南亞熱帶鄉土闊葉林與馬尾松林土壤細菌群落多樣性和結構
2024-01-20 10:00:06滕金倩肖指柔明安剛席守鴻朱茂鋒
林業科學研究 2023年6期
關鍵詞:分類
滕金倩,肖指柔,明安剛,朱 豪,席守鴻,何 江,朱茂鋒,陶 怡,覃 林*
(1.廣西大學林學院,廣西森林生態與與保育重點實驗室,廣西 南寧 530004;2.中國林業科學研究院熱帶林業實驗中心,廣西 憑祥532600;3.廣西友誼關森林生態系統定位觀測研究站,廣西 憑祥 532600)
為了滿足木材、燃料和其它林產品的需求,將天然闊葉林改造成速生豐產的人工針葉林是過去幾十年里中國南方普遍采用的經營策略[1]。然而,大面積多代連栽的針葉林極易造成一系列嚴重的生態環境問題(如土壤侵蝕、肥力衰退和生態系統穩定性降低等),最終導致人工林生產力的不斷下降[2]。如何科學培育人工林,有效提高其生產力并維持生物多樣性,是人工林多目標經營亟需解決的關鍵問題。有研究表明,將現存的大面積低質低效針葉純林轉化為經濟價值、生產力和物種多樣性高的鄉土闊葉樹種人工林,將會改變土壤理化性質和養分循環,有助于提高土壤肥力[3]。將針葉林改造成鄉土闊葉林,是近年來我國最有前景的人工林經營模式之一。
土壤細菌群落是陸地生態系統中最多樣化和最主要的微生物群落,對生態系統營養物質循環、分解有機質及維持土壤肥力等具有關鍵效用[4]。凋落物與根系分泌物能為土壤細菌生命活動提供所需要的碳源,因而不同樹種可直接引起土壤細菌群落組成多樣性的變化[5]。土壤細菌群落多樣性包括分類和系統發育兩方面。傳統的分類多樣性僅僅量化物種名稱和豐度變化,沒有傳達有關物種的生態和系統發育的關鍵信息,無法解釋群落動態,因此系統發育多樣性的變化越來越受到人們的關注[6]。系統發育多樣性反應了群落內物種之間的系統發育距離和作用,有助于估計生物多樣性對生態系統功能的影響[7]。群落結構是物種在不同空間尺度上相互作用的結果,量化物種分布模式的空間差異可以深入了解調控物種多樣性形成的機制以及群落趨同、轉換率等群落狀態[8]。有研究表明,中國南方地區的針葉林和闊葉林的土壤細菌群落分類結構差異顯著[9]。目前,生態位理論和中性理論分別代表了群落構建的確定性過程(環境過濾和競爭排斥)和隨機性過程(不可預測的干擾、擴散、出生和死亡)[10]。有學者使用群落分類結構的Raup-Crick相異指數(βRC)探究群落構建過程,發現環境過濾是中國華北亞高山林線區域土壤細菌群落構建驅動因素[11]。同時,通過系統發育結構也可以反映群落構建過程中物種的進化歷史,若由環境過濾主導,則篩選出適應能力相似、親緣關系較近的物種,導致系統發育聚類;若由競爭排斥主導,則生態位相似的物種將無法共存,導致系統發育過度分散[12]。至今關于森林土壤細菌群落系統發育結構的研究結果不一。有研究認為,森林土壤(0~10 cm)細菌群落系統發育結構是聚集的,群落構建由確定性過程決定,主要受環境過濾影響[13-14];但也有研究指出,確定性過程和隨機性過程共同作用于森林土壤(0~20 cm)細菌群落構建,其中確定性過程占主要作用[15]。另外,土壤細菌對生物地球化學循環過程的影響在整個土壤剖面中一直存在[16],且不同土層深度的土壤理化性質不同[9],或許會導致不同的細菌群落分布模式。然而,現有的研究僅僅考慮了森林表層土壤(0~20 cm)細菌群落構建機制,對于人工林深層土壤(>20 cm)的細菌群落如何構建知之甚少。
