惡臭假單胞菌F1表達(dá)P450酶催化檸檬烯生物合成紫蘇醇
2024-02-02 14:59:54底心怡劉春立劉秀霞楊艷坤白仲虎
食品與發(fā)酵工業(yè) 2024年2期
關(guān)鍵詞:產(chǎn)量
底心怡,劉春立*,劉秀霞,楊艷坤,白仲虎*
1(江南大學(xué),糧食發(fā)酵與食品生物制造國(guó)家工程研究中心,江蘇 無(wú)錫,214122) 2(江南大學(xué),工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無(wú)錫,214122)3(江南大學(xué) 生物工程學(xué)院,江蘇 無(wú)錫,214122)
紫蘇醇,也被稱為二氫枯草醇,是一種天然存在的單環(huán)萜類(lèi)化合物,來(lái)自植物的甲羥戊酸途徑。紫蘇醇性能穩(wěn)定,耐熱、耐酸,可溶于甲醇、乙醇、乙腈等有機(jī)溶劑,具有類(lèi)似于芳樟醇、松油醇的特殊氣味,在醫(yī)藥、日化、食品等行業(yè)具有廣闊的應(yīng)用前景[1]。例如,在動(dòng)物實(shí)驗(yàn)中,紫蘇醇能抑制法尼基轉(zhuǎn)移酶和香葉酰轉(zhuǎn)移酶,從而抑制翻譯后腫瘤蛋白的法尼基化、異戊二烯化,從而抑制腫瘤細(xì)胞激活,使其細(xì)胞周期停止在G2/M期[2]。作為一種植物代謝物,紫蘇醇也存在于多種植物精油成分中,如薰衣草、薄荷和檸檬草等。目前獲取紫蘇醇的主要方法分為化學(xué)合成和生物合成。對(duì)于化學(xué)合成,可使用有機(jī)溶劑或超臨界二氧化碳萃取等方法從天然植物精油中獲得紫蘇醇,但產(chǎn)品含量低,且分離不易,成本較高[1,3]。以天然存在的單萜類(lèi)化合物為原料,通過(guò)異構(gòu)化、皂化等過(guò)程也可生成,但選擇性不高,且所使用的氧化劑對(duì)于環(huán)境污染較大[4-5]。對(duì)于生物合成,常使用檸檬烯作為底物,通過(guò)生物酶系統(tǒng)如嗜熱脂肪芽孢桿菌(Bacillusstearothermophilus) BR388中的微生物酶系統(tǒng)進(jìn)行轉(zhuǎn)化[6]。然而,氧化酶體系大多較為復(fù)雜,不具有足夠的區(qū)域特異性,會(huì)產(chǎn)生大量沸點(diǎn)和疏水性相似的副產(chǎn)物,如香芹酮、松油醇等,后續(xù)需要昂貴的純化過(guò)程才能獲得較高純度的紫蘇醇[7]。因此,紫蘇醇生物合成的宿主菌株及生物酶尚有開(kāi)發(fā)和改造空間。
細(xì)胞色素P450 (CYP450),又稱P450酶,是一類(lèi)單加氧酶超家族,以血紅素作為輔因子,能夠?qū)Φ孜镞M(jìn)行區(qū)域選擇性氧化,且往往是立體選擇性氧化[8]。P450酶廣泛存在于動(dòng)物、植物、真菌、原生動(dòng)物、細(xì)菌和古菌中。在哺乳動(dòng)物中,P450酶附著于內(nèi)質(zhì)網(wǎng),用于氧化類(lèi)固醇、脂肪酸等物質(zhì),并在激素合成和分解過(guò)程中發(fā)揮著重要作用。在植物中,P450酶用于合成激素、脂肪酸等對(duì)植物本身具有保護(hù)作用的化合物。在微生物中,P450酶參與各種生化反應(yīng),包括初級(jí)、次級(jí)代謝物合成及反硝化作用[9]。目前已有較為成熟的應(yīng)用P450酶的研究成果。