甜味覺受體在2型糖尿病發病及中醫藥作用研究進展*
2024-01-22 08:15:30孫鐵鋒戰麗彬臧凝子杭天怡朱連連
世界科學技術-中醫藥現代化 2023年8期
孫鐵鋒,戰麗彬,臧凝子,杭天怡,朱連連
(1.南京中醫藥大學中醫學院·中西醫結合學院 南京 210023;2.遼寧中醫藥大學中醫藥創新工程技術中心 沈陽 110847;3.遼寧中醫藥大學中醫臟象理論及應用教育部重點實驗室 沈陽 110847;4.遼寧中醫藥大學附屬醫院 沈陽 110032)
2 型糖尿病與日常飲食密切相關,隨著患者數量的增加,2 型糖尿病已經成為一個全球性的健康問題。保持能量平衡是治療糖尿病的基本要求,能量新陳代謝始于進食。研究表明,消化過程中的氨基酸、脂肪酸和葡萄糖等營養物質作用于胃腸道的神經內分泌細胞,從而在能量平衡中發揮重要作用[1]。因此,胃腸道現在被認為是營養信號的傳感器系統。已有研究表明,味覺受體在腸道中起甜味感受器的作用,并通過分泌胰島素在感受糖和維持血糖動態平衡方面起著關鍵作用[2-3]。這些主要包括味覺受體1 型(Taste receptor type 1,T1R)家 族的G 蛋 白 偶 聯 受 體(G protein-coupled receptors,GPCRs)的異二聚體,用于感知甜味(T1R2-T1R3)和鮮味(T1R1-T1R3)刺激,T2R家族用于感知苦味刺激。近年來,有研究揭示了胃腸道存在額外的營養感知機制[4-5],葡萄糖傳感是通過腸道內分泌細胞(主要是腸道L 細胞)上的甜味(T1R2-T1R3)異源二聚體實現的,其在觸發胰島素激素的分泌以改善血糖和血脂控制方面發揮作用。胃腸感覺機制通過釋放作用于迷走神經和腸道傳入的胃腸激素來調節對營養的生理反應,特別是飽腹感和能量平衡,從而將神經信號傳遞到大腦。使用特定的食物、膳食補充劑或藥物化合物調節可能對治療T2DM 具有重要意義。
1 甜味覺受體的生物學特征
甜味,與如苦、咸、酸等口感一樣,是人類的一種感知,使人類非常愉悅,是大多數哺乳動物的首選。蔗糖、葡萄糖和果糖等糖是典型的甜味化合物,人類認為各種各樣的天然和合成化合物也具有甜味[6]。天然甜味劑來自包括糖、糖醇(如甘露醇、木糖醇)、蛋白質(如他馬汀、單胞素)和氨基酸等化學物質。合成甜味劑種類多樣:常見的甜味劑包括磺酸鹽(如磺素鈉)、二肽(如阿斯巴甜、新甜)、鹵代糖(如三氯蔗糖)和磺酰胺(如糖精鈉、乙酰胺鉀)[7]。
自從在舌頭上發現甜味覺受體(T1R2/T1R3)以來(其中1999 年發現T1R2[8],隨后發現T1R3[9]),已經證明T1R2/T1R3和味覺信號轉導分子不僅在口腔[10]中表達,還在胃腸道[11]、胰腺[12]、膀胱[13]、脂肪組織[12]和大腦[14]等多種非味覺器官中表達。盡管甜味覺受體在多種組織中的重要生理作用仍未確定,但已證實甜味覺受體在胰腺和胃腸道組織中表達與葡萄糖感知、葡萄糖轉運蛋白的表達以及維持葡萄糖穩態有關[11-12]。甜味覺受體是一類7 次跨膜的GPCRs,表達于味蕾上的味覺受體細胞(Taste receptor cell,TRC),包含1個大的細胞外氨基末端或“N”末端結構域、富含半胱氨酸的結構域和7個螺旋穿膜部位3個結構域[15]。