我國亞熱帶地區人工林的面積和蓄積均居全國之首[17]。用紅錐(Castanopsis hystrixMiq.)、米老排(Mytilaria laosensisLec.) 和火力楠(Michelia macclureiDandy)等珍貴鄉土闊葉林更替杉木(Cunninghamia lanceolata(Lamb.)Hook.)和馬尾松(Pinus massonianaLamb)等針葉林是目前該地區常見的營林模式。針葉林改建成闊葉林后,其凋落物數量與質量的改變將會對土壤理化性質產生作用,進而會影響土壤細菌群落多樣性和結構。本研究以南亞熱帶地區杉木林采伐跡地種植的馬尾松、紅錐、米老排和火力楠等人工林為對象,基于各林分不同土層(0~20、20~40 和40~60 cm)細菌16S rRNA 高通量測序數據,旨在通過差異性分析推測未來在馬尾松林采伐后種植鄉土闊葉林情況下土壤細菌群落的分類與系統發育多樣性和結構依土層的變化特征,以及導致變化的主要土壤理化因子。研究結果不僅對針葉林改建鄉土闊葉林的樹種選擇具有指導作用,還為闡明亞熱帶人工林土壤剖面細菌群落構建機制提供科學依據。
1 材料與方法
1.1 研究區概況及樣地設置與樣品采集
2017 年2 月在該區域選取具有立地條件(土壤類型、海拔、坡度、坡向)和經營措施相似的馬尾松、紅錐、米老排和火力楠等4 個鄉土樹種人工林為對象。4 個人工林均是在杉木采伐跡地上種植(密度為2 500 株·hm-2);其中,馬尾松林和紅錐林于1983 年營造并經過了3 次間伐,而米老排林和火力楠林于1981 年營建并經歷2 次間伐,間伐強度約30%。在各林分內,隨機設置3 個20 m × 20 m 的樣地(樣地間距20 m 以上),在每個樣地內沿左對角線任意選擇3 個采樣點,用土鉆(內徑5.0 cm)分別采集表層(0~20 cm)、中層(20~40 cm)和底層(40~60 cm)土樣,除去動植物殘體與石塊等雜質后,將同層土樣混合成一個土樣裝入無菌采樣袋,置于內含生物冰袋的泡沫箱中并帶回實驗室。新鮮混合土樣過2 mm 鋼篩后分為3 份:1 份置于-80 ℃的冰箱內冷藏,用于土壤DNA 提取;1 份置于冰箱4 ℃保存,用于測定土壤硝態氮和銨態氮含量;1 份經自然風干后過0.25 mm 篩保存用于測定土壤基本化學性質。各林分的立地條件和基本特征見表1。
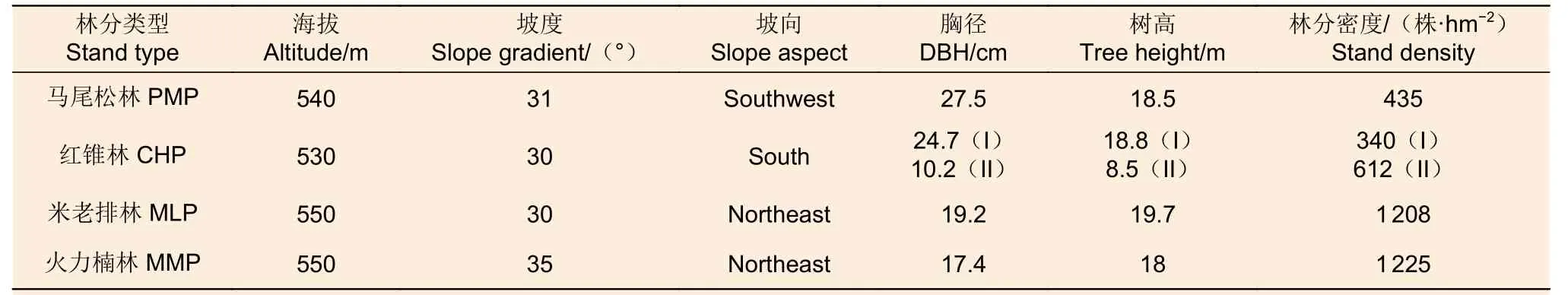
表1 4 個人工林的立地條件和林分特征Table 1 Site conditions and stand characteristics of the four planted forests
1.2 土壤理化性質測定與高通量測序