SCHNEIDER等[10]利用來(lái)自石油假單孢菌(Pseudomonasoleovorans)的輔酶A非依賴型脂肪酸吸收系統(tǒng),以表達(dá)P450 BM3的重組大腸桿菌(Escherichiacoli)為宿主催化制備12-,13-,14-羥基化十五酸,能夠有效替代傳統(tǒng)生物轉(zhuǎn)化長(zhǎng)鏈脂肪酸為亞末端羥基化脂肪酸的方法。BEZALEL等[11]利用P450酶的單加氧特性,使用糙皮側(cè)耳菌(Pleurotusostreatus)降解菲類(lèi)化合物。GIANG等[12]通過(guò)序列同源性比對(duì)發(fā)現(xiàn)球狀紅球菌(Rhodococcusgloberulus)來(lái)源的P450酶(CYP108 N12)具有能夠氧化檸檬烯、傘花烴等物質(zhì)的功能。分岐桿菌(Mycobacteriumsp.)HXN-1500來(lái)源的P450酶,即CYP153A6,可氧化C6-C11正烷烴,也可以結(jié)構(gòu)較為復(fù)雜的烴類(lèi)作底物,能得到高度區(qū)域選擇性烴化的產(chǎn)物[8,13]。當(dāng)?shù)孜餅闄幟氏r(shí),CYP153A6可將其特異性氧化為紫蘇醇,目前已在多種宿主中得到應(yīng)用(圖1)。GUDIMINCHI等[14]使用pCom8-PFR1500載體在大腸桿菌中表達(dá)CYP153A6、鐵氧還蛋白和鐵氧還蛋白還原酶,在自誘導(dǎo)培養(yǎng)基生長(zhǎng)的大腸桿菌所得紫蘇醇產(chǎn)量最高達(dá)到1.85 mol/L。VAN等[15]在惡臭假單胞菌GPo12中表達(dá)CYP153A6生物轉(zhuǎn)化檸檬烯生成紫蘇醇,但惡臭假單胞菌GPo12的檸檬烯耐受度較低,紫蘇醇產(chǎn)量也因此受到限制。

圖1 P450酶生物轉(zhuǎn)化檸檬烯生成紫蘇醇示意圖Fig.1 Biosynthesis of perillyl alcohol catalyzed by cytochrome P450
惡臭假單胞菌(Pseudomonasputida)是一種腐生營(yíng)養(yǎng)土壤桿菌,其營(yíng)養(yǎng)背景豐富,廣泛存在于土壤、水體、動(dòng)植物體表和各種富含蛋白質(zhì)的物質(zhì)中[16]。惡臭假單胞菌酶背景復(fù)雜,代謝途徑繁多,能利用多種碳源,且對(duì)pH、溶劑等極端環(huán)境條件表現(xiàn)出高度穩(wěn)健性和耐受性,因此多用于環(huán)境污染治理及有機(jī)廢物再循環(huán)利用,并因在土壤中的定植能力和降解各種化學(xué)物質(zhì)的能力而聞名[17-19]。作為革蘭氏陰性菌,惡臭假單胞菌同樣具有培養(yǎng)成本低、生長(zhǎng)周期短、安全無(wú)害等特點(diǎn)。其中,惡臭假單胞菌F1與模式菌株的KT2440不同,并不常作為外源基因表達(dá)或目的產(chǎn)物合成的宿主菌株[20-21]。惡臭假單胞菌F1含有豐富的氧化酶系,如tod、cym、cmt酶系等,使其具有對(duì)環(huán)境中芳香族、萜類(lèi)物質(zhì)較高耐受性,且cym酶系中的CymAa、CymAb具有選擇性羥化活性,作為生產(chǎn)此類(lèi)物質(zhì)的宿主菌株具有較大潛力[22]。
