兩種化學型黃樟矮林生長差異及其精油含量分布規律
2024-01-22 00:00:00顧祎昕俞勇熠丁穎魏希肖祖飛
南方農業·上旬 2024年12期




摘 要 為提高不同化學型黃樟矮林精油的利用效率,推動其精油的綜合開發與利用,以2年生芳樟醇型和檸檬醛型黃樟矮林為材料,比較分析其萌芽數、萌條數、萌條長度和粗度、單株鮮質量、單株葉鮮質量、單株小枝鮮質量、單株主枝鮮質量、葉片、小枝和主枝出油率的差異性。結果表明:芳樟醇型黃樟矮林的萌芽數達12.27個,顯著大于檸檬醛型黃樟(8.53個),但在萌條數方面,兩種化學型之間的差異并不顯著;檸檬醛型黃樟矮林的萌條長度為167.03 cm,顯著長于芳樟醇型黃樟(112.37 cm);萌條粗度為28.97 mm,也顯著大于芳樟醇型黃樟(17.98 mm)。檸檬醛型黃樟矮林單株鮮質量、單株葉鮮質量、單株小枝鮮質量和單株主枝鮮質量顯著高于芳樟醇型黃樟矮林,但檸檬醛型黃樟矮林葉/小枝、葉/主枝、葉/枝和(葉+小枝)/主枝顯著小于芳樟醇型黃樟矮林,且生物量大,葉/枝反而小。芳樟醇型黃樟矮林葉片出油率顯著高于檸檬醛型黃樟矮林,小枝、主枝的出油率差異不顯著,兩種化學型黃樟矮林的精油含量均為葉>小枝>主枝。
關鍵詞 矮林;黃樟;化學型;生物量;出油率
中圖分類號:S792.23 文獻標志碼:A DOI:10.19415/j.cnki.1673-890x.2024.23.003
黃樟[Cinnamomum porrectum" (Roxb.)" Kosterm.],又名油樟、大葉樟,屬樟科樟屬高大喬木,自然分布于長江以南各省區,是我國南方重要的材用和特種經濟林樹種[1],其樹體高大,最高可達25 m,胸徑可達100 cm。黃樟材質堅實,防蟲耐腐,有芳香氣味,是高級家具和室內裝修的優良材料[2-3]。黃樟全株各部分都含有精油,因其獨特的化學特性和廣泛的用途,在眾多領域展現出極高的應用價值,而精油的含油量決定了其潛在的應用價值。
近年來,隨著芳樟醇市場需求量的不斷激增,黃樟資源的開發也受到了更多的關注。為在短時間內獲得較高的經濟效益,樟樹采用短輪伐期矮林作業模式,通過縮短采伐周期,提高精油產量。吳航等在廣東省紫金縣地區的野生黃樟種群中鑒定出了7個化學類型[4]。本研究針對芳樟醇型和檸檬醛型黃樟矮林的生長規律、生物量分布規律、精油分布規律進行探討,以期為黃樟矮林的作業提供理論依據。
1" 材料與方法
1.1" 試驗地概況
試驗地位于本中心育苗基地,地理坐標為116°01′19″E、28°41′33″N,屬亞熱帶濕潤季風氣候區,氣候濕潤溫和,光照充足,降雨量充沛,年無霜期長、冰凍期短。年平均氣溫17.1~17.8 ℃,極端最高氣溫達40 ℃以上,極端最低氣溫低于-10 ℃,年日照時數1 772~1 845 h,年降雨量1 567.7~1 654.7" mm,年平均相對濕度為78.5 %。
1.2" 試驗材料
以2年生芳樟醇型黃樟矮林和檸檬醛型黃樟矮林(株距1.0 m、行距1.0 m)為試驗材料。2021年11月份開始伐樁,伐樁高度為30 cm,2022年4月上旬,每株穴施尿素50 g、復合肥50 g,此后按常規進行養護。
1.3" 試驗設計
2022年5月測定留樁上的萌芽數,同年9月在120株無性系樹種中隨機選取芳樟醇型黃樟矮林和檸檬醛型黃樟矮林各12株,將留樁上的萌條砍伐,記錄留樁上萌條數。分別測定各單株主枝的枝粗、長度和單株、葉片、小枝、主枝鮮質量,測定葉片、小枝和主枝出油率。
1.4" 測定方法