參照魯如坤[19]測定如下指標:烘干法測定土壤含水量(SWC);pH 計(Prtavo 907 MULTI pH,德國)測定土壤pH 值(土∶水=1∶2.5);重鉻酸鉀法測定土壤有機碳(SOC)含量;酚二磺酸比色法測定硝態氮()含量;擴散法測定銨態氮()含量;雙酸浸提-鉬銻抗比色法測定速效磷(AP)含量。經H2SO4與HClO4消煮后,使用全自動間斷化學元素分析儀SmartChem200(Alliance,法國)測定總氮(TN)和總磷(TP)含量;土壤有機碳與總氮含量的比值即土壤碳氮比(C/N)。
土壤DNA 使用 PowerSoil?DNA Isolation Kit 試劑盒(美國MoBio 公司)提取。提取到的土壤DNA 純度和濃度通過紫外微量分光光度計(Thermo Nano Drop 2000)進行測定。以515F( 5 ′-GTGCCAGCMGCCGCGGTAA-3 ′ )/926R(5′-CCGTCAATTCMTTTGAGTTT-3′)為引物[20],通過PCR 擴增技術進行細菌16S rRNA V4-V5 區基因片段的擴增。兩次PCR 擴增的條件和反應體系見彭雯等[21]。對PCR 產物進行2%瓊脂糖凝膠電泳檢測,使用美國AXYGEN 公司的AxyPrepDNA凝膠回收試劑盒對凝膠進行回收純化,通過FTC-3000TM real-time PCR 儀實現膠回收產物的熒光定量,均一化處理后測序文庫構建結束。由上海微基生物科技有限公司通過Illumina Miseq 2x300 bp完成高通量測序。
根據barcode 信息對高通量測序得到的原始序列進行拼接、質量控制和過濾。有效序列拼接由FLASH 軟件完成,質控和過濾由Trimmomatic軟件實現,優化序列通過Mothur V.1.33.3 軟件獲得。對優化序列基于97% 相似水平的聚類由UPARSE 軟件完成, 生成OTU (operational taxonomic unit)的特征序列,用UCHIME 軟件檢測和去除嵌合體序列,使用Mothur(classify.seqs)軟件比對OTU 特征序列與Silva(Release 123, http://www.arb-silva.de)數據庫后進行物種注釋(置信度閾值設置為0.8)。
以最低樣本測序深度17 291 為基準進行抽平,結果用于后續分析。采用最大似然法構建系統發育樹,在QIIME2 2 021.8 軟件中的FASTTREE程序完成[22]。
1.3 土壤細菌群落分類與系統發育多樣性、結構及轉換率
1.3.1 細菌群落分類與系統發育多樣性 采用Chao1、Shannon 和Simpson 指數反映細菌群落的分類多樣性;PD 指數[23]表征系統發育多樣性,其數值大小取決于物種共同祖先的多寡(即系統發育樹的節點數量)。
1.3.2 細菌群落分類與系統發育結構 利用基于Bray-Cruits 距離的主坐標分析(PCoA)和置換多元方差分析(PERMANOVA)檢測不同林分土壤細菌群落分類結構之間的差異,并用Monte Carlo檢驗理化性質的顯著性。用Raup-Crick 相異指數(βRC)表示兩個群落之間的分類結構相異程度,其取值范圍為[-1,1],可表征群落的構建過程[24]。使用凈親緣關系指數(NRI)和最近親緣關系指數(NTI)量化系統發育結構[25],從而揭示群落構建機制。
