兩階段pH控制和碳氮源協(xié)同補(bǔ)加促進(jìn)谷氨酸棒桿菌高產(chǎn)L-谷氨酰胺
2024-01-18 09:35:50劉暢陸丹丹浦軍平張春枝陳明
食品與發(fā)酵工業(yè) 2024年1期
劉暢,陸丹丹,浦軍平,張春枝,陳明*
1(大連工業(yè)大學(xué) 生物工程學(xué)院,遼寧 大連,116034)2(張家港市華昌藥業(yè)有限公司,江蘇 蘇州,215635)
L-谷氨酰胺是L-谷氨酸的γ-羥基酰胺化產(chǎn)物,是人類及其他哺乳動(dòng)物體液中含量最豐富的一種游離氨基酸[1],對(duì)細(xì)胞增殖、保護(hù)腸道、減輕炎癥、增強(qiáng)免疫、調(diào)節(jié)相關(guān)疾病通路等具有重要生理功能[2-4]。L-谷氨酰胺廣泛應(yīng)用于醫(yī)藥、保健食品、飼料等領(lǐng)域[5-6],其市場(chǎng)需求量日益增加。因此,增加我國(guó)L-谷氨酰胺產(chǎn)量、提高其工業(yè)化生產(chǎn)水平具有重要意義。
微生物發(fā)酵法是L-谷氨酰胺工業(yè)化生產(chǎn)的主要方法,具有成本低廉、質(zhì)量可控、適宜大規(guī)模生產(chǎn)等優(yōu)勢(shì)。谷氨酸棒桿菌(Corynebacteriumglutamicum)是其主要生產(chǎn)菌種[7],但目前仍存在發(fā)酵水平不夠高、糖酸轉(zhuǎn)化率低、發(fā)酵周期較長(zhǎng)等制約工業(yè)化發(fā)酵生產(chǎn)的關(guān)鍵技術(shù)問(wèn)題。白婧[8]利用枯草芽孢桿菌谷氨酰胺合成酶基因構(gòu)建重組谷氨酸棒桿菌,搖瓶發(fā)酵L-谷氨酰胺產(chǎn)量達(dá)32.5 g/L;王書平等[9]通過(guò)谷氨酸棒桿菌誘變,篩選L-谷氨酰胺高產(chǎn)菌株,搖瓶產(chǎn)量達(dá)33.54 g/L;郭春前等[10]通過(guò)優(yōu)化培養(yǎng)基組分,將L-谷氨酰胺搖瓶產(chǎn)量提高到41.0 g/L;江衍哲[11]對(duì)菌種進(jìn)行基因改造,5 L發(fā)酵罐中L-谷氨酰胺產(chǎn)量達(dá)57.5 g/L。
谷氨酸棒桿菌生物合成L-谷氨酰胺代謝途徑見(jiàn)圖1:葡萄糖經(jīng)過(guò)糖酵解途徑(embden meyerhof pathway,EMP)生成丙酮酸,進(jìn)入三羧酸(tricarboxylic acid,TCA)循環(huán),中間產(chǎn)物α-酮戊二酸經(jīng)谷氨酸脫氫酶(glutamate dehydrogenase,GDH)催化與NH4+反應(yīng)生成L-谷氨酸,進(jìn)一步經(jīng)谷氨酰胺合成酶(glutamine synthetase, GS)催化與NH4+反應(yīng)生成L-谷氨酰胺,生成的L-谷氨酰胺可在谷氨酸合成酶(glutamate synthase, GOGAT)催化下與α-酮戊二酸反應(yīng)生成2分子L-谷氨酸。