生長指標和生物量的測定:使用游標卡尺(ABS數顯卡尺)、卷尺測量萌條的長度及枝粗,隨后將葉片、小枝和主枝分離,用電子天平測定其鮮質量。
樟樹精油提取:將新鮮黃樟地上部分分為葉片、小枝和主枝后,各稱取一定質量。使用水蒸氣蒸餾法來提取精油,用棕色玻璃瓶進行儲存,對其進行稱量,計算出油率。
出油率=(瓶油重-瓶重)/測定生物量
1.5" 數據統計與分析
數據利用Excel 2020整理、計算和方差分析,出油率先進行數據轉換,再進行方差分析,顯著性水平為p<0.05。
2" 結果與分析
2.1" 兩種化學型黃樟矮林萌芽數及萌條數比較
由表1可知,芳樟醇型黃樟矮林的萌芽數為12.27個,萌條數為4.17個,萌芽數與萌條數均大于檸檬醛型黃樟,呈現顯著差異。芳樟醇型黃樟矮林的萌芽數比檸檬醛型黃樟矮林萌芽數多3.74個,約是檸檬醛型黃樟萌芽數的1.44倍。在萌條數上,兩者僅相差0.09條,差異不顯著。
2.2" 兩種化學型黃樟矮林萌條長度和粗度比較
由表2可知,檸檬醛型與芳樟醇型黃樟矮林的萌條長度、粗度之間存在顯著差異。檸檬醛型黃樟矮林的萌條長度、粗度均大于芳樟醇型黃樟,兩者的萌條長度、粗度分別相差54.66 cm、10.99 mm。檸檬醛型黃樟矮林的萌條長度約是芳樟醇型黃樟的1.49倍,粗度約是芳樟醇型黃樟矮林的1.61倍。
2.3" 兩種化學型黃樟矮林生物量及分配
2.3.1" 兩種化學型黃樟矮林生物量差異
由表3可知,檸檬醛型和芳樟醇型黃樟矮林的單株、單株葉、單株小枝及單株主枝的鮮質量之間存在顯著差異。檸檬醛型黃樟矮林的單株、單株葉、單株小枝及單株主枝的鮮質量均高于芳樟醇型黃樟矮林,兩者分別相差937.46 g、406.17 g、262.29 g和269 g。檸檬醛型黃樟矮林的單株、單株葉、單株小枝及單株主枝鮮質量比芳樟醇型黃樟矮林分別高了55.79%、38.70%、63.57%和123.24%。
2.3.2" 兩種化學型黃樟矮林生物量分配規律
由表4可知,檸檬醛型黃樟矮林的葉、(葉+小枝)質量分數小于芳樟醇型黃樟矮林,分別相差0.069和0.056,存在顯著差異。兩種化學型黃樟矮林的小枝質量分數之間差異不顯著,兩者僅相差0.013。檸檬醛型黃樟矮林的主枝質量分數比芳樟醇型黃樟矮林的主枝質量分數高出0.056,存在顯著差異。
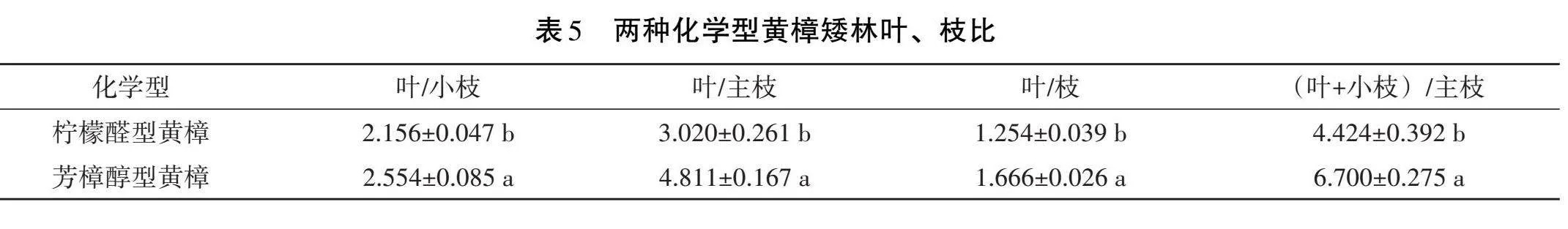
由表5可知,檸檬醛型黃樟矮林的葉與小枝、葉與主枝、葉與枝、(葉+小枝)與主枝的比值均小于芳樟醇型黃樟,分別為0.398、1.791、0.412和2.276,存在顯著差異。芳樟醇型黃樟矮林葉與小枝、葉與主枝、葉與枝、(葉+小枝)與主枝的比值分別是檸檬醛型黃樟矮林的1.12、1.59、1.33和1.51倍。
2.3.3" 兩種化學型黃樟矮林出油率及分配規律
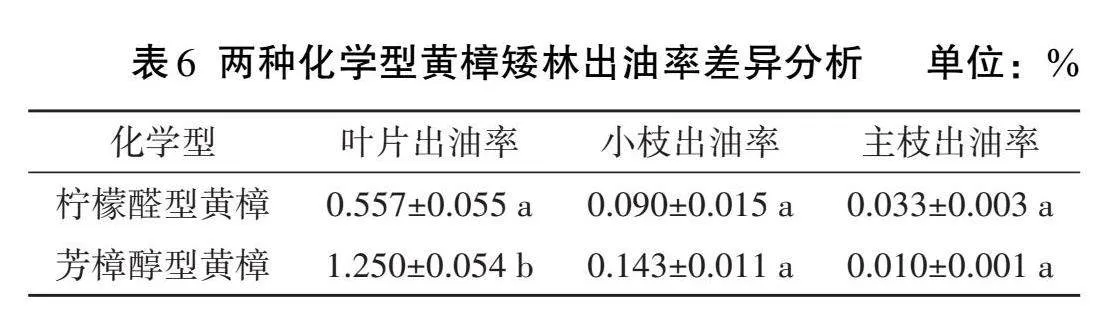
由表6可知,芳樟醇型黃樟矮林的葉片出油率大于檸檬醛型黃樟矮林,兩者相差0.693%,存在顯著差異。檸檬醛型黃樟矮林的小枝出油率略高于檸檬醛型黃樟矮林,兩者相差0.053%,主枝出油率略低于檸檬醛型黃樟矮林,兩者相差0.023%,差異不顯著。檸檬醛型與芳樟醇型黃樟矮林的精油分布均是葉片出油率>小枝含油率>主枝含油率,其精油分配規律相似,葉片出油率顯著高于小枝和主枝。檸檬醛型黃樟矮林的葉片出油率是小枝的6.19倍、主枝的16.88倍,小枝的出油率是主枝的2.73倍;芳樟醇型黃樟矮林的葉片出油率是小枝的8.74倍、主枝的125倍,小枝的出油率是主枝的14.3倍。