1.3.3 細菌群落分類與系統發育轉換率 為了測量不同林分土壤細菌群落分類和系統發育的空間動態變化,使用Bray-Cruits 距離表征群落分類轉換率,使用豐度加權的平均成對系統發育距離(βMPD)和平均最近相鄰系統發育距離(βMNTD)量化群落系統發育轉換率[26]。以上分析均在R 軟件中進行。
總的來說,初中語文的作文教學是提升學生寫作能力的重要關鍵,教師應當重視自身教學中存在的問題,在解決這些問題與困境的基礎上,多讓學生以自己的視角進行人與事的分析,這樣才能夠讓學生的寫作更有內涵,同時也能夠提升他們的實際寫作感悟,進而保障他們寫作能力的提升。
1.4 統計分析
采用單因素方差分析(One-way ANOVA)及多重比較(Tukey)判定相同土層不同林分之間土壤理化性質、細菌群落的分類與系統發育多樣性以及分類與系統發育轉化率的差異。采用Pearson相關方法,探討土壤理化性質與分類和系統發育多樣性之間的相關性。利用SPSS 26.0 軟件(SPSS, Inc, Chicago, IL)完成計算。
2 結果與分析
2.1 馬尾松林與鄉土闊葉林土壤理化性質的比較
方差分析表明(表2),就相同土層不同林分而言,在土壤表層,馬尾松林的SWC 顯著低于米老排林和火力楠林;在中層,馬尾松林的SWC 顯著低于米老排林和火力楠林以及TN 顯著低于火力楠林,但其pH 和均顯著高于紅錐林和米老排林;在底層,馬尾松林的pH 顯著高于火力楠林,而其顯著高于紅錐林和米老排林(P<0.05)。土壤SOC、、TP、AP 及C/N 在相同土層不同林分間均無顯著差異(P>0.05)。

表2 4 個研究林分的土壤理化性質(均值 ± 標準誤,n=3)Table 2 Physical and chemical properties of soil in four plantations (mean ± SE, n=3)
2.2 馬尾松林與鄉土闊葉林土壤細菌群落多樣性的差異
方差分析表明,在土壤表層,馬尾松林的Chao1、PD 指數顯著低于米老排林和火力楠林(P<0.05),而與紅錐林差異不顯著(P>0.05)(圖1A、D),其中,相較于馬尾松林,米老排林的Chao1、PD 指數分別提高了20.5%和20.7%,火力楠林則提高了23.8%和20.6%;馬尾松林與3 鄉土闊葉林的Shannon 和Simpson 指數均無顯著差異(圖1B、C)。在中層和底層,馬尾松林與3 個鄉土闊葉林的分類和系統發育多樣性指數均無顯著差異(P>0.05)(圖1)。
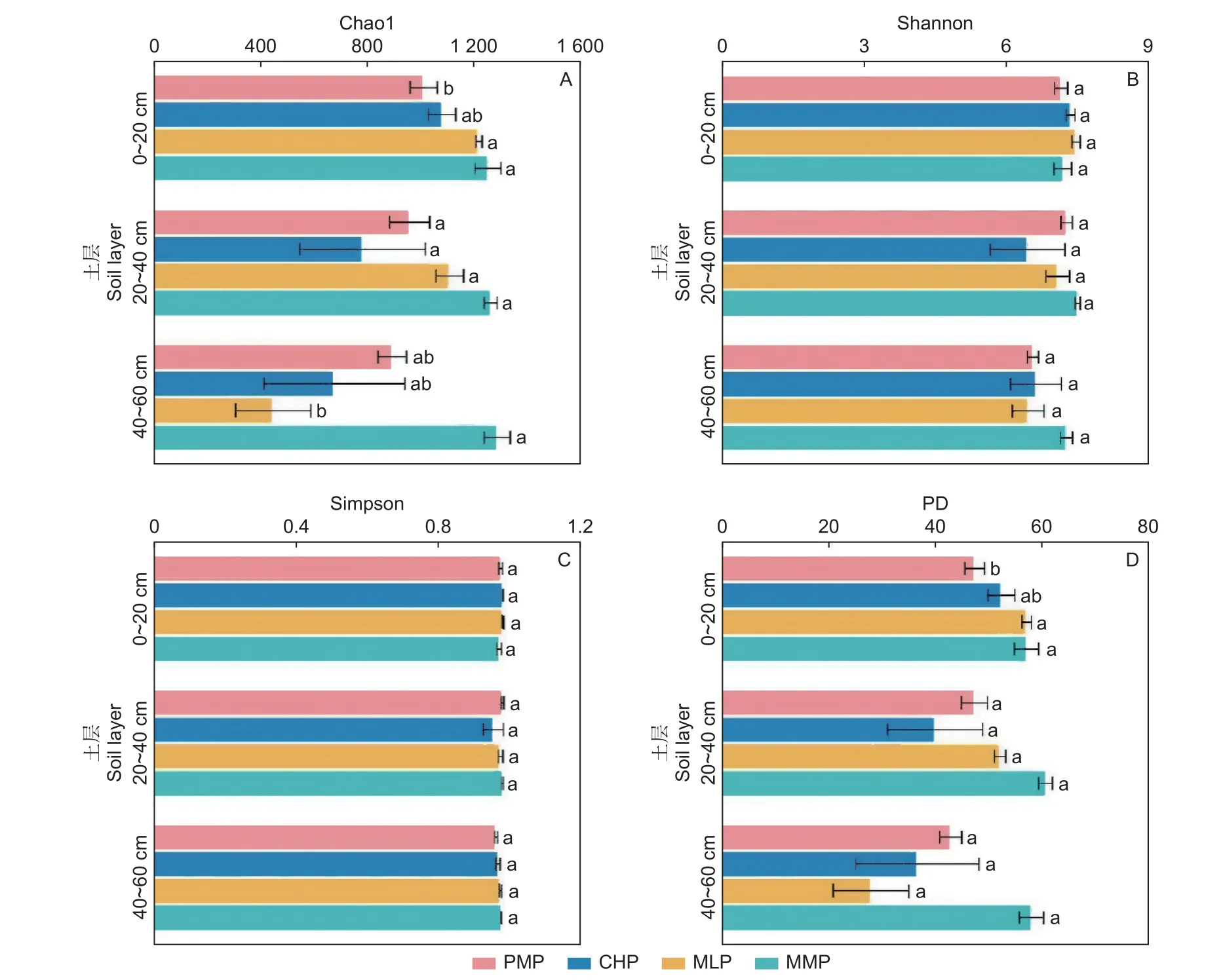
圖1 4 個人工林土壤細菌群落的分類與系統發育多樣性(均值 ± 標準誤,n=3)Fig.1 Taxonomic and phylogenetic diversity of soil bacterial communities in four plantations (mean ± SE, n=3)
