不同QNs抗生素脅迫下8個指標視為一個整體,即灰色系統。將綜合耐受性指數作為參考數列(母序列),各指標原始數據標準化處理后的值作為比較數列(子序列),建立灰色系統,計算各指標與其綜合耐受性指數的關聯度并排序(關聯序),指標的關聯度值越大,即表示該指標與綜合耐受性的關聯密切程度越高[35]。平均隸屬函數值和灰色關聯度計算公式如下:
1.4 數據分析
采用Excel 2016 對數據進行統計,SPSS 27 進行單因素方差分析(One-way ANOVA)和Pearson 相關性分析,利用LSD法進行多重比較,用Origin 2021擬合回歸曲線、作圖。使用SmartPLS 4.1對QNs抗生素對留蘭香萌發期指標的影響建立偏最小二乘結構方程模型(PLS-SEM)。
2 結果與分析
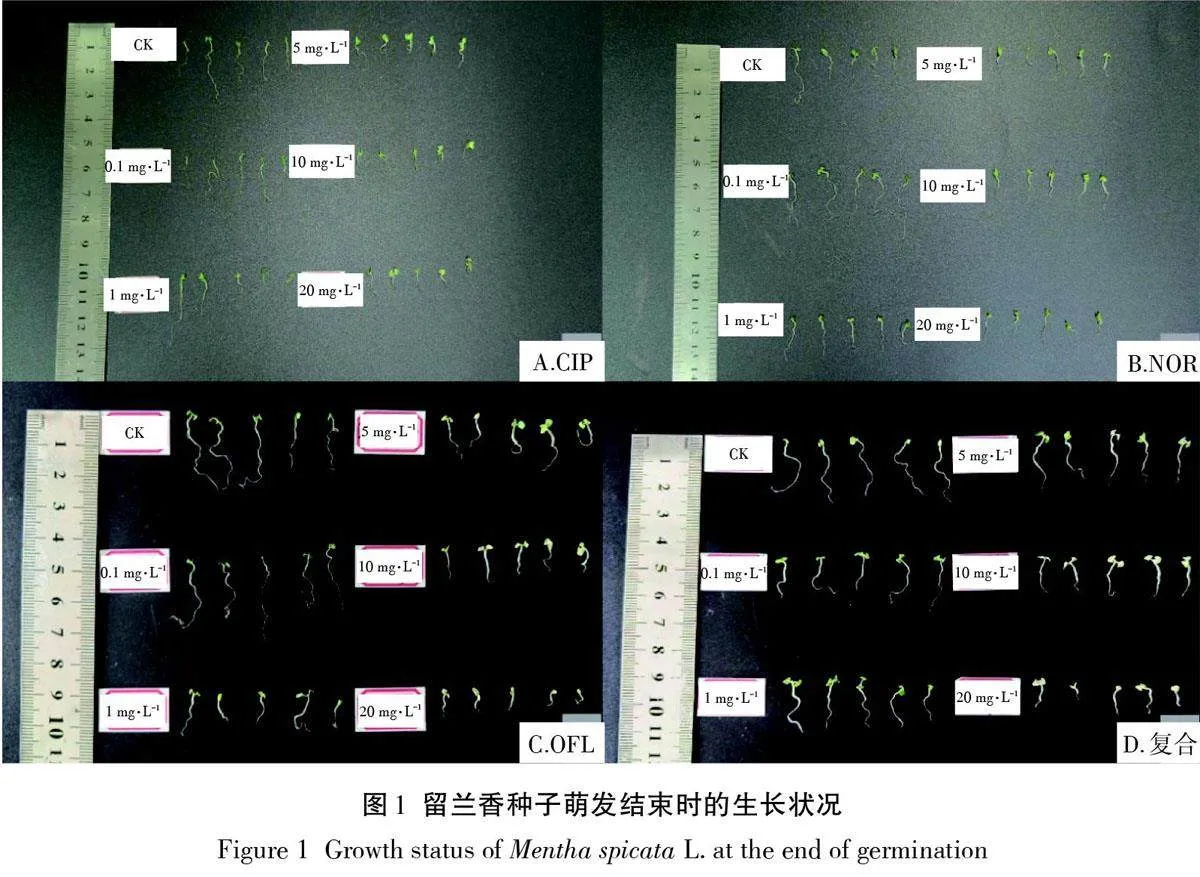
CIP、NOR和OFL單一及復合脅迫下留蘭香培養14 d后的生長狀況如圖1所示,各處理組中幼苗的生長情況差異明顯:OFL及復合脅迫下生長良好,幼根更粗;CIP和NOR處理組幼苗葉片面積小,葉發黃,幼根細,長勢不良。
2.1 單一及復合QNs抗生素對留蘭香種子發芽的影響
CIP、NOR和OFL單一及復合脅迫對留蘭香種子萌發的影響如圖2所示。各處理組中,留蘭香均在第2天發芽,說明單一及復合脅迫未改變種子初始萌發時間。QNs抗生素也不影響留蘭香萌發結束時間,第12天后留蘭香種子發芽數基本不再增加,但日發芽種子數達到最高峰的時間不同。CIP和NOR處理時第4天發芽種子數最多,OFL和復合處理時第5天的種子發芽數最多。