3" 討論與結論
3.1" 兩種化學型黃樟矮林萌芽數及萌條數差異性
伐樁萌芽更新是指伐樁上的休眠芽萌發進而生長形成植株[5],萌芽能力直接關系到生物量積累及經濟效益。萌芽能力除了受品種影響外,外界環境、栽植密度、樹齡及伐樁高度等多重因素均會對其產生影響[6-8]。影響矮林萌條生長的因素有基因型、光照、施肥、種植密度等,其中基因型是主導因子。有研究表明,伐樁的萌芽能力與其直徑呈正相關關系,與伐樁的高度則呈負相關關系,伐樁的萌芽植株數量隨著伐樁基徑的增大,初期增長后趨于減少;相反,隨著伐樁高度的提升,萌芽植株數量則呈現增加的趨勢[9-11]。本研究結果表明,檸檬醛型和芳樟醇型黃樟矮林在萌條數量上并未表現出顯著差異。這一現象可能歸因于矮林密植的生長環境,其中萌條之間在生長空間、光照條件及養分資源等方面存在激烈的競爭,最終導致僅有部分萌芽能夠成功發展成為萌條。
3.2" 兩種化學型黃樟矮林生物量積累及分配規律
生物量是衡量植物物質積累及環境資源利用效能的關鍵指標,其大小直接體現了植物在空間資源利用與積累方面的能力[12-13]。本研究中,檸檬醛型黃樟矮林的總生物量、各組分生物量都大于芳樟醇型黃樟矮林,造成這種差異的原因可能是生物量與萌條長度、粗度指標吻合,即萌條長度、粗度大,所得的生物量就更大。由于種植密度較高,兩種化學型黃樟矮林處于相互競爭生長狀態,此時不利于樹冠橫向生長和枝干增粗生長,從而轉向增高生長,究其原因可能由于基因型的不同、種植密度過高導致兩種化學型黃樟矮林的生物量存在差異。植物生物量的分配可以改變苗木生長發育過程中生長、繁殖和防御等功能的選擇次序變化,同時也是影響植物資源獲取、競爭的重要因素[14-15]。本研究結果表明,檸檬醛型黃樟矮林的生物量均大于芳樟醇型黃樟矮林,在兩種化學型黃樟矮林中,葉片生物量占總生物量比例最大,再者是小枝生物量,占比最小的是主枝生物量,(葉+小枝)生物量占總生物量的80%以上。造成芳樟醇型黃樟矮林生物量小,但葉枝比高的原因可能是因為芳樟醇型黃樟矮林需要更多的葉進行光合作用促進自身生長,從而達到增加總生物量的目的,因此導致葉枝比反而高。
3.3" 兩種化學型黃樟矮林出油率及分配規律
精油是樟樹次級代謝的產物,而精油含量則受遺傳、光合作用等因素綜合影響[16-17]。出油率是影響精油產量的絕對指標,出油率與精油產量呈正相關。在不同化學型的黃樟中,其各個部位的精油含量存在顯著差異,這很可能是由于遺傳基因的差異性控制所致,也可能與不同化學型在精油合成過程中,相關酶的種類及活性存在不同有關[18-19]。本研究發現,在兩種化學型的黃樟矮林中,葉片的出油率最高,其次是小枝,而主枝的出油率最低,研究結果和肖祖飛等、周松松等的研究結果相同,均是葉片出油率大于枝條出油率[20-21]。導致葉片出油率最高的因素可能是葉片需要進行光合作用,與生長節律有關。樟樹的精油在植株各部分的含量不同,本研究結果表明,在兩種化學型的黃樟矮林中,精油含量從高到低依次為:葉片含油量、小枝含油量、主枝含油量、葉片精油含量顯著高于小枝和主枝。有研究表明,芳樟精油含量在不同月份、不同季節均有變化,在5月份和7—9月份葉精油含量都處于較高水平[22]。本研究中,檸檬醛型黃樟矮林出油率和芳樟醇型黃樟矮林出油率差異顯著,究其原因可能是基因型的差異造成的。
參考文獻:
[1] 張月婷,劉新亮,秦愛文,等.插穗、基質及生根劑對黃樟組培苗扦插生根影響[J]南方林業科學,2022,50(1):5-8.