土壤理化性質與土壤細菌群落多樣性指數的Pearson 相關性分析表明(圖2),Chao1 和PD 在表層分別與SWC 顯著正相關;在中層分別與TN 顯著正相關且與C/N 顯著負相關;在底層均與9 個理化因子無顯著相關性。Shannon 和Simpson 在表層與呈顯著負相關,且Simpson 還與C/N 顯著正相關;在中層分別與C/N 呈顯著負相關;在底層,Simpson 與pH 顯著負相關(P<0.05),而Shannon 與9 個理化因子均無顯著性相關。
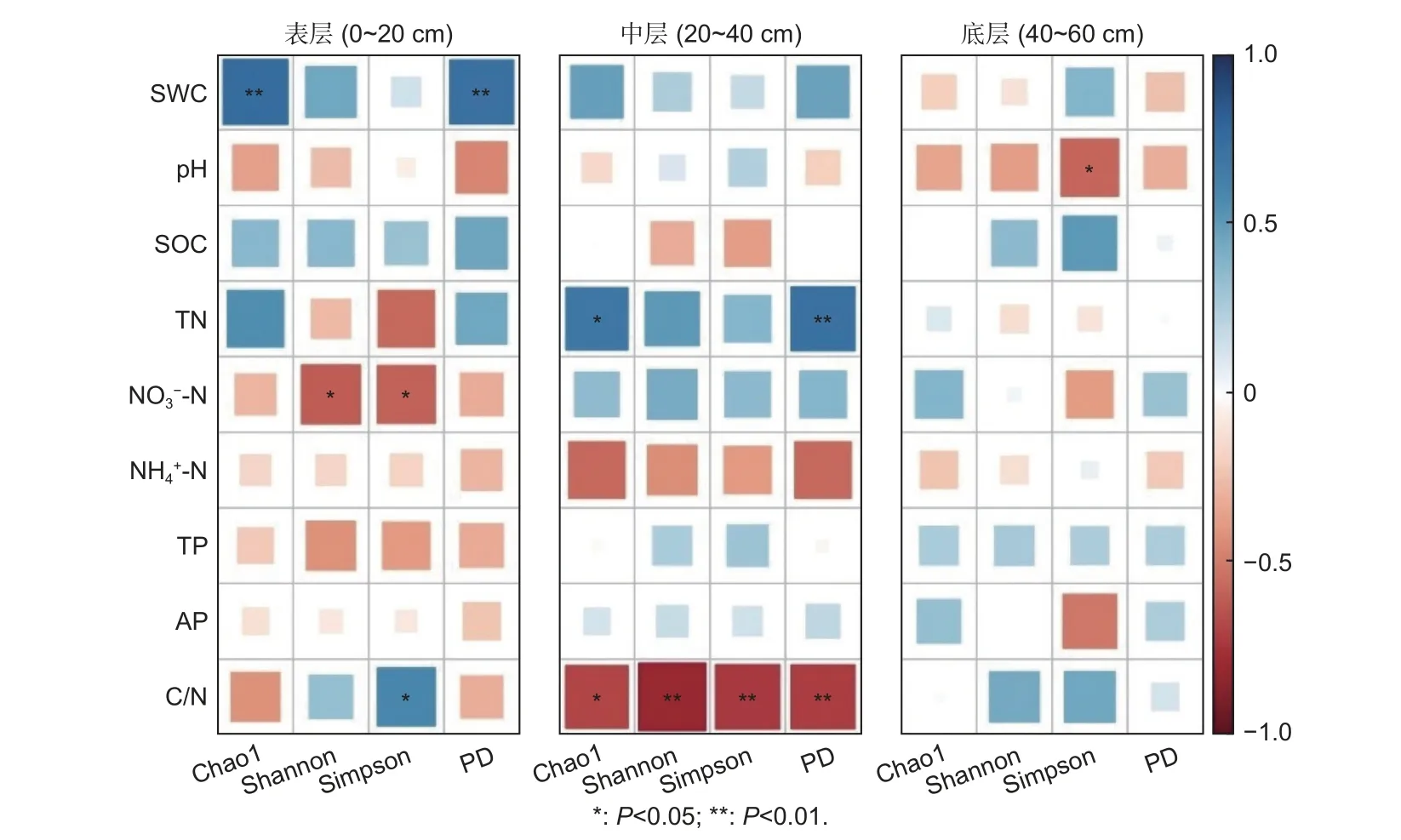
圖2 全部研究林分土壤理化性質與細菌群落分類及系統發育多樣性的相關性熱圖Fig.2 Correlation heatmap of the soil physicochemical properties and bacterial community diversity(taxonomic and phylogenetic) in all plantations
2.3 馬尾松針葉林與鄉土闊葉林土壤細菌群落結構的變化
2.3.1 細菌群落分類結構 在土壤表層,主坐標分析(PCoA)細菌群落分類結構總解釋度為54.4%(圖3),PERMANOVA 分析顯示馬尾松林分別與紅錐林(Fpseudo=3.1,P=0.017)、米老排林(Fpseudo=2.9,P=0.025)、火力楠林(Fpseudo=2.0,P=0.039)有顯著差異。dbRDA 結果表征(圖4),前兩軸累計解釋了58.2% 的樣品細菌群落分類結構與土壤理化性質因子的關系;馬尾松林土壤樣品位于RDA2 的負方,而紅錐林、米老排林和火力楠林土壤樣品幾乎位于RDA2 的正方;Monte Carlo 檢驗進一步表明,土壤TP、SWC、TN 是制約馬尾松林與紅錐林、米老排林、火力楠林土壤細菌群落分類結構差異的環境因素(P<0.05)。
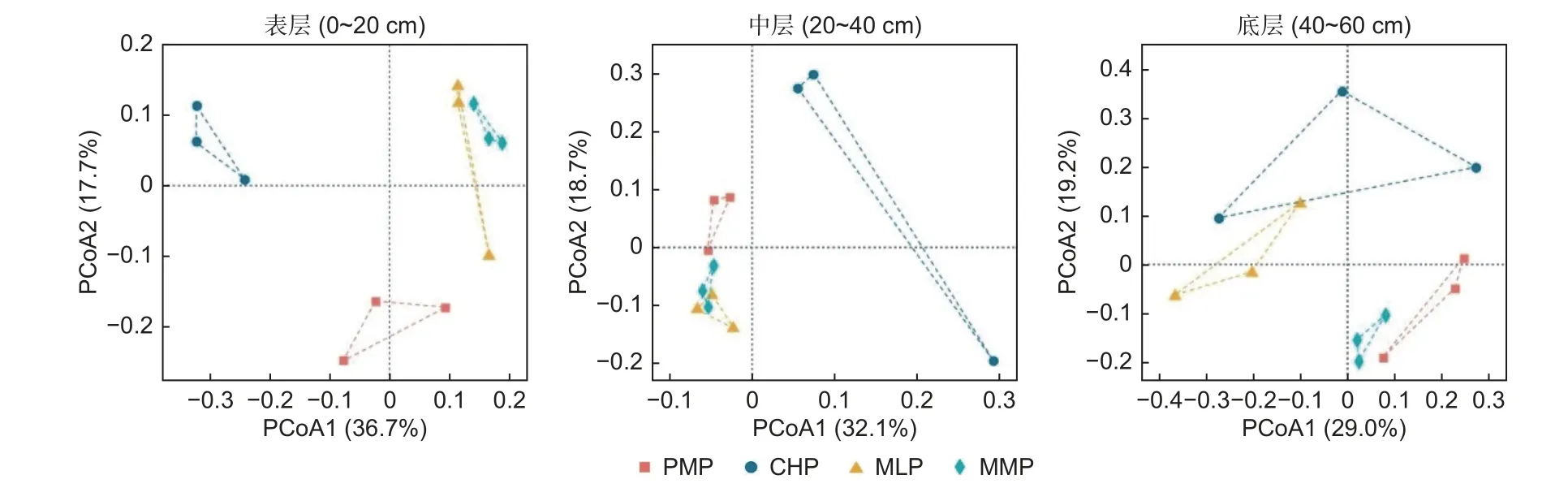
圖3 馬尾松林與3 個鄉土闊葉林土壤細菌群落分類結構的主坐標分析(PCoA)Fig.3 Principal Coordinate Analysis (PCoA) of soil bacterial community taxonomic structure between Pinus massoniana plantation and three broadleaved plantations

圖4 土壤理化性質對4 個人工林細菌群落分類結構影響的冗余分析(RDA)Fig.4 Redundancy analysis (RDA) on effects of soil physical and chemical properties on bacterial community taxonomic structure of four plantations