可見(jiàn),較高的GS活性、較低的GOGAT活性、充足的碳源(葡萄糖)、氮源(NH4+)和ATP供應(yīng),是谷氨酸棒桿菌大量合成并積累L-谷氨酰胺的必要條件[12],研究表明,谷氨酸棒桿菌生長(zhǎng)最適pH值為7.0左右,GS最適pH為5.6左右,而GOGAT在pH值為7.0~8.0活性較強(qiáng)[8,13]。因此,發(fā)酵過(guò)程中分階段控制pH策略有利于前期菌體生長(zhǎng)和后期L-谷氨酰胺積累。L-谷氨酰胺的碳骨架來(lái)源于葡萄糖,其胺基來(lái)源于NH4+,培養(yǎng)基中適宜的碳氮源比例有利于L-谷氨酰胺產(chǎn)量的提高[14];同時(shí),在碳氮源充足的情況下,GS活性相較于氮源不足時(shí)更高,而GOGAT的活性則不受影響[15]。因此,在發(fā)酵過(guò)程中協(xié)同補(bǔ)加葡萄糖和(NH4)2SO4,不僅能保證發(fā)酵過(guò)程中碳氮源充足供應(yīng),而且還能提高GS活性。此外,充足的ATP則可通過(guò)改善O2供應(yīng)來(lái)實(shí)現(xiàn)。基于上述分析,本文在響應(yīng)面法優(yōu)化發(fā)酵培養(yǎng)基組成的基礎(chǔ)上,在50 L發(fā)酵罐中采用兩階段控制pH的碳氮源協(xié)同補(bǔ)加策略,促進(jìn)菌體高效合成L-谷氨酰胺,以期為L(zhǎng)-谷氨酰胺的工業(yè)化發(fā)酵生產(chǎn)提供參考。
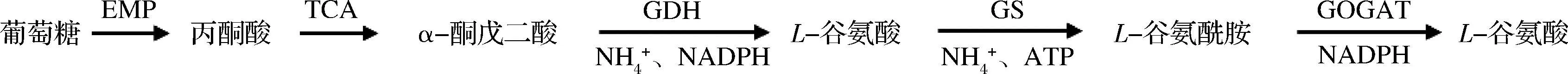
圖1 谷氨酸棒桿菌中L-谷氨酰胺生物合成途徑Fig.1 Schematic of L-glutamine biosynthetic pathway in C.glutamicum
1 材料與方法
1.1 材料
1.1.1 菌種
谷氨酸棒桿菌(Corynebacteriumglutamicum),大連工業(yè)大學(xué)生物工程學(xué)院微生物菌種室保藏。
1.1.2 儀器與設(shè)備
UV-2450紫外可見(jiàn)分光光度計(jì),島津企業(yè)管理(中國(guó))有限公司;SPX-250C恒溫恒濕箱,上海博訊實(shí)業(yè)有限公司醫(yī)療設(shè)備廠;HYL-B往復(fù)式全溫?fù)u瓶柜,太倉(cāng)市強(qiáng)樂(lè)實(shí)驗(yàn)設(shè)備有限公司;TG16-WS臺(tái)式高速離心機(jī),湘儀集團(tuán);50 L自動(dòng)控制發(fā)酵罐,上海百侖生物科技有限公司。
1.1.3 培養(yǎng)基
斜面完全培養(yǎng)基(g/L):牛肉膏15、蛋白胨10、酵母浸粉5、NaCl 5、瓊脂粉25,pH 7.0。
種子培養(yǎng)基(g/L):葡萄糖50、玉米漿55、尿素6、KH2PO41、K2HPO41、MgSO4·7H2O 0.5,pH 7.0。
初始發(fā)酵培養(yǎng)基(g/L):葡萄糖140、(NH4)2SO450、玉米漿20、KH2PO43.5、MgSO4·7H2O 0.6、MnSO4·H2O 0.01、ZnSO4·7H2O 0.01、CuSO4·5H2O 0.01、CaCO330,pH 7.0。