2.2 單一及復合QNs抗生素對留蘭香種子萌發指標的影響
由表1可知,高濃度的CIP和OFL能夠顯著抑制留蘭香種子的日平均發芽率(Plt;0.01),20 mg·L-1處理時脅迫指數分別為0.21和0.31;但10~20 mg·L-1 NOR和復合處理對日平均發芽率的抑制作用不顯著。所有萌發指標中,平均發芽時間的可塑性指數變化最大(0.040~0.642),即留蘭香的平均發芽時間對不同QNs的響應不同。0.1 mg·L-1 QNs處理時均能縮短平均發芽時間,20 mg·L-1 CIP和OFL能顯著縮短平均發芽時間,而20 mg·L-1 NOR和復合處理則顯著延長平均發芽時間(Plt;0.05)。高濃度QNs抗生素對留蘭香發芽勢的影響不同,0.1 mg·L-1 CIP能顯著提高留蘭香發芽勢(脅迫指數為-0.55,Plt;0.01),20 mg·L-1 NOR 和OFL處理極顯著降低發芽勢(脅迫指數均為0.59,Plt;0.01),但20 mg·L-1 CIP和復合處理時的發芽勢高于CK組。
低濃度的CIP和NOR處理能顯著提高留蘭香的發芽指數,0.1 mg·L-1處理時發芽指數最高(脅迫指數分別為-0.17和-0.20,Plt;0.01);20 mg·L-1脅迫下,除OFL顯著降低留蘭香發芽指數(脅迫指數為0.38)外,其他處理對留蘭香發芽指數均無顯著抑制作用。