在中層,PCoA1 和PCoA2 的解釋度分別為32.1%和18.7%(圖3),且PERMANOVA 分析表明馬尾松林僅與紅錐林有顯著差異(Fpseudo=2.3,P=0.012)。dbRDA分析表明(圖4),dbRDA1解釋度為36.3%,dbRDA2 解釋度為21.3%;馬尾松林土壤樣品處于RDA1 的負側,而紅錐林土壤樣品處于RDA1 的正側;Monte Carlo 檢驗揭示土壤C/N 是制約馬尾松林與紅錐林土壤細菌群落分類結構差異的環境因素(P<0.05)。
在底層,PCoA 分析的前兩軸累計解釋度為48.2%(圖3),PERMANOVA 發現馬尾松林與3 個闊葉林均無顯著差異(P>0.05)。dbRDA 前兩軸的總解釋度為51.3%(圖4),Monte Carlo檢驗表明土壤和SWC 是影響不同土壤樣品細菌群落分類結構差異的主要因素(P<0.05)。
另外,相同土層相同林分的兩兩土壤樣品之間的βRC均為-1,意指環境過濾在4 個人工林不同土層的細菌群落構建中均起主導作用。
2.3.2 細菌群落系統發育結構 4 個林分在各土層的細菌群落凈親緣關系指數NRI(圖5A)和最近親緣關系指數NTI(圖5B)均大于1.96,表現為系統發育聚集,說明確定性過程在土壤細菌群落構建中起重要作用,環境過濾導致系統發育聚類。
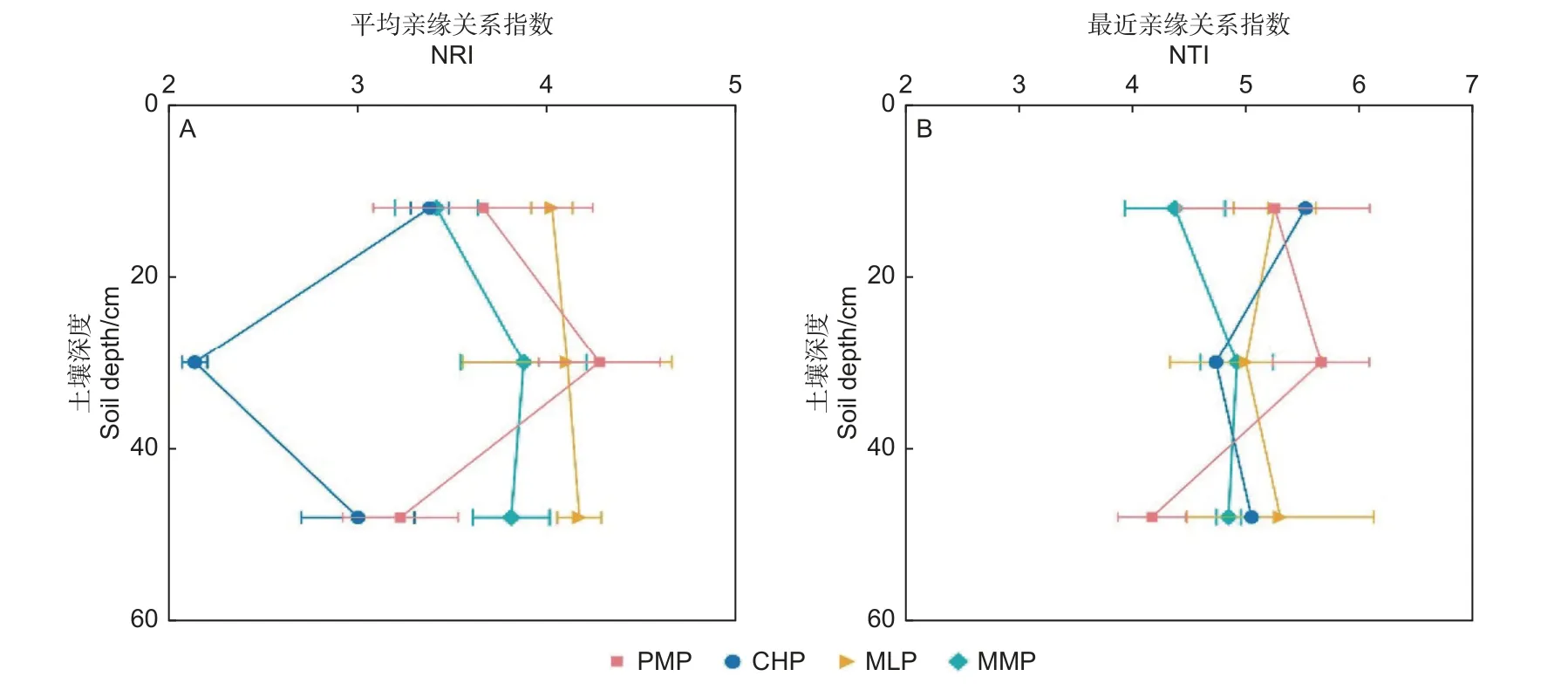
圖5 4 個人工林在不同土壤深度的細菌群落系統發育結構指數(均值 ± 標準誤,n=3)Fig.5 Phylogenetic structure index of bacterial communities in the four plantations at different soil depths (mean ± SE, n=3)
2.4 馬尾松林改建成鄉土闊葉林的土壤細菌群落分類與系統發育轉換率
2.4.1 細菌群落分類轉換率 馬尾松林轉換為3 個闊葉林的土壤細菌群落分類轉換率的差異因土層不同(圖6A)。在0~40 cm 土層,馬尾松林轉換為紅錐林的轉換率顯著高于轉換為米老排林和火力楠林;在40~60 cm,馬尾松林轉換為紅錐林和米老排林的轉換率均顯著高于轉換為火力楠林(P<0.05)。
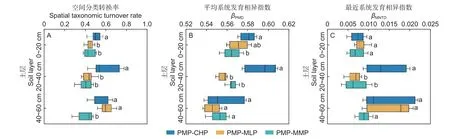
圖6 馬尾松林轉換為3 闊葉林的土壤細菌群落分類和系統發育轉換率Fig.6 Taxonomic and phylogenetic turnover rates of soil bacterial communities from PMP to three broadleaved plantations
2.4.2 細菌群落系統發育轉換率 土壤細菌群落平均系統發育相異指數βMPD(圖6B)和最近系統發育相異指數βMNTD(圖6C)的方差分析表明,在土壤表層,馬尾松林轉換為紅錐林的βMPD顯著大于轉換為火力楠林(P<0.05),而馬尾松林分別轉換為3 個闊葉林的βMNTD無顯著差異(P>0.05);在中層,馬尾松林轉換為紅錐林的βMPD和βMNTD都顯著大于轉換米老排林和火力楠林(P<0.05);在底層,馬尾松林轉換為3 個闊葉林的βMPD、βMNTD均無顯著差異。
3 討論
3.1 針葉林與闊葉林土壤細菌群落多樣性
