產(chǎn)活性氧假交替單胞菌GCY全基因組序列分析
2024-01-22 08:09:26岳昊,吳碩,王競(jìng)*,李澤龍,顧晨
大連理工大學(xué)學(xué)報(bào) 2024年1期
岳 昊, 吳 碩, 王 競(jìng)*, 李 澤 龍, 顧 晨
(1.大連理工大學(xué) 環(huán)境學(xué)院, 遼寧 大連 116024;2.中國(guó)電建集團(tuán)華東勘測(cè)設(shè)計(jì)研究院有限公司, 浙江 杭州 311122 )
0 引 言

由Gauthier等[11]在1995年從交替單胞菌中分離得到的假交替單胞菌(Pseudoalteromonas),是在全球海洋生態(tài)系統(tǒng)中普遍存在的革蘭氏陰性菌.在本課題組之前的研究中,Gu等[12]證明了Pseudoalteromonassp.GCY可通過(guò)產(chǎn)生的胞外ROS完成四溴雙酚A的好氧共代謝降解;Li等[7]揭示了Pseudoalteromonassp.GCY降解布洛芬首先由胞外ROS啟動(dòng),然后中間產(chǎn)物被細(xì)胞內(nèi)的酶進(jìn)一步降解.然而Pseudoalteromonassp.GCY的參考基因組還不清晰,且影響假交替單胞菌胞外ROS產(chǎn)生的因素也未知.
從大連近海表層沉積物中分離得到一株能產(chǎn)胞外ROS的Pseudoalteromonassp.GCY.采用Illumina測(cè)序技術(shù)對(duì)Pseudoalteromonassp.GCY基因組進(jìn)行測(cè)序,并對(duì)其產(chǎn)ROS特性進(jìn)行研究,以提供Pseudoalteromonassp.GCY(以下簡(jiǎn)稱(chēng)菌株GCY)的基因組信息,明晰影響產(chǎn)ROS的因素.研究結(jié)果將在分子水平上加深對(duì)生物源ROS產(chǎn)生以及影響機(jī)制的理解.
1 材料與方法
1.1 菌株的分離與鑒定
近海表層沉積物采集自中國(guó)遼寧省大連市黑石礁(38°52′34″N,121°33′58″E).連續(xù)培養(yǎng)120 d后,沉積物中的菌群被富集.將富集得到的菌群在人工海水制備的固體BP培養(yǎng)基中25 ℃下培養(yǎng)2 d,然后在培養(yǎng)基上進(jìn)行重復(fù)劃線(xiàn),以純化菌株GCY[12].固體BP培養(yǎng)基成分如下:牛肉膏1 mg/L,蛋白胨2 mg/L,四溴雙酚A 50 mg/L,瓊脂20 mg/L.利用場(chǎng)發(fā)射掃描電子顯微鏡(FEI Nova NanoSEM 450)觀察菌株GCY的形態(tài).
挑取固體BP培養(yǎng)基中的單個(gè)菌落,溶于10 μL無(wú)菌水中,80 ℃水浴變性15 min.以離心上清液為模板進(jìn)行聚合酶鏈反應(yīng)(PCR)擴(kuò)增.TaKaRa 16S rDNA細(xì)菌鑒別PCR試劑盒用于擴(kuò)增菌株GCY的16S rRNA基因,測(cè)序的正向引物和反向引物分別為RV-M和M13-47.?dāng)U增反應(yīng)條件:94 ℃變性5 min,隨后以94 ℃、60 s,55 ℃、60 s,72 ℃、1.5 min為一輪,反應(yīng)30輪,最終在72 ℃下延伸5 min.切割凝膠測(cè)序,回收PCR產(chǎn)物,即獲得菌株GCY的16S rRNA基因序列.DNA測(cè)序部分由寶日醫(yī)生物技術(shù)(北京)有限公司大連分公司完成.Blastn程序用于分析菌株GCY序列與已知的16S rRNA基因序列的同源性.除特別說(shuō)明外,本文提及的所有軟件均使用默認(rèn)參數(shù).OrthoANI用于計(jì)算兩個(gè)16S rRNA基因序列之間的平均核苷酸同源性.Clustal X用于比較相關(guān)序列.使用MEGA11,基于16S rRNA基因,利用鄰近算法推斷菌株GCY的系統(tǒng)發(fā)育樹(shù).
1.2 全基因組測(cè)序和注釋
利用Illumina NovaSeq PE150短讀平臺(tái)和PacBio RSII長(zhǎng)讀平臺(tái)對(duì)菌株GCY的全基因組進(jìn)行測(cè)序.PacBio RSII長(zhǎng)讀取用于基因組組裝,Illumina NovaSeq PE150短讀取用于scaffold組裝.對(duì)低質(zhì)量的reads進(jìn)行過(guò)濾(<500 bp),獲得干凈數(shù)據(jù).對(duì)照Gene Ontology(GO)數(shù)據(jù)庫(kù)、Kyoto Encyclopedia of Genes and Genomes(KEGG)數(shù)據(jù)庫(kù)、Clusters of Orthologous Groups(COG)數(shù)據(jù)庫(kù)和Swiss-Prot數(shù)據(jù)庫(kù)對(duì)基因功能進(jìn)行注釋?zhuān)畬?duì)照綜合抗生素耐藥性數(shù)據(jù)庫(kù)(Comprehensive Antibiotic Resistance Database,CARD)注釋抗性基因.采用SignalP工具預(yù)測(cè)該蛋白序列是否為分泌蛋白.
1.3 ROS產(chǎn)生特性
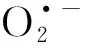
1.4 比較基因組
為進(jìn)一步了解環(huán)境中生物源ROS的分布,首先在假交替單胞菌屬內(nèi)進(jìn)行比較基因組分析,最終以與菌株GCY基因組高度相似的Pseudoalteromonasflavipulchrastrain JG1和Pseudoalteromonaspiscicidastrain JCM 20779為代表.利用Orthovenn對(duì)3個(gè)菌株的蛋白質(zhì)序列比較聚類(lèi),構(gòu)建基因家族.使用Mauve計(jì)算和繪制3個(gè)菌株基因組之間保守和高度同源基因組區(qū)域的共線(xiàn)性.對(duì)照整合微生物基因組(Integrated Microbial Genomes,IMG)數(shù)據(jù)庫(kù)檢索上述3個(gè)菌株的ROS產(chǎn)生和消除相關(guān)基因.此外,利用AnnoTree(http://annotree.uwaterloo.ca/)檢測(cè)胞外ROS產(chǎn)生相關(guān)酶在環(huán)境微生物中的分布.
2 結(jié)果與討論
2.1 菌株GCY鑒定與系統(tǒng)發(fā)育
菌株GCY在瓊脂平板上的菌落與典型的假交替單胞菌相似,呈橘黃色不透明光滑的圓形.該菌是革蘭氏陰性、兼性好氧、異養(yǎng)的動(dòng)桿菌.場(chǎng)發(fā)射掃描電子顯微鏡觀察表明,菌株GCY的平均尺寸為0.4~2.0 μm.16S rRNA測(cè)序結(jié)果顯示,菌株GCY的16S rRNA序列全長(zhǎng)1 447 bp(GenBank accession number KY 583737.1).16S rRNA同源性分析結(jié)果顯示,菌株GCY與Pseudoalteromonaspiscicidastrain NBRC 103038和Pseudoalteromonaspiscicidastrain IAM 12932的關(guān)系最為密切.基于16S rRNA基因序列的系統(tǒng)發(fā)育分析結(jié)果如圖1所示,菌株GCY與Pseudoalteromonaspiscicidastrain NBRC 103038形成進(jìn)化枝.平均核苷酸同源性為100%,這表明它們可能屬于同一種.