本研究將構(gòu)建穿梭質(zhì)粒表達(dá)載體,并以大腸桿菌BL21 (DE3)、惡臭假單胞菌F1作為宿主菌株,表達(dá)選擇性羥化酶CymAa、CymAb和P450酶、鐵氧還蛋白、鐵氧還蛋白還原酶,以檸檬烯為底物生物合成紫蘇醇,展現(xiàn)惡臭假單胞菌F1作為萜類(lèi)物質(zhì)合成載體菌株的優(yōu)勢(shì)。
1 材料與方法
1.1 材料與試劑
1.1.1 主要試劑
2×TaqPCR Mastermix、2×PrimeStar,Takara公司;2×Multif Seamless Assembly Mix,Abclonal公司;引物,蘇州金唯智生物科技有限公司。
1.1.2 菌株與質(zhì)粒
本研究中使用的菌株與質(zhì)粒如表1所示,所使用引物如表2所示。

表1 本研究中使用的菌株與質(zhì)粒Table 1 Strains and plasmids used in this study

表2 本研究中使用的引物Table 2 Primers used in this study
1.1.3 培養(yǎng)基
LB培養(yǎng)基(g/L):酵母提取物5,蛋白胨5,NaCl 10,主要用于質(zhì)粒構(gòu)建菌株及發(fā)酵種子液培養(yǎng)。LB固體培養(yǎng)基為上述液體培養(yǎng)基中添加1%(質(zhì)量分?jǐn)?shù))的瓊脂粉。
M9培養(yǎng)基(1 L):0.1 mL 1%維生素B12,0.01 mL 10% 維生素B1,0.1 mL trace礦物鹽溶液(108 g/L FeCl3·6H2O, 2 g/L H3BO3, 4 g/L ZnCl2·7H2O, 5 g/L MnSO4·H2O, 7 g/L CoCl2·6H2O, 7 g/L Na2MoO4·H2O, 8 g/L CuSO4·6H2O),0.1 mL 1 mol/L CaCl2, 2 mL 1 mol/L MgSO4, 50 mL 20%D-葡萄糖,50 mL 10% 酪蛋白水解物, 100 mL 10×M9鹽溶液 (60 g/L Na2HPO4·7H2O, 30 g/L KH2PO4, 5 g/L NaCl),主要用于產(chǎn)物發(fā)酵菌株培養(yǎng)。
抗生素:卡那霉素終質(zhì)量濃度為50 mg/L。
1.1.4 儀器與設(shè)備
ETC821 PCR基因擴(kuò)增儀,東勝公司;411BR8608 MicroPulser電轉(zhuǎn)儀,BIO-RAD;759S21023 UV-Vis Spectrophotometer紫外可見(jiàn)分光光度計(jì),上海棱光公司;GCMS-QP2020 SYSTEM氣相色譜質(zhì)譜聯(lián)用儀,島津公司。
1.1.5 標(biāo)準(zhǔn)品
檸檬烯標(biāo)準(zhǔn)品:(-)-檸檬烯,(-)-Limonene,別名(S)-(-)-二戊烯,CAS編號(hào)5989-54-8,分子式C10H16,分子量136.24,規(guī)格及純度為>95.0%(GC),沸點(diǎn)為64 ℃/15 mmHg(lit.),密度0.845,購(gòu)于Aladdin公司。
紫蘇醇標(biāo)準(zhǔn)品:紫蘇醇,Perillyl alcohol,CAS編號(hào)536-59-4,分子式C10H16O,分子量152.23,規(guī)格及純度為90%,室溫貯存,購(gòu)于Aladdin公司。
乙酸乙酯:Ethyl acetate,CAS編號(hào)141-78-6,分子式C4H8O2,分子量88.11,規(guī)格及純度為≥99.5%,沸點(diǎn)為77 ℃,相對(duì)密度0.902,購(gòu)于國(guó)藥滬試公司。