土壤細菌群落分類多樣性是表征細菌在生態系統中功能的重要指標[27],分類多樣性越高,則越有利于土壤的可持續利用和抵抗力[28]。有研究指出,樹種變化是影響土壤細菌群分類多樣性的原因[29-30]。由于針葉與闊葉樹種的凋落物和根系對土壤傳輸的養分不同,改變了土壤理化性質,從而影響土壤細菌群落分類多樣性。本研究發現,米老排林和火力楠林相比于馬尾松林,土壤表層的細菌群落Chao1 指數有顯著提高(圖1A),而中層和底層土壤細菌群落的分類多樣性(Chao1、Shannon 和Simpson 指數)沒有明顯差異,說明樹種轉換導致土壤細菌群落分類多樣性的變化也會因土壤深度而異。米老排林和火力楠林表層土壤細菌群落Chao1 指數高于馬尾松林,其原因在于其表層土壤SWC 顯著高于馬尾松林(表2),而表層土壤細菌群落Chao1 指數與土壤SWC 顯著正相關(圖2A)。細菌在相對較高的水勢環境下活性更高[31],SWC 高的土壤能夠為土壤細菌的生長提供了一個穩定而潮濕的環境促進其生長繁殖。
系統發育多樣性可以表示物種的差異性和生態位[32],系統發育多樣性越高的生態系統越穩定,且群落中的細菌相互作用越弱[33]。細菌以非常小的尺度(very small scale)棲息在土壤生態位中,其系統發育多樣性通常與土壤理化性質密切相關[34]。本研究發現,系統發育多樣性與分類多樣性呈相同的分布規律,即相較于馬尾松林,米老排林和火力楠林表層土壤細菌群落系統發育多樣性有顯著增加(圖1D),而中層和底層無明顯變化。Taketani等[35]研究指出,土壤SWC 對細菌群落系統發育多樣性具有至關重要的作用,土壤SWC 降低會直接引起系統發育多樣性減少。本研究也發現4 個林分土壤表層SWC與PD極顯著正相關(P<0.01,圖2A),因此,于馬尾松林而言,米老排林和火力楠林表層土壤SWC 的顯著增加是導致細菌群落系統發育多樣性顯著提高的主要緣由。盡管土壤pH 值被普遍認為是影響土壤細菌群落系統發育多樣性的重要因素[36],但由于本研究中紅錐林、米老排林、火力楠林和馬尾松林表層土壤的pH 值無顯著差異(3.88~4.15),因而未發現土壤pH 值對土壤細菌群落系統發育多樣性產生明顯影響。
3.2 針葉林與闊葉林土壤細菌群落結構
針葉林與闊葉林的土壤細菌群落分類結構不同[30],這是多種環境因子綜合作用的結果。Gunina等[37]研究表明,造林樹種不同導致其土壤理化性質差異,最終影響其土壤細菌群落分類結構。土壤TN[38-39]、TP[40]和C/N[41]是調節森林土壤細菌群落分類結構的重要因素。本研究的RDA 分析結果較好的反映了理化因子與土壤細菌群落分類結構的關系表層土壤SWC 增加、TN 增加和TP 減少是驅動馬尾松林分別與3 個闊葉林土壤細菌群落分類結構具有顯著差異的重要因素;而中層土壤C/N 增加是導致馬尾松林與紅錐林土壤細菌群落分類結構差異顯著的主要因子;但在底層馬尾松林與3 個闊葉林土壤細菌群落分類結構都沒有顯著差異(圖3、圖4)。可見,馬尾松林與3 個鄉土闊葉林土壤細菌群落分類結構的差異因土層而異,這是因為人工林凋落物累積在土壤表層,且針葉林與闊葉林的凋落物分解率不同[42],表層土壤理化性質差異較大,但隨著土層加深,針葉林與闊葉林的理化性質差異縮小,使得細菌群落分類結構逐漸趨同。