2 甜味覺受體的信號傳導機制
在口腔中,甜味化合物激活味蕾中的甜味感受器[16],誘導細胞內轉導級聯,導致味覺感受器細胞去極化(圖1),在味蕾中,活化的T1R2/T1R3 與α-Gustducin[17]、Gβ3和Gγ13的異三聚體G蛋白相互作用。G蛋白亞基解離后,Gβ3、Gγ13 與磷脂酶C-β2(PLC-β2)相互作用,PLC- β2 進 而 將 磷 脂 酰 肌 醇4, 5 二 磷 酸(phosphatidylinositol(4,5)bisphosphate,PIP2)分裂成肌醇1,4,5-三磷酸(inositol 1,4,5-trispoosphate,IP3),IP3通過IP3 受體(IP3-R)激活刺激Ca2+從內質網釋放。細胞內Ca2+升高激活TRPM5 通道,導致膜去極化,使ATP通道Pannexin-1(PX1)打開并釋放ATP,刺激傳出神經纖維。在腸道中,葡萄糖刺激甜味覺受體,觸發胰高血糖素樣肽-1(Glucagon-like peptide 1,GLP-1)和促胰島素釋放肽(Glucose-dependent insulinotropic polypeptide,GIP)的分泌,并增加鈉-葡萄糖共轉運體-1(sodium-glucose cotransporter 1,SGLT-1)的 表 達。在β 細胞中,葡萄糖通過葡萄糖轉運體-2(Glucose transporter-2,GLUT-2)運輸,ATP 增加,使KATP 通道關閉,從而去極化[18-21]。這種去極化反過來激活電壓依賴的鈣通道,導致Ca2+在細胞質中積累和胰島素分泌。甜味覺受體被認為與胰島素分泌的調節有關。
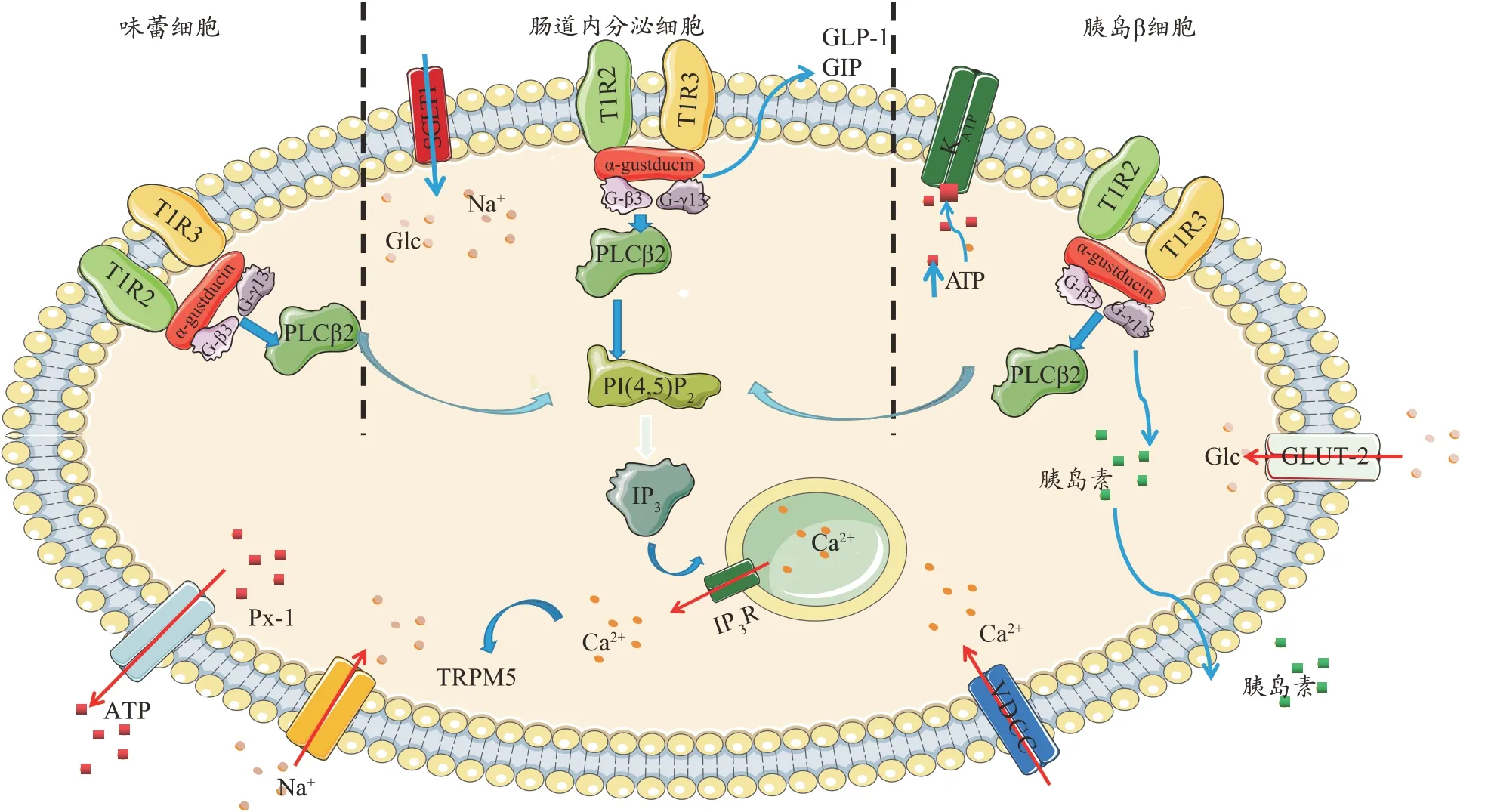
圖1 甜味覺受體的信號傳導機制
3 甜味覺受體與T2DM
目前,全球約有1/11 的成年人患有糖尿病,其中90%為T2DM,且每年人數呈上升趨勢[22-23]。據預測,2030年全球糖尿病患者人數將上升至5.78億;2045年將上升至7 億[24]。T2DM 患者經常表現出對“營養”胃腸道異常反應。據報道,腸道甜味受體T1R2 和T1R3在人類近端腸道的不同上皮細胞中表達,并且轉錄水平隨著T2DM 患者的血糖水平而變化[25]。在嚙齒動物中,在急性暴露于葡萄糖或人工甜味劑的情況下,腸道甜味覺受體轉錄和蛋白水平會迅速下調[26]。在健康人群中,根據血糖狀態,腸道T1R2 的表達受到管腔葡萄糖的相互調節,但在T2DM 患者中,在急性高血糖時,T1R2 的表達是紊亂的[27]。這一缺陷可能會增強T2DM 患者的葡萄糖吸收,并加劇餐后高血糖。最近,有令人信服的證據表明,T1R3 參與了糖脂代謝的調控,并控制了胰島素和胰島素的產生,以T1R3 基因敲除小鼠為研究對象,以親本C57BL/6J 株為對照,研究了T1R3對胰島形態特征的影響[28]。