1.2 實(shí)驗(yàn)方法
1.2.1 搖瓶發(fā)酵
取生長(zhǎng)良好的谷氨酸棒狀桿菌斜面菌種,挑取一環(huán),接種到裝有50 mL種子培養(yǎng)基的500 mL三角瓶中,于32 ℃、110 r/min振蕩培養(yǎng)12 h。按8%接種量將種子液接入裝有20 mL發(fā)酵培養(yǎng)基的500 mL三角瓶中,32 ℃、110 r/min振蕩培養(yǎng)。
1.2.2 發(fā)酵培養(yǎng)配方優(yōu)化
選擇發(fā)酵培養(yǎng)基中的葡萄糖、(NH4)2SO4和玉米漿作為3個(gè)因素,以發(fā)酵72 h的L-谷氨酰胺產(chǎn)量為響應(yīng)值,采用Box-Behnken設(shè)計(jì)3因素3水平的響應(yīng)面試驗(yàn),因素及水平如表1所示。
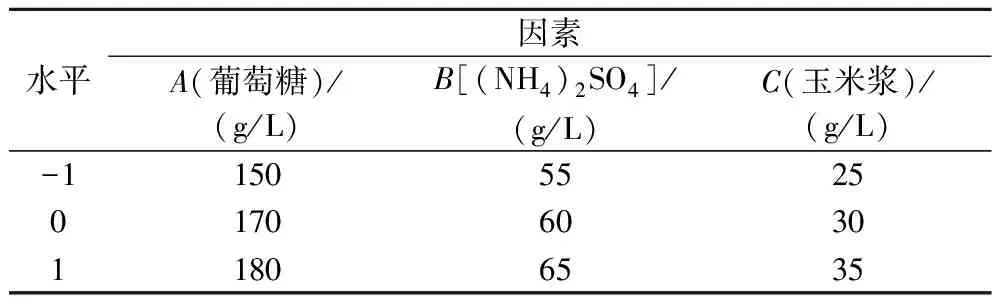
表1 Box-Behnken試驗(yàn)因素及水平Table 1 Factors and levels in Box-Behnken experiment
1.2.3 50 L發(fā)酵罐中L-谷氨酰胺發(fā)酵
1.2.3.1 控制pH 7.0的分批發(fā)酵
按總裝液量35 L配制發(fā)酵培養(yǎng)基,滅菌冷卻后,調(diào)pH 7.0,按照10%接種量接入種子液,100 r/min、32 ℃培養(yǎng),流加氨水控制發(fā)酵液pH 7.0,培養(yǎng)80 h,通氣量分階段控制(0~10 h,0.5 vvm;10~65 h,0.7 vvm;65~80 h,0.5 vvm),當(dāng)接種后溶氧從100%降至10%時(shí),設(shè)置溶氧關(guān)聯(lián)攪拌轉(zhuǎn)速,控制發(fā)酵液的溶氧10%~15%范圍。
1.2.3.2 兩階段控制pH的碳氮源協(xié)同補(bǔ)加發(fā)酵
按總裝液量35 L配制發(fā)酵培養(yǎng)基,滅菌冷卻后,調(diào)pH 7.0,按照10%接種量接入種子液,100 r/min、32 ℃培養(yǎng),采用兩階段控制pH:0~12 h流加氨水控制pH 7.0,12 h后控制pH≥5.6,培養(yǎng)52 h,通氣量分階段控制(0~4 h,0.5 vvm;4~32 h,1.1 vvm;32~52 h,0.5 vvm),當(dāng)接種后溶氧從100%降至10%時(shí),設(shè)置溶氧關(guān)聯(lián)攪拌轉(zhuǎn)速,控制發(fā)酵液的溶氧10%~15%范圍;在發(fā)酵過(guò)程的第24、36 h時(shí)分2次且每次補(bǔ)加2.5 L質(zhì)量濃度為500 g/L的葡萄糖溶液和1 L質(zhì)量濃度為300 g/L的(NH4)2SO4溶液。
1.2.4 菌體濃度測(cè)定
用0.25 mol/L的鹽酸將發(fā)酵液稀釋到適當(dāng)倍數(shù)后于600 nm波長(zhǎng)下測(cè)定吸光度。