留蘭香的所有萌發指標中,活力指數的可塑性指數變化最小(范圍為0.732~0.863),說明留蘭香的活力指數對QNs 的耐受性差別不大。活力指數隨著CIP 和NOR濃度的增加先升后降,0.1 mg·L-1處理時發芽指數最高(脅迫指數分別為-0.32和-0.36,Plt;0.01);OFL和復合處理時,活力指數隨濃度的增加而逐漸降低,20 mg·L-1處理時活力指數的抑制程度最高(脅迫指數分別為0.86和0.73)。通過計算留蘭香萌發指標的可塑性指數比較各萌發指標的可塑性程度,萌發可塑性指數大小為OFLgt;CIPgt;復合gt;NOR。
2.3 單一及復合QNs抗生素對留蘭香幼苗生長的影響
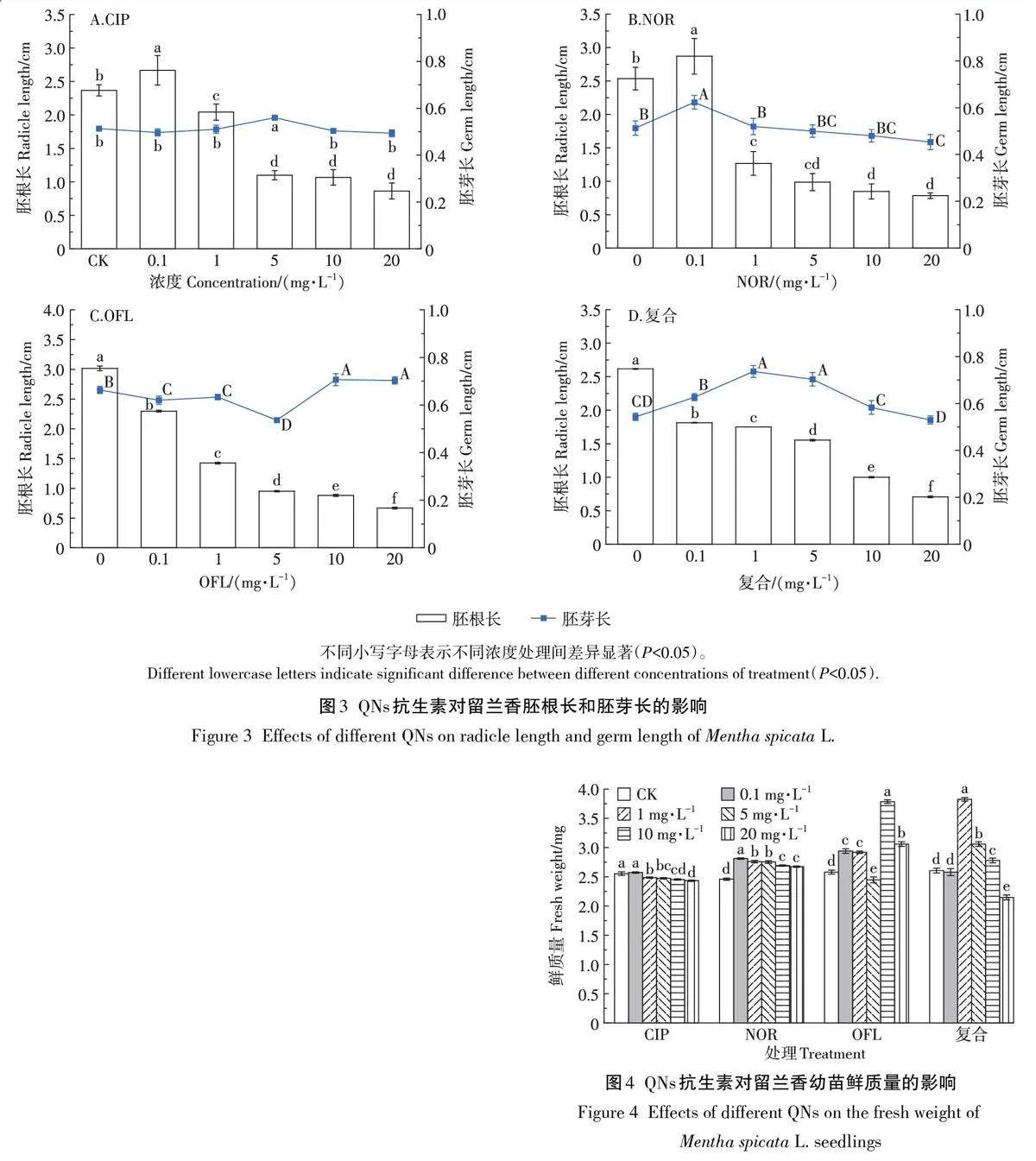
CIP和NOR對留蘭香胚根長和胚芽長的影響如圖3A 和圖3B 所示,0.1 mg·L-1處理均能促進留蘭香種子胚根的生長;較于CK 組,1~20 mg·L-1處理均能顯著抑制留蘭香種子胚根的生長(Plt;0.05),且濃度越高抑制作用越強。試驗濃度范圍內OFL和復合處理均能抑制留蘭香胚根的生長,且濃度越高抑制作用越大,20 mg·L-1 OFL和復合脅迫下胚根長較CK組分別下降了78%和73%(圖3C和圖3D)。不同濃度的QNs抗生素對留蘭香胚芽的作用不同。