在T1R3基因敲除的動物中,胰島的大小和密度在胰腺組織中都有所減少。所獲得的數據表明,編碼甜味覺受體蛋白的功能基因的缺失導致胰島組織營養不良,并與人類T2DM和肥胖癥所特有的胰腺病變的發生有關。同時T1R2基因敲除實驗證實,這些效應是通過調節GLUT2轉運體轉運到腸上皮細胞的頂膜而特異性地介導的。GLUT2 易位和葡萄糖轉運依賴于胰高血糖素樣肽2(Glucagon-like peptide 2,GLP-2)的分泌和隨后的腸神經元激活。最后,野生型小鼠喂食高蔗糖后,腸道中甜味覺受體的表達迅速下調,導致葡萄糖吸收減少。Campbell 等[29]的研究表明甜味覺受體通過調節葡萄糖的轉運來調節葡萄糖的吸收,并防止由于攝入高糖而加劇高血糖的發展。
3.1 甜味覺受體與胃腸激素
作為機體最大的內分泌系統,胃腸道分泌的激素在維持代謝穩態、調節胃腸道內外組織功能中發揮重要作用。最近在胃腸道中發現味覺受體及其下游信號蛋白,而且食物的攝入可以通過與特定味覺受體結合的化合物產生的胃腸刺激來調節。這些受體在腸道營養的功能檢測中發揮作用,這可能反過來啟動激素或神經級聯通路[30]。胰島素肽,主要是GLP-1 和GIP,調節胰島激素分泌、葡萄糖濃度、脂代謝、腸道動力、食欲和體重以及免疫功能。GLP-1 和GIP 受體的激活還通過直接作用于表達胰島素受體的組織,以及通過神經和內分泌途徑介導的間接機制,在多種組織中導致非血糖效應[29]。
3.1.1 膽囊收縮素
膽囊收縮素(Cholecystokinin,CCK),多肽激素,由33 個氨基酸組成。全部生物活性存在于C 端的八肽片段,CCK 是一種由腸內分泌細胞釋放的腸道激素[31]。Yoshida 等[32]重點研究了CCK 在味覺刺激初始反應中的作用,CCK 與Ⅱ型味覺細胞標志物如α-gustducin、PLCβ2 和TRPM5 共表達;此外,小部分表達CCK 的味覺細胞(Ⅱ型味覺細胞))在味蕾中表達甜味/鮮味味覺受體成分,因為Ⅱ型味細胞是甜味、鮮味或苦味細胞,所以大多數表達CCK 的味細胞可能是苦味細胞。總的來說,CCK 通過作用于迷走神經傳入神經和下丘腦表達的膽囊收縮素受體,以及瘦素之間的協同相互作用,起到控制飲食。
3.1.2 胰高血糖素樣肽-1
在健康人中,胰高血糖素樣肽-1(Glucagon-like peptide 1,GLP-1)是在進食后分泌的,通過增加胰島素分泌和抑制胰升糖素釋放來降低葡萄糖濃度。GLP-1的其他作用包括延緩胃排空、抑制食欲,并可能抑制β細胞的凋亡。天然的GLP-1 在循環中約2-3 min 內被降解;因此,各種GLP-1 受體激動劑已被開發出來,以提供更久的體內作用[33]。黃精花多糖[34]通過甜味覺受體介導的cAMP 信號促進腸內分泌L 細胞的GLP-1 分泌。干預實驗表明,黃精花多糖刺激的GLP-1 分泌不僅受到抗體、siRNA 和T1R2/T1R3抑制劑的顯著抑制,而且還受到腺苷酸環化酶抑制劑的抑制。這些結果提示黃精花多糖可能通過激活T1R2/T1R3 介導的cAMP信號通路刺激腸內分泌細胞分泌GLP-1。