1.2.5 殘?zhí)呛繙y(cè)定
采用斐林試劑滴定法。
1.2.6 (NH4)2SO4含量測(cè)定
采用靛酚藍(lán)比色法[16]。
1.2.7L-谷氨酰胺含量的測(cè)定
利用紙層析-色斑洗脫比色法測(cè)定L-谷氨酰胺含量[17],展開(kāi)劑為苯酚-水(4∶1,體積比)。配制質(zhì)量濃度為5、10、15、20、25、30、35、40 g/L的L-谷氨酰胺標(biāo)準(zhǔn)溶液,分別取1 μL點(diǎn)樣于濾紙進(jìn)行層析展開(kāi)。展開(kāi)完成后烘干,噴上5 g/L茚三酮-丙酮溶液進(jìn)行顯色。顯色后將L-谷氨酰胺顯色斑點(diǎn)剪下,裝入具塞試管,利用體積比V(0.1% CuSO4)∶V(75%乙醇)=2∶38的洗脫液進(jìn)行洗脫,洗脫液體積為10 mL,洗脫液置于37 ℃水浴40 min,在波長(zhǎng)520 nm的條件下測(cè)量吸光度,制作L-谷氨酰胺濃度標(biāo)準(zhǔn)曲線。
取1.5 mL發(fā)酵液于離心管中,4 000 r/min離心5 min,取1 μL上清液點(diǎn)樣于濾紙進(jìn)行展開(kāi),進(jìn)行上述相同顯色及洗脫操作,在波長(zhǎng)520 nm的條件下測(cè)量吸光度,對(duì)照標(biāo)準(zhǔn)曲線計(jì)算相應(yīng)L-谷氨酰胺濃度。
1.2.8 發(fā)酵液pH值的測(cè)定
采用pH計(jì)測(cè)定。
1.3 數(shù)據(jù)處理與分析
利用Design Expert V 8.0.6軟件進(jìn)行數(shù)據(jù)處理和分析,使用Origin 2018軟件進(jìn)行方程擬合及繪圖。
2 結(jié)果與分析
2.1 搖瓶條件下發(fā)酵培養(yǎng)基的響應(yīng)面優(yōu)化
2.1.1 Box-Behnken試驗(yàn)結(jié)果
利用Box-Behnken設(shè)計(jì)響應(yīng)面試驗(yàn),L-谷氨酰胺發(fā)酵結(jié)果如表2所示。
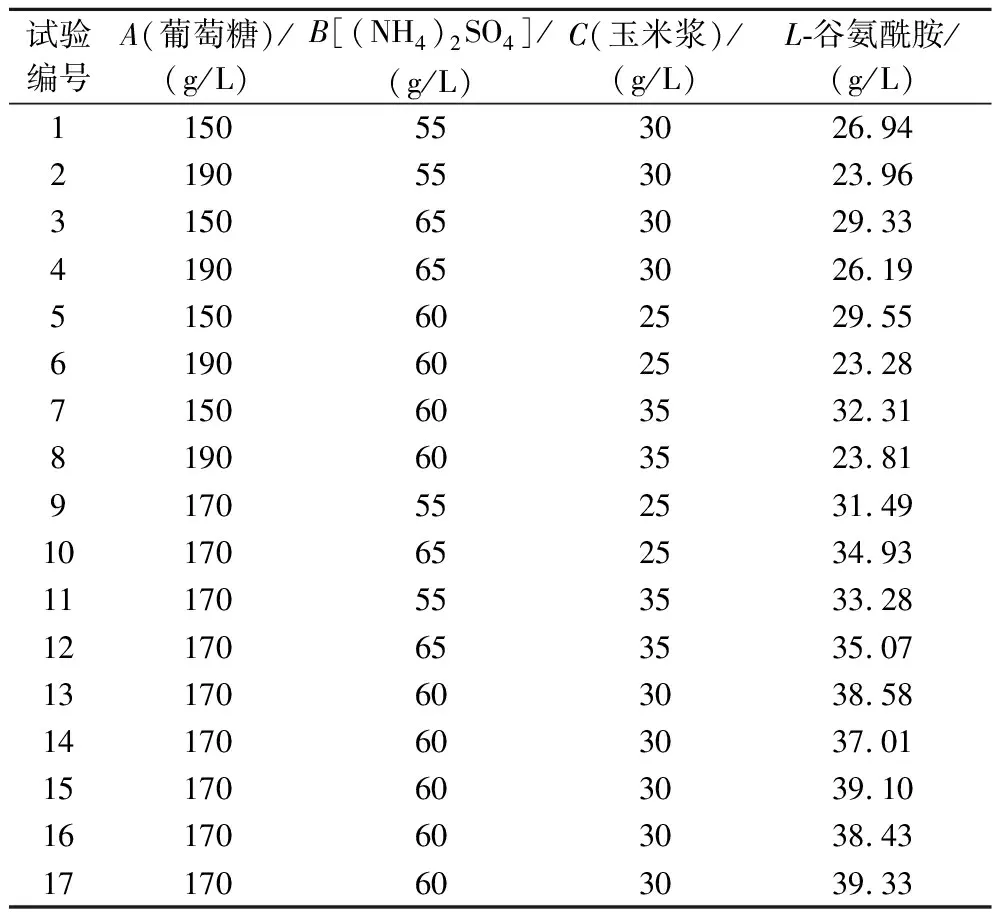
表2 Box-Behnken試驗(yàn)設(shè)計(jì)及結(jié)果Table 2 Design and results of Box-Behnken experiment
2.1.2 方差分析
通過(guò)Design Expert V 8.0.6軟件分析得到L-谷氨酰胺產(chǎn)量的回歸方程如公式(1)所示:
Y=38.49-2.61A+1.23B+0.65C-0.037AB-0.56AC-0.41BC-9.17A2-2.72B2-2.08C2
(1)