[2] 肖珍泉.黃樟在園林綠化中的地位及其栽培技術[J].廣東建材,2008(2):180-181.
[3] 肖祖飛,鐘麗萍,張北紅,等.黃樟的研究進展[J].南方林業科學,2020,48(2):62-65.
[4] 吳航,王建軍,劉馳,等.黃樟化學型的研究[J].植物資源與環境,1992(4):45-49.
[5] 林武星,葉功富,黃金瑞,等.杉木萌芽更新原理及技術述評[J].福建林業科技,1996(2):20-24.
[6] 肖祖飛,李鳳,龍清鑫,等.伐樁高度對樟樹及不同種源萌發特性的影響[J].南昌工程學院學報,2020,39(6):64-68.
[7] 易平,韋鑠星,韋穎文,等.伐樁高度對香樟萌芽性能的影響分析[J].廣西林業科學,2012(3):268-270.
[8] 姜睿,張北紅,李鳳,等.留樁高度對芳樟矮林萌芽特性及得油率的影響[J].安徽林業科技,2021(6):13-18.
[9] 明安剛,陳厚榮,李武志,等.伐樁高度對尾巨桉萌芽更新的影響[J].遼寧林業科技,2011(5):20-21.
[10] 杜春花,楊衛明,陸斌.次生林改造應用草甘膦毒殺幾種喬灌木樹種伐樁的除萌試驗[J].西部林業科學,2008,31(6):82-85.
[11] 李榮,張文輝,何景峰,等.遼東櫟伐樁萌苗的發育規律[J].林業科學,2012(3):82-87.
[12] 李奮燦.密度制約的菊芋生長及產量特性研究[D].蘭州:蘭州大學,2019.
[13] 陳聞,王晶,吳海平,等.5種海島典型園林綠化植物的耐鹽性[J].浙江農林大學學報.2017,34(2):283-293.
[14] 謝然,陶冶,常順利.四種一年生荒漠植物構件形態與生物量間的異速生長關系[J].生態學雜志,2015,34(3):648-655.
[15] 董道瑞,李霞,萬紅梅,等.塔里木河下游胡楊(Populuseuphratica)地上生物量估測[J].中國沙漠,2013,33(3):724-730.
[16] 王以紅,覃子海,吳幼媚,等.芳樟醇型樟樹選優與其無性系的含樟油性狀評價[J].西部林業科學,2010(2):18-21.
[17] 盛盼進,張北紅,另青艷,等.GA_3對芳樟矮林萌芽及生長的影響[J].農業研究與應用,2023(3):43-48.
[18] 汪信東,張月婷,秦政,等.樟樹MVA途徑基因克隆與表達模式初探[J].中南林業科技大學學報,2021,41(5):110-121.
[19] 黃秋良,張國防,謝亞兵,等.無性繁殖方式對芳樟葉精油及主成分的影響[J].福建林業科技,2016(2):156-159.
[20] 肖祖飛,艾卿,金志農,等.芳樟矮林生長節律及精油動態變化研究[J].江西農業大學學報,2021,43(4):834-841.
[21] 周松松,李江,何小三,等.不同化學型黃樟不同部位的精油變化[J].經濟林研究,2022(4):256-266.
[22] 張國防,馮娟,于靜波,等.不同化學型芳樟葉精油及主成分含量的時間變化規律[J].植物資源與環境學報,2012,21(4):82-86.
(責任編輯:敬廷桃)