3.1.3 YY肽
YY 肽(Peptide tyrosine tyrosine,PYY)是小腸細胞分泌的一種激素,這種激素與進食直接相關,飯后釋放到血液中,與大腦中的受體結合,使人感到飽足;PYY也在消化道發揮作用,有助于控制消化過程的速度,哺乳動物味蕾細胞表達多種肽的受體,這些肽在調節新陳代謝、營養同化和飽腹感方面與身體其他部位有關[35]。PYY存在于唾液和味覺細胞亞群中,可以影響行為味覺反應,減少食物攝入和體重。采用基質膠誘導NCIH716細胞建立小腸內分泌模型[36],結果表明海藻糖、赤蘚糖醇、山梨糖醇可通過激活T1R2、T1R3、α-Gustducin、PLCβ2、TRPM5等基因,增加胞內Ca2+濃度,以激活甜味覺受體信號通路促進GLP-1、PYY分泌。
3.1.4 血清素
血清素(Serotonin,5-HT),廣泛分布于人體,尤其是大腦皮層和突觸。大腦中的5-HT 控制著人類的飲食行為、認知功能和情緒調節[37-38]。共有27 名健康男性受試者接受了甜味覺受體抑制劑-2-(4-甲氧基苯氧基)-丙酸鈉干預實驗。在120 min 內測定了CCK、胃饑餓素(Ghrelin)、5-HT 的餐后血漿濃度以及體溫的變化。本研究表明,甜味覺受體抑制劑能增加能量攝入,降低血漿5-HT濃度[39]。
3.2 甜味覺受體與腸道菌群
脾為后天之本,腸道在中醫學藏象理論體系中隸屬于脾胃之范疇,而腸道菌群居于腸中,其作為龐大而復雜的生態系統,對宿主能量代謝、營養吸收、免疫功能等方面起到重要作用[40],與中醫對脾主運化、氣血生化之源等認識高度契合。研究表明,除環境和遺傳因素外,胃腸道微生物群在T2DM 和肥胖的發生發展中起著重要的作用。目前,相關研究主要關注的是遠端腸道菌群,而忽略了同樣重要的近端腸道菌群,如十二指腸中的菌群。有研究定性分析了十二指腸菌群的結構,并與相關臨床指標做了關聯分析,雙歧桿菌屬細菌可能是T2DM 和肥胖發生發展的生物標志物[41]。甜味覺受體在口腔和胃腸道都有表達,參與食欲、葡萄糖穩態和腸道運動的調節。因此,甜味覺受體可能與胃腸道微生物群相互作用,這種相互作用可能解釋T2DM、肥胖和代謝結果之間的關系。
舌上皮和腸上皮所共有的化學感覺成分在食物消耗和腸道功能調節中起著至關重要的作用。除營養信號外,包括微生物在內的其他管腔內容物在整個胃腸黏膜的信號傳遞和啟動消化功能的改變方面也很重要。腸道微生物區系在影響食物攝入量、能量平衡和體重增加方面的潛在作用已被提出[42-44]。然而,腸道微生物區系是否調節營養反應受體和轉運蛋白的表達,從而導致食物消費的改變,尚不清楚。Swartz等[45]用偏好實驗比較了無菌(GF,C57BL/6J)小鼠與常規(CV,C57BL/6J)對照組小鼠對營養性(蔗糖)和非營養性(糖精)甜味劑的偏好。定量了T1R3、α-Gustducin 和SGLT-1 在CV 和GF 小鼠腸上皮中的基因和蛋白表達,還包括舌上皮中T1R2、T1R3 和α-Gustducin 的基因表達。