用Box-Behnken設(shè)計(jì)構(gòu)建數(shù)學(xué)模型進(jìn)行回歸方程的方差分析,所得結(jié)果如表3所示;3種因素交互對(duì)響應(yīng)值的影響所對(duì)應(yīng)的響應(yīng)面圖如圖2所示。由表3可知,本次設(shè)計(jì)所得模型F值為30.21,有較好的顯著性(P<0.000 1),失擬項(xiàng)不顯著(P=0.109 5>0.05),說(shuō)明此方程回歸顯著。在該模型可信度分析中,方程的決定系數(shù)R2=0.974 9,說(shuō)明所得到的L-谷氨酰胺含量的真實(shí)值與預(yù)測(cè)值之間有良好的擬合度,變異系數(shù)C·V%=4.25%,小于10%,表明該試驗(yàn)穩(wěn)定性強(qiáng),可用此模型來(lái)對(duì)3種物質(zhì)對(duì)L-谷氨酰胺產(chǎn)量的影響進(jìn)行分析和預(yù)測(cè);調(diào)整后的決定系數(shù)AdjR2=0.942 6,該模型具有較高的可信度。響應(yīng)值Y與A、A2、B2顯著性差異極顯著(P<0.01),與B、C2差異顯著(P<0.05),提示各試驗(yàn)因素對(duì)L-谷氨酰胺產(chǎn)量的影響并非簡(jiǎn)單的線性關(guān)系,根據(jù)回歸方程一次項(xiàng)系數(shù)的絕對(duì)值,可得出影響L-谷氨酰胺產(chǎn)量的主次因素為葡萄糖(A)>(NH4)2SO4(B)>玉米漿(C)。
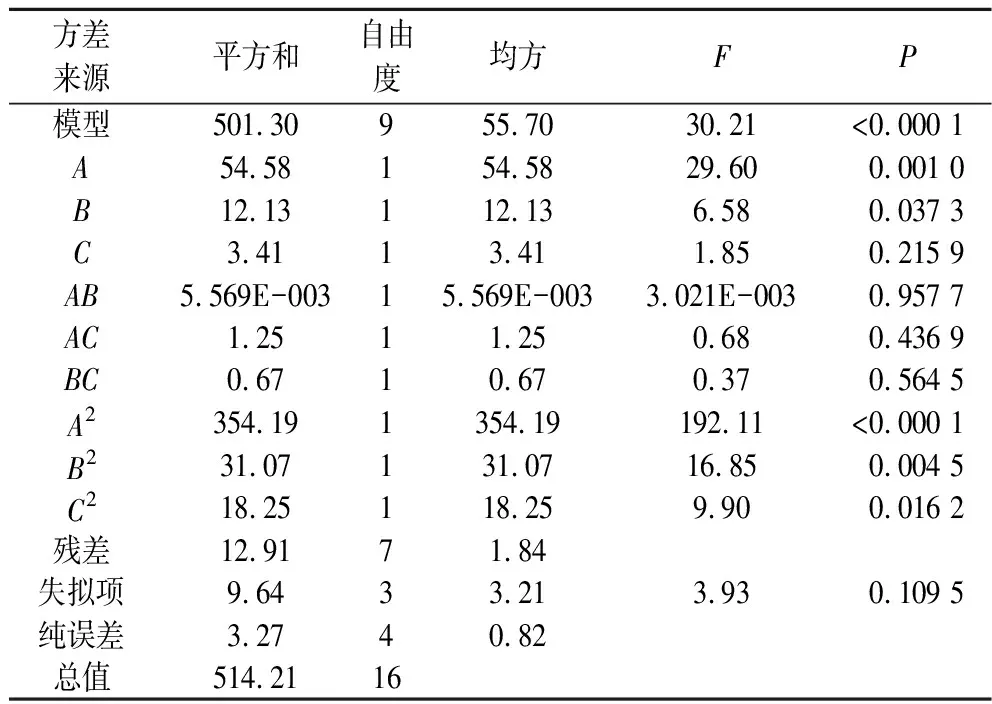
表3 回歸方程的方差分析Table 3 Analysis of variance for regression equation
2.1.3 最佳配方確定與檢驗(yàn)
通過(guò)Design Expert V 8.0.6軟件分析,最適葡萄糖、(NH4)2SO4和玉米漿的添加量分別為167.05、61.08、30.78 g/L,此條件下L-谷氨酰胺的產(chǎn)量達(dá)最高值38.89 g/L。數(shù)值取整后各因素最適添加量為:葡萄糖167 g/L,(NH4)2SO461 g/L,玉米漿31 g/L。在此條件下重復(fù)3次搖瓶發(fā)酵試驗(yàn),L-谷氨酰胺平均產(chǎn)量為38.13 g/L,與理論預(yù)測(cè)值較為接近,模型模擬結(jié)果可信,L-谷氨酰胺搖瓶發(fā)酵水平較優(yōu)化前提高了34.12%。王霞[18]通過(guò)響應(yīng)面法將L-谷氨酰胺搖瓶發(fā)酵水平提高了18.0%,達(dá)到40.6 g/L;郭春前等[10]通過(guò)響應(yīng)面法優(yōu)化L-谷氨酰胺發(fā)酵培養(yǎng)基,產(chǎn)量提高了37.6%;結(jié)果表明響應(yīng)面法優(yōu)化培養(yǎng)基組成對(duì)于L-谷氨酰胺發(fā)酵水平的提高具有顯著效果。
2.2 利用50 L發(fā)酵罐進(jìn)行L-谷氨酰胺發(fā)酵
2.2.1 控制pH 7.0的分批發(fā)酵