5 mg·L-1 CIP極顯著促進胚芽的生長(Plt;0.01),而其他濃度的CIP對留蘭香種子胚芽的生長均無顯著作用(Pgt;0.05);NOR對留蘭香種子胚芽呈低促高抑的效應,0.1 mg·L-1 NOR促進留蘭香種子胚芽的生長,隨濃度升高對胚芽的抑制作用增強;0.1~10 mg·L-1 復合處理時能促進留蘭香種子胚芽的生長,而20 mg·L-1 處理抑制了留蘭香種子胚芽的生長。0.1~5 mg·L-1 OFL能抑制留蘭香種子胚芽的生長,而10~20 mg·L-1 處理能促進留蘭香種子胚芽的生長。
如圖4所示,相較于CK組,0.1 mg·L-1 CIP和復合處理對留蘭香鮮質量無顯著抑制作用,而高濃度處理時能抑制留蘭香鮮質量,20 mg·L-1處理時鮮質量較CK極顯著降低(Plt;0.01)。NOR和OFL處理時鮮質量整體呈增加趨勢,且處理濃度分別為0.1 mg·L-1和10mg·L-1時鮮質量最大。
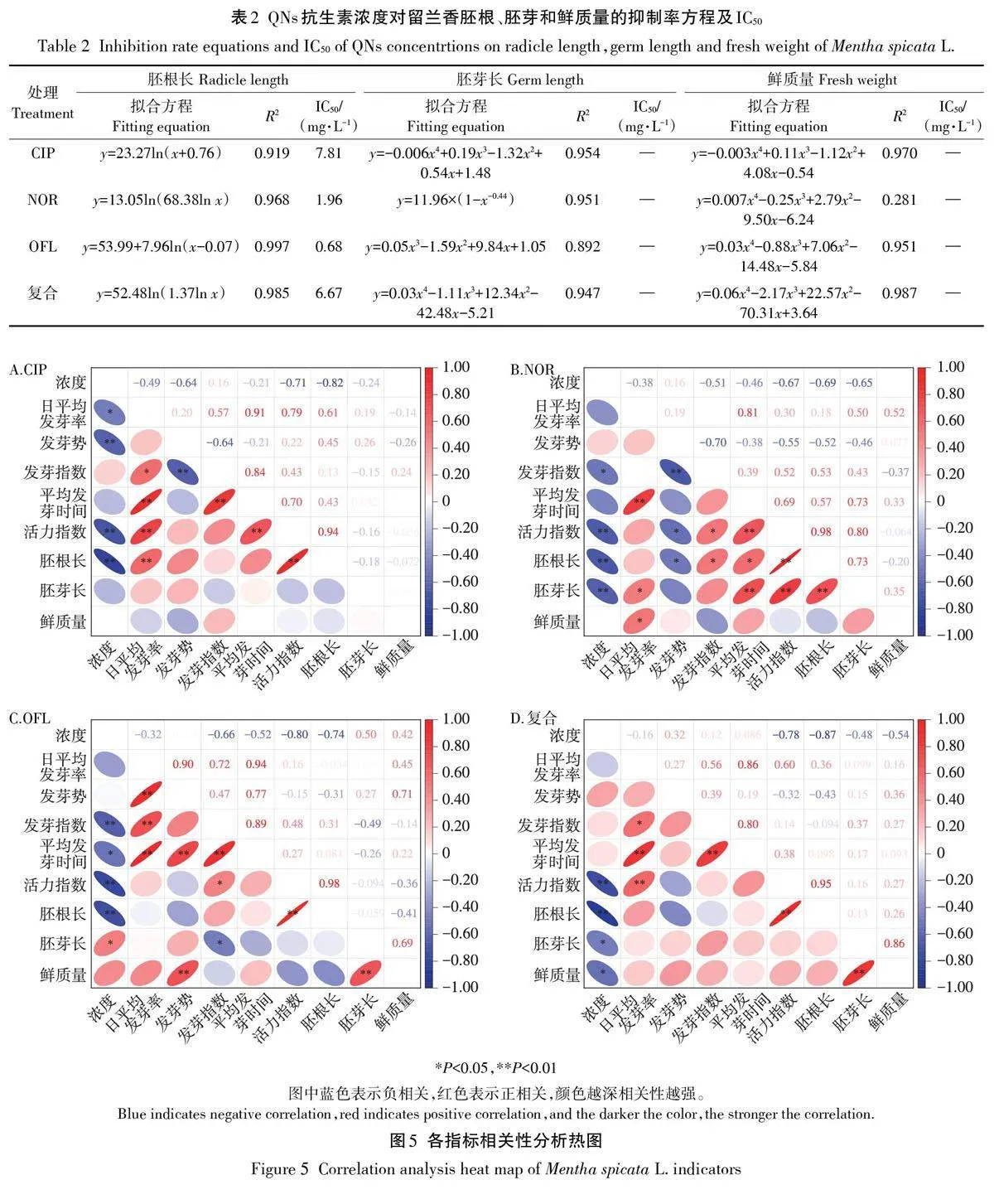
留蘭香各形態指標對QNs抗生素的敏感度不同,通過QNs抗生素脅迫留蘭香種子胚根、胚芽和鮮質量的生長抑制率擬合回歸方程計算IC50(表2)。QNs抗生素濃度對胚根長抑制率回歸方程的相關系數(R2)均大于0.9,相關性好,IC50值大小排序為OFLNORgt;復合gt;CIP。胚芽長和鮮質量的IC50值較大,無法通過回歸方程計算,胚根長可作為QNs抗生素對留蘭香生態毒性的敏感指標。留蘭香萌發期各形態指標的可塑性指數大小為復合(1.449)gt;OFL(1.374)gt;NOR(1.124)gt;CIP(0.849)。萌發期各指標的累計可塑性指數大小為OFL(4.448)gt;復合(3.468)gt;CIP(3.071)gt;NOR(3.038)。