1.2 實(shí)驗(yàn)方法
1.2.1 基因克隆和質(zhì)粒構(gòu)建
使用引物pBBR1-vector-F/pBBR1-vector-R,以廣宿主載體pBBR1MCS2為模板,擴(kuò)增含有卡那霉素抗性基因、pBBR1復(fù)制子的DNA片段;使用引物promoter-F/promoter-R,以質(zhì)粒載體pTrc99a為模板,擴(kuò)增含有l(wèi)acI基因、trc啟動(dòng)子、lacO序列以及rrnB T1終止子的DNA片段。使用Gibson組裝法連接上述兩片段,構(gòu)成質(zhì)粒pBBR1-Ptrc。
從惡臭假單胞菌數(shù)據(jù)庫(kù)(https://www.pseudomonas.com/)獲得cymAa(Pput_2903)、cymAb(Pput_2902)基因的核苷酸序列信息。從NCBI(https://www.ncbi.nlm.nih.gov/)獲得P450酶、鐵氧還蛋白、鐵氧還蛋白還原酶基因(GenBank:AJ783967.1)的核苷酸序列信息,并對(duì)其編碼序列進(jìn)行密碼子優(yōu)化(參考大腸桿菌BL21),由蘇州金唯智生物科技有限公司合成質(zhì)粒pET-Cyp450-Fdr-Fdx。使用引物pBBR1cymAabv-F/pBBR1cymAabv-R和pBBR1P450v-F/pBBR1P450v-R,以pBBR1-Ptrc為模板擴(kuò)增線性化載體片段;使用引物cymAab-F/cymAab-R,以惡臭假單胞菌F1基因組為模板擴(kuò)增含有cymAa和cymAb基因的DNA片段;使用引物P450-F/P450-R,以實(shí)驗(yàn)室保存質(zhì)粒pET-Cyp450-Fdr-Fdx為模板擴(kuò)增含有P450酶、鐵氧還蛋白、鐵氧還蛋白還原酶基因的DNA片段。分別使用Gibson組裝法連接載體和片段,構(gòu)成質(zhì)粒pBBR1-cymAab和pBBR1-P450-Fdr-Fdx。
1.2.2 惡臭假單胞菌F1電轉(zhuǎn)化感受態(tài)制備
將保存于-80 ℃的惡臭假單胞菌F1接種于裝有5 mL LB培養(yǎng)基的50 mL離心管,于30 ℃過(guò)夜培養(yǎng),轉(zhuǎn)接至100 mL LB培養(yǎng)基培養(yǎng)至OD600=0.8。將培養(yǎng)液分裝至2個(gè)50 mL離心管于5 000×g,4 ℃離心10 min。將獲取的細(xì)胞分別用30 mL預(yù)冷的無(wú)菌水、3 mmol/L 4-羥乙基哌嗪乙磺酸溶液{2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid,HEPES}重懸并再次離心,最后加入2 mL預(yù)冷的10%甘油重懸并每80 μL分裝于500 μL離心管備用。
1.2.3 菌株培養(yǎng)