在50 L發(fā)酵罐中分批發(fā)酵進(jìn)程如圖3-a所示。在發(fā)酵前期,菌體處于延遲期,葡萄糖消耗慢;從10 h開(kāi)始,菌體進(jìn)入對(duì)數(shù)生長(zhǎng)期,生長(zhǎng)迅速,葡萄糖消耗量增大,發(fā)酵液中的L-谷氨酰胺開(kāi)始積累;到發(fā)酵后期(65 h以后),菌體處于穩(wěn)定期,菌體量逐漸達(dá)到最大值,代謝產(chǎn)物大量積累,碳源被進(jìn)一步被消耗;發(fā)酵至80 h時(shí),L-谷氨酰胺的積累趨近最大值,此時(shí)L-谷氨酰胺產(chǎn)量達(dá)42.06 g/L,較搖瓶發(fā)酵提高了10.31%,按照實(shí)際發(fā)酵液體積37 L計(jì)算,糖酸轉(zhuǎn)化率為25.85%,生產(chǎn)強(qiáng)度為0.53 g/(L·h)。

a-發(fā)酵進(jìn)程;b-菌體生長(zhǎng)擬合曲線;c-底物消耗擬合曲線;d-產(chǎn)物生成擬合曲線圖3 50 L發(fā)酵罐中控制pH 7.0條件下谷氨酸棒桿菌分批發(fā)酵進(jìn)程及動(dòng)力學(xué)模型擬合曲線Fig.3 Batch fermentation process and fitted curve of kinetic models by C.glutamicum in 50 L fermenter with pH 7.0 control
選擇Logistic方程[19]對(duì)菌體生長(zhǎng)進(jìn)行非線性擬合,結(jié)果如圖3-b所示,其中R2為0.993 6,菌體生長(zhǎng)動(dòng)力學(xué)模型如公式(2)所示:
(2)
選擇Luedeking-Piret-Like方程對(duì)發(fā)酵液中葡萄糖消耗進(jìn)行非線性擬合,結(jié)果如圖3-c所示,其中R2為0.994 4,底物消耗動(dòng)力學(xué)模型如公式(3)所示:
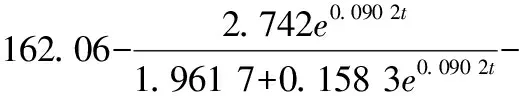
(3)
選用Boltzman模型對(duì)產(chǎn)物L(fēng)-谷氨酰胺生成進(jìn)行非線性擬合,結(jié)果如圖3-d所示,其中R2為0.993 8,產(chǎn)物生成動(dòng)力學(xué)模型如公式(4)所示:
(4)
基于對(duì)50 L發(fā)酵罐中控制pH值為7.0條件下L-谷氨酰胺發(fā)酵進(jìn)程及菌體生長(zhǎng)、底物消耗、產(chǎn)物生成動(dòng)力學(xué)分析,可以看出,整個(gè)發(fā)酵過(guò)程中(0~80 h)谷氨酸棒桿菌處于較好生長(zhǎng)態(tài)勢(shì),說(shuō)明pH值為7.0適合菌體生長(zhǎng),但從葡萄糖消耗和L-谷氨酰胺生成情況來(lái)看,葡萄糖代謝生成L-谷氨酰胺的效率并不高,L-谷氨酰胺產(chǎn)量、生產(chǎn)強(qiáng)度和糖酸轉(zhuǎn)化率較工業(yè)化生產(chǎn)技術(shù)水平還存在較大差距,這可能是由于控制pH值為7.0導(dǎo)致谷氨酸棒桿菌的GS活性較低所致;同時(shí)發(fā)現(xiàn),發(fā)酵前期菌體生長(zhǎng)需要較高的溶氧,所設(shè)定的通氣量可能無(wú)法滿足菌體生長(zhǎng)所需,而通過(guò)溶氧關(guān)聯(lián)攪拌則容易導(dǎo)致攪拌轉(zhuǎn)速過(guò)高,產(chǎn)生較大剪切力,不利于發(fā)酵初期菌體生長(zhǎng),容易導(dǎo)致菌體絮凝[20]。因此,調(diào)整發(fā)酵過(guò)程通氣量并采用分階段控制pH策略,結(jié)合碳氮源協(xié)同補(bǔ)加,可能有利于前期菌體生長(zhǎng)和后期L-谷氨酰胺積累。