結果發現,盡管兩組小鼠對蔗糖的偏好相似,但GF小鼠比CV小鼠攝入更多高濃度(8%)的蔗糖溶液,糖精的攝入量和偏好均無差異。與對照組相比,GF 組小鼠腸上皮細胞T1R3 和SGLT-1mRNA 和蛋白的表達明顯增加,而舌味覺受體mRNA 的表達差異無統計學意義。因此腸道微生物區系的缺失改變了近端小腸甜味覺受體的表達和過量供應,這與營養甜味劑的攝入量增加有關。有研究者使用豚鼠模型來研究糊精(SS)在微生物群-腸道-下丘腦軸中的生理作用[46]。飲水中添加1.5 mmol·L-1SS 可提高血清葡萄糖水平,糖精鈉處理激活了回腸的甜味覺受體信號。在SS 和對照(CN)組的下丘腦中,RNA-seq 鑒定了1370 個不同表達的基因(796 個上調,574 個下調),富集到味覺信號轉導和神經活性配體-受體相互作用中,SS處理顯著增強了回腸微生物群發酵的功能。總之,SS 激活了腸道中的甜味覺受體信號以及腸道菌群。
3.3 甜味覺受體與胃腸免疫
甜味覺受體最近被證實參與越來越多的免疫過程,在胃腸道免疫方面,有學者發現攝入脂多糖后,蔗糖敏感性降低,同時T1R2 和T1R3 轉錄物的舌側表達降低,被翻譯成形成甜味覺受體的T1R2+T1R3 亞單位[47]。在LPS 被消耗后,T1R2 和T1R3 mRNA 表達水平和對蔗糖的神經反應得以恢復。缺乏脂多糖Toll樣受體(Toll-likereceptor,TLR)的小鼠對脂多糖攝入后的神經生理變化有抵抗力。這些發現表明,在單一時期攝入脂多糖會特異性地暫時抑制對蔗糖的神經反應。綜上,LPS 會啟動依賴TLR4 的激素信號,下調味蕾中的甜味覺受體基因。甜味信號延遲抑制可能會影響食物選擇以及胃腸道細菌和肥胖之間復雜的相互作用。胃腸道上皮中表達的先天性免疫受體可感知入侵的病原體,先天性免疫受體信號通路的失調與炎癥疾病相關。Kayisoglu等[48]通過構建人與小鼠的胃腸道不同部位的類器官,并利用全基因組RNA 測序,發現胃腸道上皮的先天性免疫應答信號主要受到位置(而非菌群)的影響。另外,對Toll 樣受體4(TLR4)在胃腸道上皮中的表達與功能進行分析發現,免疫相關基因在胃腸道中的表達及功能在發育期間已被決定,且不受環境影響。
4 甜味覺受體與“脾在味為甘”
戰麗彬教授多年來從事脾藏象及從脾論治代謝性疾病的相關研究[49],認為“脾虛痰濕”為肥胖T2DM之中醫關鍵病機,治則當以“健脾化濕”。《內經》曰“五味所入……甘入脾”,“脾在味為甘”是中醫脾藏象的重要理論內容,是脾臟生理、病理、治法要點的理論基礎。①生理上,脾主運化,主氣太陰濕土,而甘味屬土,與脾相配,在運化之水谷精微之中,脾最喜甘味;②病理上,甘味最易傷脾,而致中滿,日久化熱,傷及脾陰,而致消渴;③治療上,甘味藥多能補養脾胃中氣,喻嘉言云“脾胃者土也,土雖喜燥,然太燥則草木枯槁,土雖喜潤,然太濕則草木濕爛,以補滋潤之劑,使燥濕相宜”,即補脾胃者,當使潤燥相濟,以甘味為主,其中甘不僅指藥食甜之口味,更指其和緩、補養、潤燥之功效特點。