將質(zhì)粒轉(zhuǎn)化至大腸桿菌BL21 (DE3)與惡臭假單胞菌F1中。PCR驗(yàn)證為陽(yáng)性后于平板上隨機(jī)挑取3個(gè)菌落,接種至裝有5 mL LB培養(yǎng)基的50 mL離心管中,于220 r/min恒溫培養(yǎng)箱培養(yǎng),其中大腸桿菌BL21 (DE3)生長(zhǎng)溫度為37 ℃,惡臭假單胞菌F1生長(zhǎng)溫度為30 ℃。16 h后轉(zhuǎn)接至裝有5 mL M9培養(yǎng)基的20 mL搖瓶中,當(dāng)OD600值至0.6~0.8時(shí)添加相應(yīng)濃度的檸檬烯作為底物、以及1 mmol/L 異丙基β-D-1-硫代吡喃半乳糖苷(isopropyl-beta-D-thiogalactopyranoside,IPTG)誘導(dǎo)表達(dá),于30 ℃培養(yǎng)24 h和48 h 取樣進(jìn)行檢測(cè)。
1.2.4 紫蘇醇檢測(cè)
取1 mL發(fā)酵液、0.5 mL乙酸乙酯與過(guò)量無(wú)水Na2SO4加入2 mL離心管,渦旋振蕩5 min后于13 000 r/min離心10 min,用針管取最上層有機(jī)相過(guò)濾膜后注入氣相小瓶待檢。GC-MS儀器設(shè)置:電子碰撞(electron impact, EI)檢測(cè)器和DB-5MS色譜柱(30 m×0.32 mm×0.25 μm)。方法條件:入口溫度為260 ℃,氮?dú)廨d氣恒定流量為44 cm/s,離子源溫度為250 ℃,掃描30~500m/z。執(zhí)行以下程序:初始溫度為80 ℃保持1 min,然后以10 ℃/min的速率升至260 ℃,最后保持10 min。分流比5.0。通過(guò)與已知濃度的紫蘇醇標(biāo)品繪制的標(biāo)準(zhǔn)曲線比較,將峰面積轉(zhuǎn)換為紫蘇醇濃度進(jìn)行定量。
2 結(jié)果與分析
2.1 廣宿主載體pBBR1-Ptrc質(zhì)粒構(gòu)建
惡臭假單胞菌F1具有酶背景豐富、環(huán)境耐受度高、培養(yǎng)成本低、生長(zhǎng)周期短等優(yōu)點(diǎn),但并不常作為外源基因表達(dá)的宿主菌株,因而缺少可用的基因表達(dá)載體。為使承載目的基因的表達(dá)載體可以正常在大腸桿菌BL21 (DE3)和惡臭假單胞菌F1中擴(kuò)增表達(dá),本研究利用廣宿主載體pBBR1MCS2作為模板擴(kuò)增其復(fù)制子pBBR1,通過(guò)Gibson組裝連接來(lái)自質(zhì)粒pTrc99a的trc啟動(dòng)子和lacI基因,構(gòu)建pBBR1-Ptrc質(zhì)粒。質(zhì)粒元件構(gòu)成如圖2所示。該質(zhì)粒載體能夠在大腸桿菌、惡臭假單胞菌等多種革蘭氏陰性菌中正常擴(kuò)增表達(dá),具有可以連接外源基因片段的多克隆位點(diǎn),并可以通過(guò)控制IPTG的添加調(diào)控其連接的目的基因的表達(dá)。

圖2 質(zhì)粒構(gòu)建示意圖Fig.2 Construction of plasmids.
2.2 CymAab和P450酶表達(dá)載體構(gòu)建及驗(yàn)證
獲得可用的表達(dá)載體后,需要表達(dá)目的蛋白CymAab和P450酶。CymAab為惡臭假單胞菌F1來(lái)源,分為CymAa和CymAb兩部分,以純化的惡臭假單胞菌F1基因組作為模板,通過(guò)引物擴(kuò)增與Gibson組裝構(gòu)建成質(zhì)粒pBBR1-cymAab。本研究中所使用的P450酶為Mycobacteriumsp.HXN-1500 來(lái)源,分為主體P450酶、輔因子鐵氧還蛋白、鐵氧還蛋白還原酶3個(gè)部分,其序列經(jīng)過(guò)大腸桿菌密碼子偏好優(yōu)化后通過(guò)公司合成為質(zhì)粒pET-Cyp450-Fdr-Fdx,并通過(guò)引物擴(kuò)增與Gibson組裝構(gòu)建成質(zhì)粒pBBR1-Cyp450-Fdr-Fdx。質(zhì)粒元件構(gòu)成與P450酶生物合成紫蘇醇示意圖如圖2所示。