2.4 留蘭香不同指標與濃度間及各指標間的相關性和PLS-SEM分析
如圖5所示,留蘭香種子的活力指數、胚根長與QNs抗生素濃度極顯著負相關(Plt;0.01),線性關系良好(| r |gt;0.6)。除CIP處理外,其他處理的胚芽長與濃度顯著相關(Plt;0.05)。OFL處理下與濃度相關的指標最多,該處理下發芽指數、胚根長與濃度極顯著負相關(Plt;0.01),胚芽長與濃度顯著正相關(Plt;0.05)。各處理組中日平均發芽率與平均發芽時間均呈極顯著正相關(Plt;0.01)。
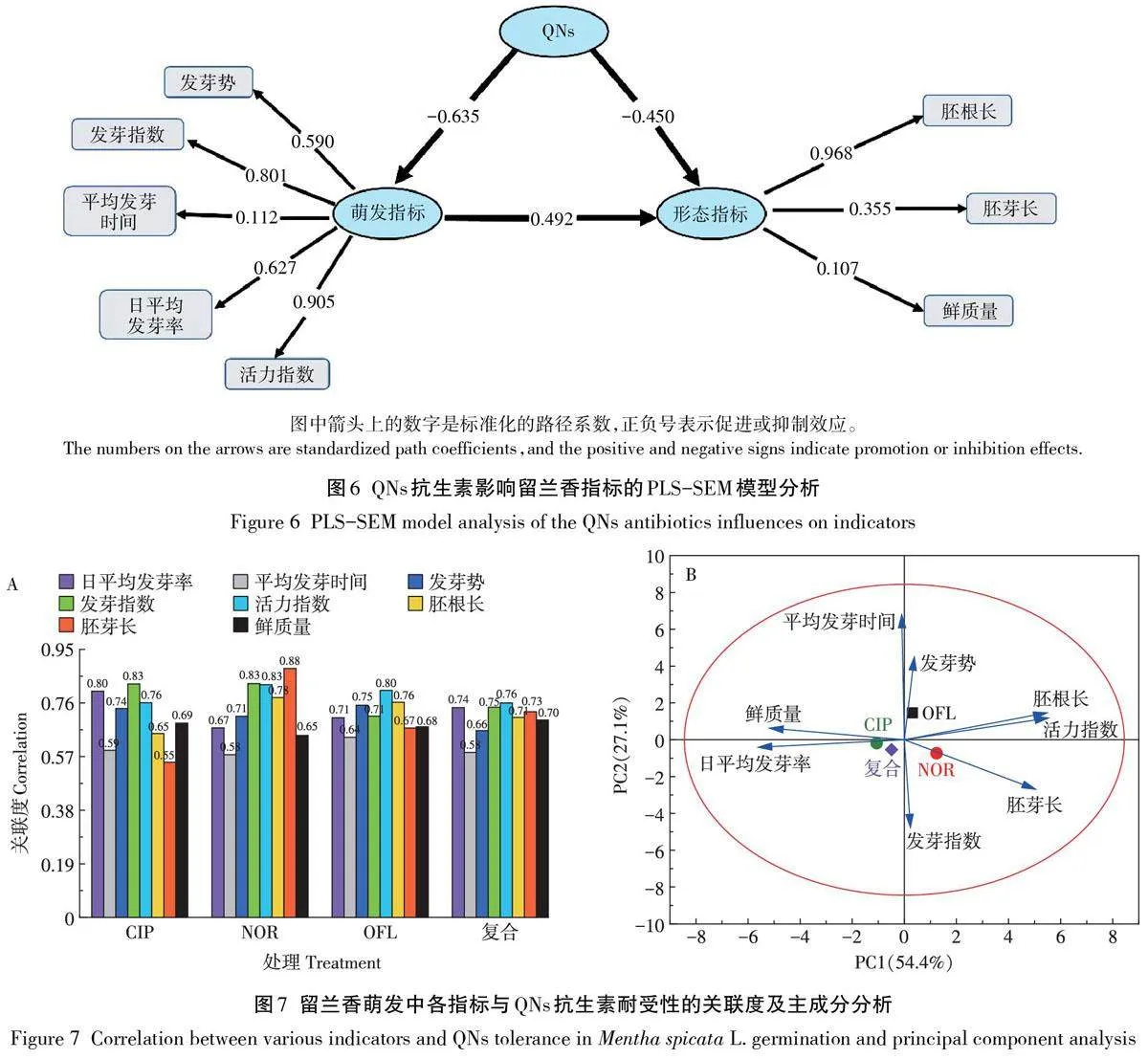
PLS-SEM探究QNs抗生素對留蘭香種子的潛在變量(萌發指標和幼苗形態指標)影響發現,QNs抗生素濃度對留蘭香種子萌發和幼苗生長有顯著負效應,且對萌發的抑制程度大于幼苗生長,萌發指標與形態指標呈極顯著正相關(Plt;0.01)(圖6)。該模型共線性指數(VIF)均小于10,不存在共線性問題,區別效度(Discriminant validity)中HTMT 指數均小于0.90,模型擬合程度較好,結果可信。
2.5 QNs 抗生素脅迫下留蘭香的耐受性及各指標與耐受性的關聯度分析