圖1 菌株GCY的系統(tǒng)發(fā)育樹(shù)
2.2 菌株GCY基因組特性
經(jīng)過(guò)Illumina和PacBio測(cè)序得到了菌株GCY基因組的結(jié)構(gòu)(圖2).完整的基因組由Chr1和Chr2兩個(gè)染色體環(huán)組成,圖中由外到內(nèi)分別為重復(fù)密度,COG、KEGG、GO基因注釋結(jié)果,ncRNAs,基因組GC含量,基因組GC偏倚值.結(jié)果表明,完整基因組由兩條環(huán)狀染色體組成,大小為5.47 Mb,平均GC含量為43.39%.全部基因總長(zhǎng)為4.84 Mb,占基因組總長(zhǎng)的88.5%.預(yù)測(cè)到4 626個(gè)蛋白質(zhì)編碼基因、106個(gè)tRNA基因、28個(gè)rRNA基因和4個(gè)sRNA基因(表1).氨基酸運(yùn)輸和代謝(E)、翻譯、核糖體結(jié)構(gòu)和生物發(fā)生(J)、信號(hào)轉(zhuǎn)導(dǎo)機(jī)制(T)和轉(zhuǎn)錄(K)是最豐富的COG類(lèi)別.此外,在菌株GCY基因組序列中預(yù)測(cè)到了500個(gè)分泌蛋白.