將含有目的基因的質(zhì)粒轉(zhuǎn)化進(jìn)大腸桿菌BL21 (DE3)與惡臭假單胞菌F1中,并以含有空質(zhì)粒pBBR1-Ptrc的菌株EcC和PpC作為空白對(duì)照,添加10 mmol/L檸檬烯作為底物,檢測(cè)誘導(dǎo)后24 h和48 h后發(fā)酵液中的細(xì)胞密度和紫蘇醇濃度。由OD600值測(cè)定結(jié)果可以看出,添加了10 mmol/L檸檬烯后,大腸桿菌BL21 (DE3)的生長(zhǎng)狀態(tài)受到很大程度抑制,而惡臭假單胞菌F1幾乎沒(méi)有受到影響(圖3-a)。對(duì)于紫蘇醇的產(chǎn)量檢測(cè),表達(dá)CymAab酶的實(shí)驗(yàn)組EccymAab、PpcymAab均沒(méi)有檢測(cè)到紫蘇醇生成;而表達(dá)P450酶的實(shí)驗(yàn)組中,由于前述大腸桿菌BL21 (DE3)的生長(zhǎng)受到抑制,只檢測(cè)到較低濃度的紫蘇醇產(chǎn)量,而惡臭假單胞菌F1組即菌株P(guān)pP450在24 h 時(shí)達(dá)到最高紫蘇醇積累,且產(chǎn)量最高組為96.43 mg/L(圖3-b)。從單位OD600的紫蘇醇產(chǎn)量結(jié)果也可看出,惡臭假單胞菌F1發(fā)酵24 h的產(chǎn)量為15.32 mg/L,明顯高于大腸桿菌BL21發(fā)酵48 h的產(chǎn)量7.64 mg/L(圖3-c)。
2.3 不同底物濃度與紫蘇醇產(chǎn)量關(guān)系驗(yàn)證
由于前序?qū)嶒?yàn)中菌株EccymAab、PpcymAab沒(méi)有檢測(cè)到紫蘇醇產(chǎn)物,且大腸桿菌BL21 (DE3)的生長(zhǎng)狀態(tài)受到較大影響,因此選擇P450酶進(jìn)行后續(xù)實(shí)驗(yàn),并調(diào)整了底物檸檬烯的添加濃度,設(shè)置1 mmol/L、5 mmol/L和10 mmol/L的濃度梯度范圍,使用大腸桿菌BL21 (DE3)與惡臭假單胞菌F1作為發(fā)酵宿主,設(shè)置了不添加檸檬烯的空質(zhì)粒組pBBR1-Ptrc作為對(duì)照,檢測(cè)誘導(dǎo)24、48 h后發(fā)酵液中的細(xì)胞密度和紫蘇醇產(chǎn)量。在菌體生長(zhǎng)方面,加入檸檬烯后大腸桿菌BL21 (DE3)的生長(zhǎng)狀態(tài)依然受到了較大的抑制,并且抑制程度隨著添加濃度的升高而增強(qiáng);與之相對(duì),添加檸檬烯的惡臭假單胞菌F1實(shí)驗(yàn)組生長(zhǎng)情況隨著檸檬烯濃度升高實(shí)驗(yàn)組的生長(zhǎng)情況保持穩(wěn)定(圖4-a)。在紫蘇醇產(chǎn)量方面,大腸桿菌BL21 (DE3)由于生長(zhǎng)受到限制,在添加1 mmol/L檸檬烯的情況下經(jīng)過(guò)48 h發(fā)酵所得紫蘇醇最高濃度為28.8 mg/L;而對(duì)于惡臭假單胞菌F1,各組添加了不同濃度檸檬烯后所獲得的紫蘇醇產(chǎn)量也沒(méi)有明顯差異,菌株P(guān)pP450在添加1 mmol/L檸檬烯的情況下經(jīng)過(guò)48 h發(fā)酵所得紫蘇醇最高質(zhì)量濃度為75.6 mg/L,對(duì)于檸檬烯的轉(zhuǎn)化率達(dá)到49.66%(圖4-b)。