指標的平均隸屬函數值可綜合評價留蘭香萌發期對QNs 抗生素耐受性大小(表3),0.1 mg·L-1 CIP、0.1 mg·L-1 NOR、CK組OFL和5 mg·L-1復合處理時指標的平均隸屬函數值最大,耐受性最高;綜合各平均隸屬函數值,比較不同QNs抗生素的綜合處理耐受性大小為OFLgt;復合gt;CIPgt;NOR。隸屬函數方法評價結果與可塑性指數法評價結果一致,進一步解釋了圖1各處理組生長情況的差異,QNs抗生素對留蘭香的毒性為OFLlt;復合
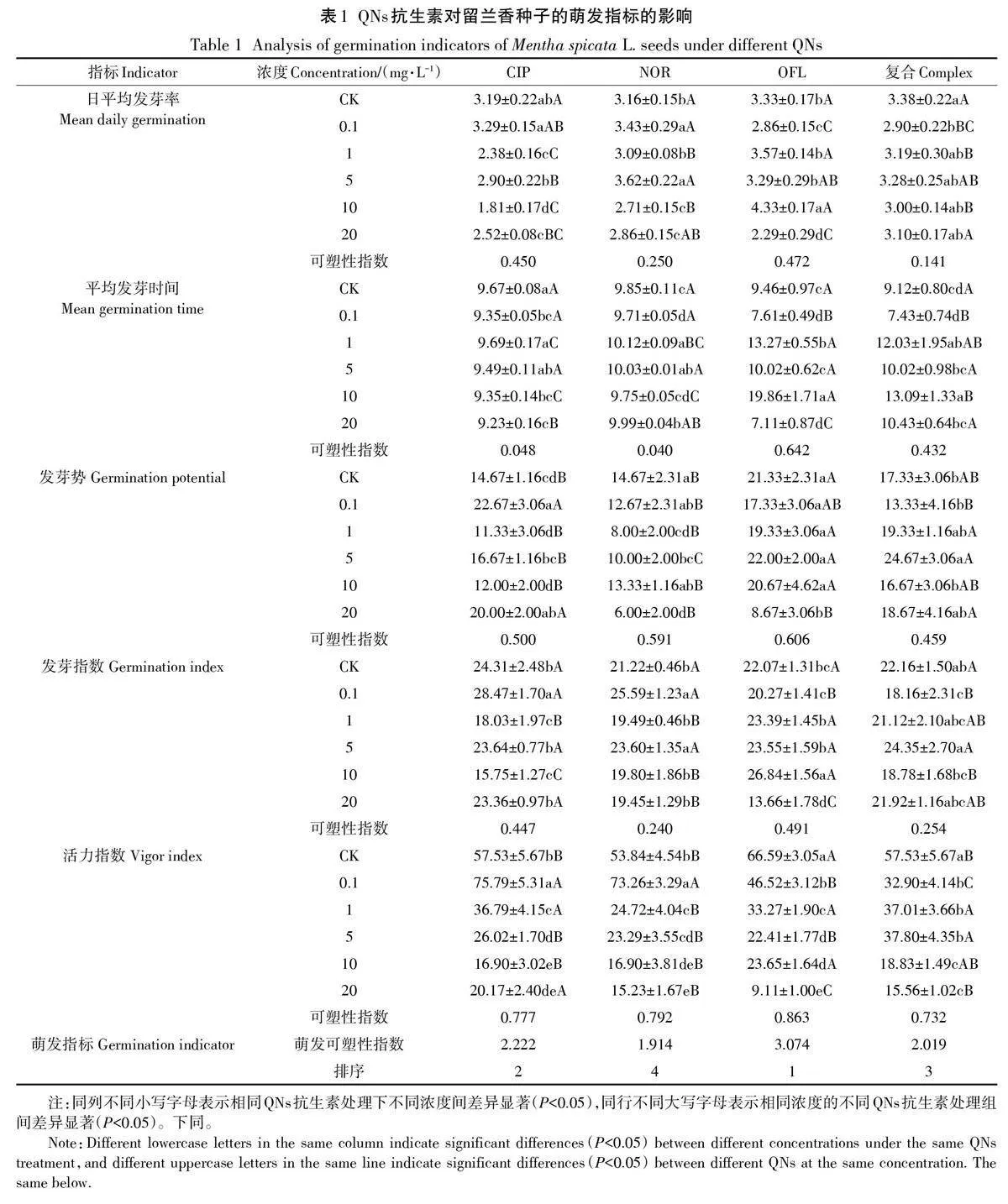
利用灰色關聯度分析法研究各指標與對QNs抗生素綜合耐受度的關聯程度,各QNs抗生素處理組的指標與綜合耐受關聯度大小順序不同,關聯度最高的5個指標的關聯度值均大于0.68(圖7A)。綜合各指標QNs抗生素耐受性關聯度的主成分分析可知,留蘭香對QNs抗生素綜合耐受性的影響主要與日平均發芽率、活力指數、胚根長、鮮質量和胚芽長密切相關(圖7B)。鑒于前人對種子萌發過程中耐受性的評價方法,可將發芽率、胚根長、鮮質量和胚芽長作為抗生素耐受性評價的關鍵指標。
3 討論
3.1 QNs 抗生素濃度對留蘭香種子萌發和幼苗生長的影響
抗生素對種子萌發的影響存在“低濃度下促進和高濃度下抑制”的雙重效應[36-37]。植物種子在萌發過程中受到污染物的影響時,其可通過自身代謝,防止、降低或修復由逆境造成的傷害[38]。本研究發現,低濃度CIP和NOR處理促進發芽,此時留蘭香種子的萌發及形態指標的響應變化,有助于其對QNs抗生素耐受性的提高,這與其他類抗生素脅迫植物種子萌發的結果一致[39-40]。