表1 菌株GCY基因組特性
2.3 基于全基因組的分析


在菌株GCY基因組中還注釋到32個(gè)編碼糖基轉(zhuǎn)移酶的基因,如ftsI、ftsW、murG和rfaQ等,這些基因可能參與了菌株GCY胞外聚合物質(zhì)(extracellular polymeric substances,EPS)的形成[18].產(chǎn)生EPS的菌株在假交替單胞菌屬中很常見(jiàn)[19].研究表明,EPS可以增強(qiáng)細(xì)菌對(duì)氧化應(yīng)激的耐受性,從而減輕膜損傷[20].
此外還在菌株GCY基因組中發(fā)現(xiàn)了adeG、mexI、mexT、mfd、msbA、PmrE、TaeA和TriC等抗生素抗性基因.在過(guò)去的幾年里,抗生素在海洋環(huán)境中被廣泛發(fā)現(xiàn),以往的研究表明,抗生素可引起細(xì)胞氧化應(yīng)激[21-22].通過(guò)CARD分析,推測(cè)菌株GCY的耐藥性主要來(lái)源于外排泵機(jī)制.這種機(jī)制在革蘭氏陰性菌中普遍存在,涉及的基因包括adeG、TriC、mexT和mexI等.這些基因編碼的酶可以泵送多種抗生素,賦予菌株GCY抗生素耐藥性[23-24],同時(shí)減輕細(xì)胞的氧化應(yīng)激.
綜上所述,在菌株GCY基因組中鑒別到使其兼具產(chǎn)胞外ROS能力與應(yīng)對(duì)氧化脅迫能力的相關(guān)基因.
2.4 ROS產(chǎn)生特性分析
2.5 比較基因組分析
為進(jìn)一步了解環(huán)境中生物源ROS的分布,首先在假交替單胞菌屬內(nèi),通過(guò)比較基因組分析了菌株GCY與其他菌株的基因組特征.以與菌株GCY基因組高度相似的Pseudoalteromonasflavipulchrastrain JG1和Pseudoalteromonaspiscicidastrain JCM 20779為代表進(jìn)行基因組特征分析(圖4,表2).如圖4所示,3個(gè)菌株核心基因組為3 737個(gè)基因,占每個(gè)基因組的90.0%~ 92.2%;菌株GCY(4 151個(gè)基因)中分別有94.3%(3 915個(gè)基因)和95.5%(3 963個(gè)基因)在菌株JG1和菌株JCM 20779中具有同源性(圖4(a)),共線(xiàn)性分析結(jié)果與菌株GCY、JG1和JCM 20779的密切親緣關(guān)系一致(圖4(b)),被色塊覆蓋的序列區(qū)域在基因組中完全共線(xiàn)和同源.中心線(xiàn)以下的塊表示按反向補(bǔ)碼(逆)方向?qū)R的區(qū)域.重新排列用彩色線(xiàn)條表示.據(jù)此推測(cè),菌株JG1和菌株JCM 20779的基因組中也存在與胞外ROS產(chǎn)生和消除相關(guān)的基因.這表明與菌株GCY相似的菌株也具有產(chǎn)胞外ROS的潛力.同時(shí),對(duì)3個(gè)菌株基因組中ROS相關(guān)功能基因進(jìn)行檢索驗(yàn)證了這一推測(cè):3個(gè)菌株基因組中都存在編碼LAAO、Na+-轉(zhuǎn)運(yùn)NADH-醌氧化還原酶、超氧化物歧化酶和過(guò)氧化氫酶的基因.有研究發(fā)現(xiàn)假交替單胞菌屬的其他種也能產(chǎn)生L-氨基酸氧化酶[25-26]和過(guò)氧化氫酶.以上分析表明具有產(chǎn)胞外ROS能力的微生物在假交替單胞菌屬內(nèi)廣泛存在.