2.2.2 兩階段pH控制下的補(bǔ)料分批發(fā)酵結(jié)果
在50 L發(fā)酵罐中采用兩階段控制pH的碳氮源協(xié)同補(bǔ)加策略,發(fā)酵結(jié)果如圖4所示。相較于分批發(fā)酵,補(bǔ)料分批發(fā)酵過(guò)程中菌體生長(zhǎng)的延遲期縮短,提前進(jìn)入對(duì)數(shù)生長(zhǎng)期,說(shuō)明加大通氣量,保持適宜的攪拌轉(zhuǎn)速,有利于促進(jìn)菌體的生長(zhǎng)。培養(yǎng)12 h后控制發(fā)酵液pH≥5.6,此時(shí)菌體處于對(duì)數(shù)生長(zhǎng)期,pH雖有所變化,但菌體仍保持較高生長(zhǎng)速率;同時(shí),葡萄糖和(NH4)2SO4的消耗速率增大,且L-谷氨酰胺產(chǎn)量快速上升,說(shuō)明pH的改變有利于提高GS活性,促進(jìn)了菌體利用碳源和氮源生成L-谷氨酰胺,并為碳氮源的補(bǔ)加創(chuàng)造了條件。在發(fā)酵的第24、36 h,分2次且每次補(bǔ)加2.5 L 500 g/L的葡萄糖溶液和1 L 300 g/L的(NH4)2SO4溶液,補(bǔ)料后發(fā)酵液中的葡萄糖與(NH4)2SO4質(zhì)量濃度比約為3∶1。在第24 h進(jìn)行第一次補(bǔ)料時(shí),菌體處于對(duì)數(shù)生長(zhǎng)中后期,菌體生長(zhǎng)速度減緩,L-谷氨酰胺的積累量在補(bǔ)料后的第8 h(即培養(yǎng)第32 h)即達(dá)到39.30 g/L;在36 h進(jìn)行第2次補(bǔ)料前,L-谷氨酰胺產(chǎn)量已高于50 g/L,第2次補(bǔ)料后,菌體濃度基本保持穩(wěn)定,葡萄糖和(NH4)2SO4持續(xù)被消耗,產(chǎn)物繼續(xù)積累,但由于補(bǔ)料后發(fā)酵液體積增大,L-谷氨酰胺濃度增長(zhǎng)速率降低;至52 h時(shí),發(fā)酵液中葡萄糖和(NH4)2SO4濃度已處于較低水平,分別為8.53和8.91 g/L,結(jié)合50 L發(fā)酵罐中發(fā)酵液體積已接近上限的考慮,不再進(jìn)行補(bǔ)料,于52 h結(jié)束發(fā)酵。
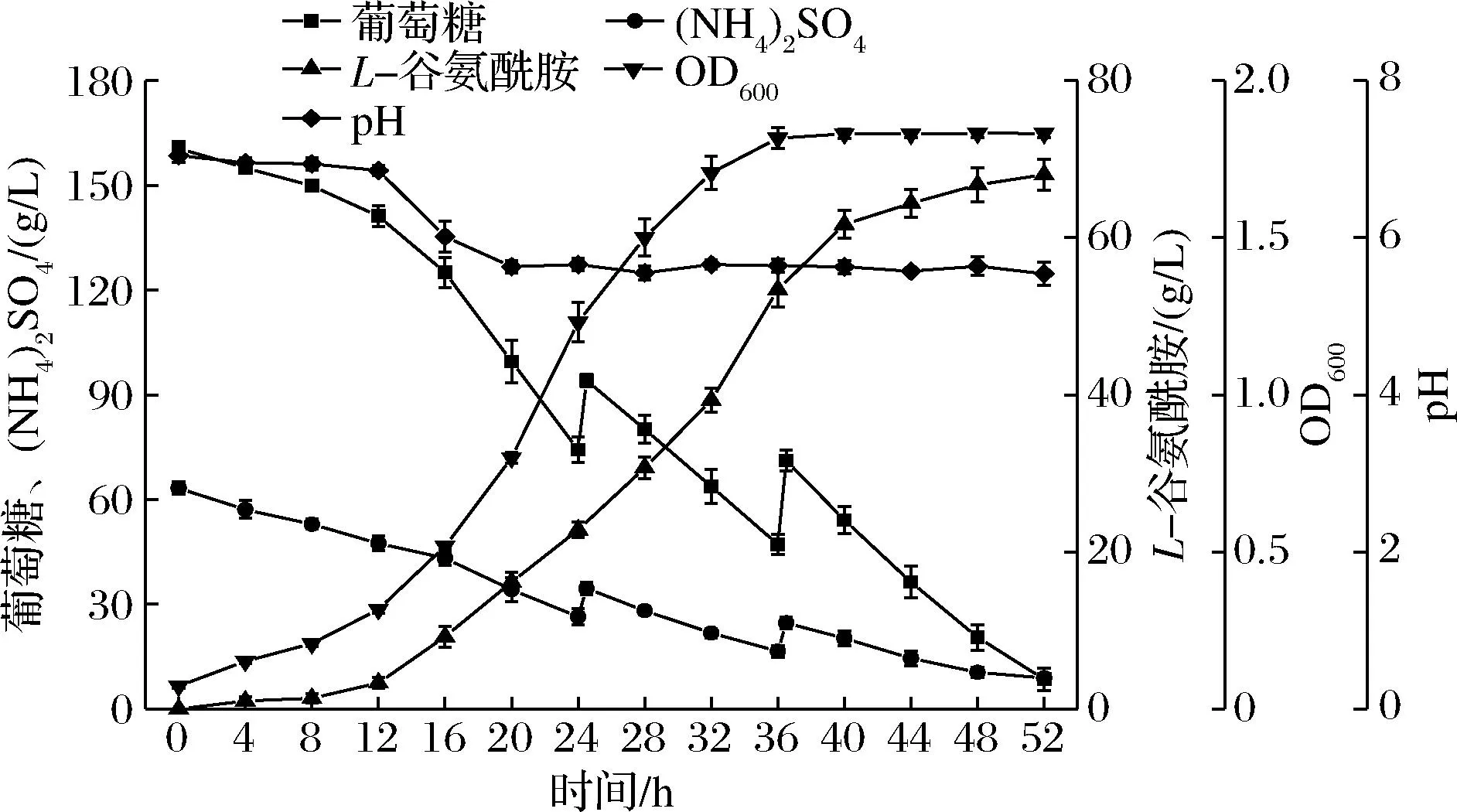
圖4 50 L發(fā)酵罐中兩階段pH控制的碳氮源協(xié)同補(bǔ)加 條件下谷氨酸棒桿菌分批補(bǔ)料發(fā)酵Fig.4 Fed-batch fermentation process of C.glutamicum in 50 L fermenter under two-stage pH control combined with synergistic addition of carbon and nitrogen sources