造成低濃度促進現象的原因可能是低濃度的污染脅迫可提高胚的生理活性,促進萌發[41];也可能是因為植物此時產生的γ-氨基丁酸(GABA)可增強α-淀粉酶活性,加速可溶性糖和其他營養物質的產生,為種子萌發提供能量,增強種子活力,為后續幼苗的生長奠定基礎[42]。
當抗生素濃度在植物種子體內的積累超出其生理耐受性時,高濃度的抗生素會抑制淀粉酶、蛋白酶的活性[43],同時抑制種子內儲藏的淀粉和蛋白質的分解,使活性氧清除系統被破壞,保護酶系統失衡,抑制胚芽和胚根的生長,使幼苗生長受到抑制[33]。
3.2 QNs 抗生素種類對留蘭香種子萌發及幼苗生長的影響
不同種類QNs抗生素對留蘭香種子萌發和幼苗生長的影響也不同。種皮在種子萌發過程中對QNs抗生素的阻礙程度的差異,與QNs抗生素理化性質有關。有研究表明,脂水分配系數(lg Kow)可能是影響抗生素植物毒性的最重要因素,其次是相對分子質量(Molecular mass,MW)[44-45]。本研究表明,3種QNs抗生素對留蘭香的毒性大小為NORgt;CIPgt;OFL,這與閆嘉寧等[46]關于NOR、CIP和OFL對蛋白核小球藻的毒性研究結果一致。QNs抗生素的毒性與各自的lg Kow正相關,而與MW 負相關。NOR 結構中含有脂肪鏈烴(疏水性官能團乙基),比CIP和OFL具有更高的疏水性和吸附系數[2],從而增加了與植物根細胞的接觸機會,因此在培養過程中可能更易吸附在根上,抑制根的生長。