圖3 ROS的產(chǎn)生特性

(a) 3個(gè)菌株的COGs維恩圖

(b) 全基因組比對(duì)
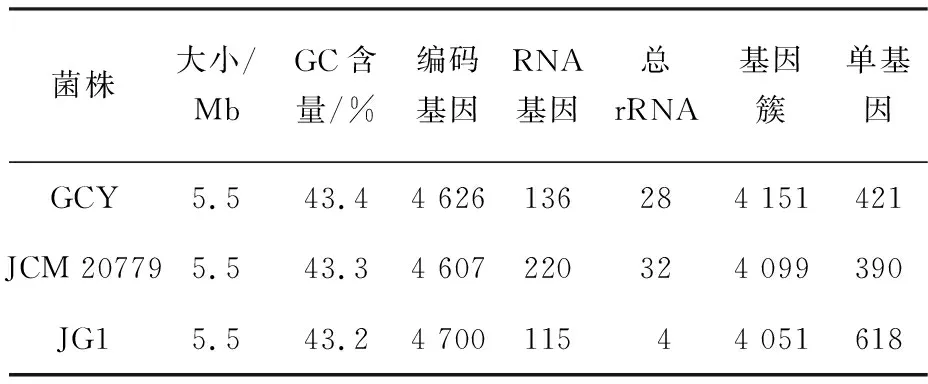
表2 菌株GCY、JG1和JCM 20779的基因組比較
進(jìn)一步利用AnnoTree(http://annotree.uwaterloo.ca/)檢測(cè)LAAO和Na+-轉(zhuǎn)運(yùn)NADH-醌氧化還原酶在環(huán)境微生物中的分布.圖5展示了具有編碼LAAO(圖5(a))和Na+-轉(zhuǎn)運(yùn)NADH-醌氧化還原酶(圖5(b))基因的微生物,除了海洋生物種群外,一些存在于淡水和土壤中的微生物也具有這種能力,例如廣泛分布于土壤、淡水的假單胞菌已被證明產(chǎn)胞外ROS并因此被用于降解四溴雙酚A[27].此外,研究表明與ROS產(chǎn)生相關(guān)的基因通過(guò)可移動(dòng)遺傳元件在物種間的頻繁水平轉(zhuǎn)移有助于它們?cè)诃h(huán)境中廣泛傳播[28],且胞外ROS對(duì)微生物益害并存[29],因此,有理由推測(cè)具備產(chǎn)胞外ROS能力的微生物可能在環(huán)境中廣泛存在.

(a) LAAO

(b) Na+-轉(zhuǎn)運(yùn)NADH-醌氧化還原酶
3 結(jié) 語(yǔ)
本研究提供了具有產(chǎn)胞外ROS能力的Pseudoalteromonassp.GCY的生物學(xué)信息,鑒別了該菌株胞外ROS產(chǎn)生相關(guān)功能基因包括lodA、lodB和nqrA-F,抗氧化應(yīng)激相關(guān)功能基因包括sod和kat等.在此基礎(chǔ)上通過(guò)實(shí)驗(yàn)探明了氨基酸和Na+濃度是影響菌株胞外ROS產(chǎn)量的關(guān)鍵因素:在一定范圍內(nèi),氨基酸和Na+濃度的增大有利于胞外ROS產(chǎn)量的增加.研究還探明了胞外ROS產(chǎn)生相關(guān)基因廣泛存在于環(huán)境中的假交替單胞菌乃至各種微生物中,拓寬了對(duì)生物源ROS存在范圍的認(rèn)識(shí).綜上,本研究結(jié)果在分子水平上加深了對(duì)生物源ROS產(chǎn)生的理解,這為開(kāi)發(fā)利用生物源ROS提供了更多啟示.此外,鑒于ROS參與環(huán)境中多種物質(zhì)轉(zhuǎn)化,推測(cè)生物源ROS可能在無(wú)光區(qū)的生物元素地球化學(xué)循環(huán)中發(fā)揮了重要作用.