添加5 mmol/L檸檬烯的大腸桿菌BL21雖然在24 h獲得較高紫蘇醇產(chǎn)量,但在48 h時(shí)產(chǎn)量驟然減少;與之相比惡臭假單胞菌F1各組的紫蘇醇生產(chǎn)積累較為穩(wěn)定,添加1 mmol/L檸檬烯時(shí)在發(fā)酵48 h后達(dá)到最高單位OD600紫蘇醇產(chǎn)量38.49 mg/L(圖4-c)。
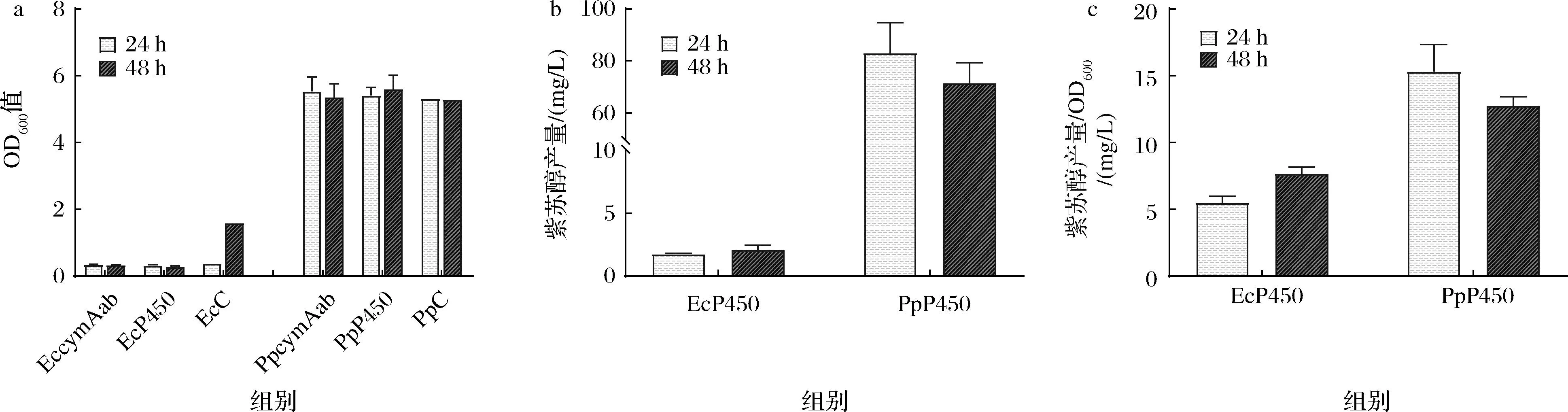
a-OD600值結(jié)果;b-紫蘇醇產(chǎn)量結(jié)果;c-紫蘇醇產(chǎn)量/OD600結(jié)果圖3 添加10 mmol/L檸檬烯大腸桿菌BL21 (DE3)和惡臭假單胞菌F1的OD600和紫蘇醇產(chǎn)量結(jié)果Fig.3 OD600 and Perillyl alcohol yield results of E.coli BL21 (DE3) and P.putida F1 with the addition of 10 mmol/L limonene

a-OD600值結(jié)果;b-紫蘇醇產(chǎn)量結(jié)果;c-紫蘇醇產(chǎn)量/OD600結(jié)果圖4 添加不同濃度檸檬烯大腸桿菌BL21 (DE3)和惡臭假單胞菌F1的OD600和紫蘇醇產(chǎn)量結(jié)果Fig.4 OD600 and perillyl alcohol yield results of E.coli BL21 (DE3) and P.putida F1 with the addition of different concentration of limonene
3 討論
紫蘇醇作為單萜類(lèi)化合物,在醫(yī)藥、食品、日化等方面具有廣泛的應(yīng)用價(jià)值,然而化學(xué)合成法的高昂成本與環(huán)境污染、生物合成法的低選擇性與對(duì)宿主毒性都是獲取紫蘇醇的過(guò)程中亟需解決的問(wèn)題。P450酶的引入實(shí)現(xiàn)了將檸檬烯特異性轉(zhuǎn)化為紫蘇醇,極大程度推動(dòng)了微生物合成紫蘇醇的研究進(jìn)程。但是由于萜類(lèi)物質(zhì)對(duì)微生物具有毒性,更高的底物、產(chǎn)物濃度意味著對(duì)宿主菌株生長(zhǎng)的更大壓力。
大腸桿菌作為常用的工業(yè)模式菌株,其明確的代謝背景、易于培養(yǎng)等特點(diǎn),使得大腸桿菌作為表達(dá)外源基因宿主受到廣泛應(yīng)用。而如實(shí)驗(yàn)結(jié)果顯示,大腸桿菌BL21 (DE3)對(duì)于檸檬烯的耐受度較低,在外源添加檸檬烯的環(huán)境中大腸桿菌BL21 (DE3)的生長(zhǎng)會(huì)受到較大影響,從而影響其蛋白表達(dá)、進(jìn)而使得生物轉(zhuǎn)化能力受到限制。而無(wú)論是否添加檸檬烯,惡臭假單胞菌F1的生長(zhǎng)狀態(tài)均沒(méi)有受到明顯影響,對(duì)于10 mmol/L以內(nèi)的檸檬烯具有較高耐受性。