研究表明,親水性有機外源物(lg Kow 1~4)很容易被根部吸收并轉運[47],而QNs 抗生素的lg Kow通常小于1,很難在植物組織中轉運[48]。種皮在種子的萌發過程中發揮著重要的保護作用,可避免抗生素脅迫對種子發芽產生的不利影響,留蘭香種子的種皮對QNs抗生素阻礙作用的差異導致QNs抗生素毒性的不同。QNs抗生素脅迫下萌發及形態指標的可塑性指數增加,可能是植物通過各指標水平提高達到補償的作用,以維持自身正常生長,提高對QNs抗生素的耐受程度。本研究初步探索了QNs抗生素單一及復合形式對留蘭香種子萌發的毒性效應,結果顯示復合處理的生態毒性介于單一脅迫之間。3種抗生素的毒性相互作用類型與變化趨勢和抗生素混合體系組分濃度比、濃度范圍和種類有關[46]。
4 結論
(1)環丙沙星(CIP)、諾氟沙星(NOR)脅迫對留蘭香的日平均發芽率、發芽指數、活力指數、胚根長等指標存在低促高抑的雙重效應,喹諾酮類抗生素對萌發指標影響程度大于幼苗形態指標;胚根長的半數抑制濃度小于胚芽長和鮮質量,可作為對留蘭香生態毒性的敏感指標。
(2)各指標與喹諾酮類抗生素濃度的相關程度不同,活力指數和胚根長與濃度極顯著負相關(Plt;0.01),氧氟沙星(OFL)脅迫下與濃度相關的指標最多。
(3)留蘭香對喹諾酮類抗生素的耐受性大小為OFLgt;復合gt;CIPgt;NOR。留蘭香5 項指標與耐受性密切相關,關聯度大小為日平均發芽率gt;活力指數gt;胚根長gt;鮮質量gt;胚芽長,發芽率、胚根長、鮮質量和胚芽長可作為留蘭香對抗生素耐受性評價的關鍵指標。