李杰[50]研究黃芩-黃連藥對高糖高脂誘導的糖尿病前期大鼠以及3-脫氧葡萄糖醛酮(3DG)損傷的INS-1 細胞胰島素分泌的改善作用,結果發現與對照組相比,模型組糖尿病前期大鼠胰腺組織中T1R2、T1R3、PLCβ2 以及TRPM5 蛋白表達顯著降低;經黃芩-黃連藥對干預后,與模型組相比,黃芩-黃連組大鼠胰腺組織中T1R2、T1R3、PLCβ2以及TRPM5蛋白表達顯著增加。楊海梅[51]研究半夏-黃連對糖尿病前期大鼠模型GLP-1 水平與STC-1 細胞GLP-1 分泌受損的改善作用,并揭示半夏-黃連對甜味受體信號通路的調控作用。同時對半夏-黃連改善GLP-1 分泌的一些潛在機制進行探究。半夏-黃連處理后T1R3、PLCβ2、TRPM5 蛋白表達增加;另外,糖尿病前期大鼠T1R3、PLCβ2、TRPM5 蛋白的表達下降,半夏-黃連干預后蛋白表達量增加。劉佳[52]的實驗表明,健脾法可影響腸道甜味覺受體及關鍵信號分子表達并干預TlRsα-Gustducin/PLCβ2 味覺通路的轉導。陳龍輝[53]的研究發現脾虛與甜味覺受體密切相關,實驗結果證實干預甜味覺受體可以恢復脾的功能。因此從甜味覺受體研究脾,有助于揭示“脾在味為甘”的理論內涵,也將有助于闡明中醫從脾論治T2DM 有著現實的研究指導意義。
5 討論
T2DM 的患病率持續增加,找到有效策略來調節血糖水平是至關重要的。盡管空腹血糖水平已被用作血糖管理的指標,但控制餐后血糖,即降低血糖峰值和波動,是血糖管理的關鍵。維持血糖在一個特定范圍內被稱為葡萄糖穩態。除限制碳水化合物外,改變直鏈淀粉與支鏈淀粉的比例或增加膳食脂肪和纖維含量的比例,這些方法可以降低碳水化合物的消化率,改變腸道平衡(包括胃腸激素、腸道菌群、胃腸免疫),促進葡萄糖從血液中轉運刺激GLP-1 的分泌(見圖2)。尋求優化餐后葡萄糖穩態的一個新興研究領域涉及甜味刺激,這種策略的前提是,口腔中或者腸道中的甜味覺受體已被證明觸發生理反應,為即將攝入的碳水化合物準備身體并調節葡萄糖穩態。在除口腔組織中,T1R2/T1R3 已被證明參與調節代謝過程,如糖感應、葡萄糖穩態和飽足激素釋放。T1R2/T1R3 在腸道和胰腺中的功能代表了利用甜味覺受體作為潛在治療靶點可能。越來越多的證據表明,甜味感受器在全身無處不在,包括胃腸道和下丘腦[54]。這些甜味覺受體在很大程度上參與營養感知,監測能量儲存的變化,并觸發代謝和行為反應,以維持能量平衡。這些途徑受到外部和內部因素的調控,其中一個或多個通路的功能障礙可能引起肥胖和T2DM。
T2DM 和肥胖是一個全球性的公共衛生問題[55]。目前的治療集中在減肥和整體飲食質量控制上。然而,對于飲食的最佳常量營養成分,特別是對于T2DM患者長期管理還需要建立規范[56]。通過對口腔組織、腸道組織等中甜味覺受體的觀察,可以設計出具有特異性和高效的新化合物,甜味覺受體及甜味劑等已經在控制體重和葡萄糖穩態方面發揮重要作用。因此,對甜味覺受體生物學的了解的進展可能有助于設計新的具有良好效果的甜味覺受體激動劑。