在羥化酶的選擇上,本研究使用了惡臭假單胞菌F1自身來(lái)源的CymAab和有生物轉(zhuǎn)化檸檬烯生成紫蘇醇相關(guān)報(bào)道的P450酶,結(jié)果證明CymAab并沒(méi)有原先猜測(cè)的能選擇性羥化(S)-(-)-檸檬烯的能力,而P450酶在惡臭假單胞菌F1中效果良好。在進(jìn)一步進(jìn)行惡臭假單胞菌F1表達(dá)P450酶催化紫蘇醇合成實(shí)驗(yàn)探究時(shí),發(fā)現(xiàn)不同檸檬烯添加濃度所獲得的紫蘇醇產(chǎn)量并無(wú)較大差別,因此可以推斷在添加1 mmol/L檸檬烯時(shí)底物濃度對(duì)于P450酶的催化效率已達(dá)到飽和,即一定范圍內(nèi)更高濃度的檸檬烯添加雖然不會(huì)影響惡臭假單胞菌F1的生長(zhǎng)狀態(tài),但也不能被利用生成更多產(chǎn)物。如果在現(xiàn)有的體系基礎(chǔ)上進(jìn)一步進(jìn)行提升紫蘇醇的產(chǎn)量的后續(xù)實(shí)驗(yàn),可以從P450酶表達(dá)量、活性等方面著手。
結(jié)合菌體生長(zhǎng)與紫蘇醇產(chǎn)量二者的數(shù)據(jù),在以檸檬烯為底物生物合成紫蘇醇的過(guò)程中,惡臭假單胞菌F1相較于大腸桿菌BL21 (DE3)或許更適合于作為發(fā)酵宿主。與VAN等[15]在使用惡臭假單胞菌GPo12時(shí)所遇到的情況不同,惡臭假單胞菌F1在含有10 mmol/L檸檬烯濃度以內(nèi)的環(huán)境下展現(xiàn)出較高的耐受性,或許可以成為突破萜類(lèi)物質(zhì)毒性影響宿主生長(zhǎng)問(wèn)題的關(guān)鍵。另一方面,梯度濃度添加的底物的紫蘇醇產(chǎn)物結(jié)果也為后續(xù)實(shí)驗(yàn)縮小了底物使用范圍,并將限制產(chǎn)量的因素引導(dǎo)向P450酶方向。如先前描述表達(dá)載體構(gòu)建時(shí),本實(shí)驗(yàn)所使用的P450酶經(jīng)過(guò)大腸桿菌密碼子偏好優(yōu)化,且惡臭假單胞菌F1為野生型菌株,后續(xù)針對(duì)產(chǎn)量提高的相關(guān)改造具有較大潛力。
4 結(jié)論
本研究通過(guò)構(gòu)建廣宿主表達(dá)載體,并在其上連接P450酶及其輔因子3個(gè)基因后,將質(zhì)粒轉(zhuǎn)入大腸桿菌BL21 (DE3)和惡臭假單胞菌F1中,外源添加檸檬烯并進(jìn)行濃度調(diào)整后,獲得對(duì)檸檬烯具有較高耐受性的惡臭假單胞菌F1,添加1 mmol/L檸檬烯底物發(fā)酵48 h后獲得最高產(chǎn)量75.6 mg/L,轉(zhuǎn)化率達(dá)到49.66%,縮小了生物合成紫蘇醇的有效底物添加范圍,并提供了具有改造潛力的宿主菌株。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當(dāng)代水產(chǎn)(2021年10期)2021-12-05 16:31:48
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
今日農(nóng)業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國(guó)化肥信息(2019年2期)2019-01-18 15:24:35
中國(guó)化肥信息(2019年1期)2019-01-17 21:31:12
中國(guó)化肥信息(2019年4期)2019-01-17 18:47:06