微生物群落構建過程一直是群落生態學關注的熱點[43],然而由于多種生態過程的作用使因果關系的確定復雜化。βRC可以作為定量估計隨機與確定性過程在微生物群落構建中貢獻的指數[44],系統發育結構的變化也已被證明可以用于評估微生物群落構建過程中環境過濾、競爭排斥和隨機過程的重要性[45]。本研究的βRC、NRI 和NTI 指數表明,馬尾松林與3 個闊葉林不同土層的土壤細菌群落構建均受環境過濾主導,這表明針葉與闊葉樹種的更替不會造成土壤細菌群落構建機制的改變。這與華北亞高山林線區域土壤[11]、不同林齡的杉木人工林土壤(0 ~10 cm)[13]及不同時間歷史的闊葉糖楓(Acer saccharumMarshall)天然林土壤有機質層[46]細菌的研究結果一致,認為是環境過濾調控細菌群落構建。此外,在土壤表層和中層,馬尾松林轉換為紅錐林的物種組成和系統發育結構轉換率高于轉換為米老排林和火力楠林(圖6),本文推測未來將馬尾松林改造成紅錐林后的土壤細菌種類更替比轉換成米老排林和火力楠林更頻繁,紅錐林的土壤細菌群落穩定性較差,細菌間的相互作用更強烈[33]。
4 結論
本文推測南亞熱帶地區的馬尾松林改建成闊葉林后,米老排林和火力楠林土壤表層細菌群落的分類與系統發育多樣性顯著提高了20.5%~23.8%,土壤SWC 是影響的主導因子;3 個闊葉林土壤細菌群落分類結構都在表層產生了顯著的變化,而在中層只有紅錐林有明顯改變,主要受土壤SWC、TP、TN 和C/N 影響;不同土層的細菌群落構建主導因素沒有發生改變,均是環境過濾;在土壤表層和中層,馬尾松林轉變為米老排林及火力楠林之間的分類和系統發育轉換率都低于轉變為紅錐林,米老排林和火力楠林的土壤細菌群落比紅錐林更穩定。基于土壤細菌群落分類與系統發育的多樣性及轉換率角度,該地區將馬尾松林改建成米老排林或火力楠林比紅錐林更能提高土壤細菌群落的穩定性。未來可開展針葉林轉換鄉土闊葉林后土壤細菌群落功能變化的研究,有助于深入理解林分轉換介導土壤生態功能改變的調控機制。
猜你喜歡
西北民族大學學報(自然科學版)(2021年4期)2021-12-29 02:54:24
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
小聰仔(科普版)(2020年12期)2021-01-18 09:16:52
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
學生天地(2019年32期)2019-08-25 08:55:22
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
小天使·一年級語數英綜合(2017年11期)2017-12-05 18:49:56
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