在50 L發(fā)酵罐中基于兩階段控制pH的碳氮源協(xié)同補(bǔ)加策略,將發(fā)酵周期縮短至52 h,L-谷氨酰胺產(chǎn)量達(dá)68.01 g/L,按照實(shí)際發(fā)酵液體積42.50 L計(jì)算,糖酸轉(zhuǎn)化率達(dá)到33.93%,生產(chǎn)強(qiáng)度為1.31 g/(L·h),顯著高于控制pH 7.0的分批發(fā)酵結(jié)果,這是因?yàn)檫m宜的通氣量及攪拌轉(zhuǎn)速促進(jìn)了菌體生長(zhǎng),同時(shí)兩階段控制pH策略提高了菌體的GS活性以及對(duì)碳氮源的底物利用速率,而且葡萄糖和(NH4)2SO4的協(xié)同補(bǔ)加為L(zhǎng)-谷氨酰胺的生成提供了充足的原料,從而促進(jìn)了谷氨酸棒桿菌高效合成L-谷氨酰胺。李爽等[21]于16 L發(fā)酵罐中進(jìn)行補(bǔ)料分批發(fā)酵44 h后,L-谷氨酰胺的產(chǎn)量為52.2 g/L;呂青蘭[22]在改造谷氨酸棒桿菌并優(yōu)化發(fā)酵策略后,于5 L發(fā)酵罐中補(bǔ)料分批發(fā)酵66 h,達(dá)到33.5%的糖酸轉(zhuǎn)化率和1.35 g/(L·h)的生產(chǎn)強(qiáng)度。
3 結(jié)論
谷氨酸棒桿菌發(fā)酵法是目前工業(yè)化生產(chǎn)L-谷氨酰胺的主要方法,但存在發(fā)酵水平不夠高、糖酸轉(zhuǎn)化率低、發(fā)酵周期長(zhǎng)等制約工業(yè)化生產(chǎn)的關(guān)鍵技術(shù)問(wèn)題。本研究利用一株谷氨酸棒桿菌發(fā)酵產(chǎn)L-谷氨酰胺,采用響應(yīng)面法優(yōu)化了發(fā)酵培養(yǎng)基中葡萄糖、(NH4)2SO4和玉米漿含量,并利用50 L發(fā)酵罐探究了兩階段控制pH的碳氮源協(xié)同補(bǔ)加策略對(duì)發(fā)酵結(jié)果的影響。 結(jié)果表明,發(fā)酵前期(0~12 h)控制pH 7.0和增大通氣量有利于谷氨酸棒桿菌的菌體生長(zhǎng),而發(fā)酵中后期(12~52 h)控制pH 5.6有利于提高L-谷氨酰胺合成關(guān)鍵酶GS的活性,從而促進(jìn)了菌體利用碳源和氮源合成并積累L-谷氨酰胺,并為碳氮源的補(bǔ)加創(chuàng)造了條件。發(fā)酵結(jié)果顯示,50 L發(fā)酵罐中采用兩階段pH控制的碳氮源協(xié)同補(bǔ)加策略,發(fā)酵周期縮短至52 h,L-谷氨酰胺產(chǎn)量達(dá)68.01 g/L,糖酸轉(zhuǎn)化率達(dá)33.93%,生產(chǎn)強(qiáng)度為1.31 g/(L·h),主要技術(shù)指標(biāo)達(dá)到較高水平,對(duì)L-谷氨酰胺的工業(yè)化發(fā)酵生產(chǎn)具有重要參考價(jià)值。