漂白粉消毒后對蝦養(yǎng)殖源水細菌群落的響應特征研究*
2024-02-24 08:45:16王庚申董鵬生謝建軍許文軍張德民張化俊
海洋與湖沼 2024年1期
王庚申 顏 懿 李 彤 董鵬生 謝建軍許文軍 張德民 張化俊 ①
(1. 寧波大學 農產品質量安全危害因子與風險防控國家重點實驗室 浙江寧波 315211; 2. 寧波大學海洋學院 浙江寧波 315211; 3. 浙江省海洋水產研究所 浙江省海水增養(yǎng)殖重點實驗室 浙江舟山 316021)
為了防控凡納濱對蝦(Litopenaeusvannamei)的傳染性疾病, 對源水消毒是養(yǎng)殖的關鍵環(huán)節(jié)之一。漂白粉是對蝦養(yǎng)殖源水消毒中常用的一種鹵素類消毒劑, 具有價格低、殺菌力強、使用方便等特點。漂白粉是氫氧化鈣、氯化鈣、次氯酸鈣的混合物, 溶解在水中時水解成次氯酸離子和氯離子。次氯酸離子的強氧化作用可以殺死細菌, 氯離子也表現(xiàn)出一定的殺菌功能(Maetal, 2019)。研究表明, 大多數(shù)病原對次氯酸鈣敏感, 且殺菌效果與劑量有關(Parketal, 2004;Limsuwanetal, 2008; Chaetal, 2012)。一般認為, 對養(yǎng)殖源水消毒可以有效去除病原微生物, 但由于消毒機制的非特異性, 水體中的有益微生物也會被殺滅。由于細菌類群生理及群落組成的多樣性, 水體中不同的微生物類群會對消毒表現(xiàn)出不同的響應或恢復能力(Becerra-Castroetal, 2016)。目前, 對消毒后養(yǎng)殖源水細菌群落的恢復動態(tài)仍知之甚少。
抗生素抗性基因(antibiotic resistant genes, ARGs)可以通過水平轉移將移動遺傳元件轉至病原菌中,使病原菌產生抗生素抗性, 威脅公共衛(wèi)生健康(Joet al, 2021)。由于河流匯入及水產養(yǎng)殖活動, ARGs 在近海海區(qū)環(huán)境中普遍存在(Hedbergetal, 2018), 近海養(yǎng)殖源水是對蝦養(yǎng)殖池塘環(huán)境中ARGs 的主要來源(夏濤濤等, 2022), 在華南及華東地區(qū)的對蝦養(yǎng)殖源水中均檢測到了多種養(yǎng)殖中常見抗生素的ARGs (Wangetal, 2019a; Yuanetal, 2019)。近幾年, 城市污水、飲用水處理領域的研究顯示, 氯消毒可以有效降低水中的ARGs 豐度(Yoonetal, 2017; Wangetal, 2020), 水產養(yǎng)殖用水消毒的研究也證實了這一點(趙曉雨等,2021)。但氯消毒不能完全去除水體中的ARGs (Yuanetal, 2015), 即使攜帶ARGs 的細菌被殺滅, ARGs 仍可以釋放到水環(huán)境中通過水平轉移進行傳播(姜瀚集等, 2019)。然而, 關于養(yǎng)殖源水消毒后ARGs 豐度的動態(tài)變化規(guī)律尚未見報道。
養(yǎng)殖水體細菌群落是對蝦養(yǎng)殖系統(tǒng)的重要組成,對維持水質穩(wěn)定及對蝦健康具有重要意義(杜世聰?shù)?2019)。一般認為, 源水進入養(yǎng)殖系統(tǒng)后除改善水體理化因子外, 也會影響水體微生物群落組成(Heyseetal,2021)。有研究報道氯消毒會引起水體細菌類群的異常增殖, 當消毒后病原菌和耐抗生素細菌增殖時, 可能導致潛在的微生物健康風險(Linetal, 2016; Dengetal, 2019; Wangetal, 2021a)。因此, 了解漂白粉消毒后養(yǎng)殖源水細菌群落、病原菌及抗生素抗性基因的恢復動態(tài), 尤其是不同消毒濃度的影響, 對優(yōu)化對蝦養(yǎng)殖系統(tǒng)微生物管控策略具有重要的指導作用。基于此, 本研究利用不同濃度漂白粉消毒養(yǎng)殖源水, 通過高通量測序比較消毒前后養(yǎng)殖源水的理化因子、細菌群落結構及病原菌豐度變化, 同時利用RT-qPCR 檢測養(yǎng)殖源水ARGs 的豐度變化, 探究源水細菌群落對不同濃度漂白粉消毒后的響應特征, 以期為對蝦養(yǎng)殖生產中漂白粉的使用提供理論基礎。
1 材料與方法
1.1 實驗材料與設計
本實驗在浙江省海洋水產研究所試驗場進行,實驗用對蝦養(yǎng)殖源水取自周邊海域, 溫度27.3~ 27.9 °C,鹽度25.5~25.7, pH 8.25~8.21。漂白粉購自湖北省應城市利必康化工有限公司, 有效氯含量28%~35%。
實驗在室內300 L 塑料水槽中進行, 共設置2 個高、低漂白粉消毒濃度: 60、20 mg/L, 每組4 個重復。消毒前(DC)、消毒后余氯衰減至0.1 mg/L 時0~3 d(D0、D1、D2、D3)定時取2 L 水樣, 1 L 用于水質理化指標測定; 1 L 用0.22 μm 濾膜過濾后保存于-80 °C,用于檢測微生物和抗性基因。
采用HANNA HI-97734 余氯和總氯便攜光度計(DPD 法, 量程0.00~10.00 mg/L, 精度為±0.03 mg/L)測定水體中的余氯濃度; 用YSI Pro2030 測定水體溶解氧, YSI Pro10 測定水體pH。水體理化因子化學需氧量(COD)采用堿性高錳酸鉀法, 氨氮采用靛酚藍分光光度比色法, 亞硝酸鹽氮(NO2-N)采用萘乙二胺分光光度法, 硝酸鹽氮(NO3-N)采用鋅-鉻還原法, 活性磷酸鹽(AP)用磷鉬藍分光光度法測定。
1.2 DNA 提取、高通量測序
使用PowerSoil ?Pro Kit (Qiagen)試劑盒提取水樣DNA, 通過0.8%瓊脂糖凝膠電泳檢測提取DNA 的質量, 用紫外分光光度計檢測DNA 的濃度和純度。用帶Barcode 的338F (5′-ACTCCTACGGGAGGCAGC AG-3′)和806R (5′-GGACTA CHVGGGTWTCTAAT-3′)引物對細菌16S rRNA 的V3~V4 區(qū)進行PCR 擴增。PCR 擴增條件: 98 °C 預變性30 s; 98 °C 變性15 s,50 °C 退火30 s, 72 °C 延伸30 s, 27 個循環(huán); 72 °C 延伸5 min。反應體系(25 μL): 5 μL 5×High GC Buffer、5 μL 5×Reaction Buffer、2 μL 10 mmol/L dNTPs、2 μL引物(10 μmol/L )、0.25 μL FastPfu 聚合酶; 2 μL 模板DNA, 滅菌雙蒸水補足25 μL。PCR 產物經純化、定量后, 使用TruSeq Nano DNA LT Library Prep Kit 建庫, 構建的文庫經質檢和定量后, 使用 Illumina NovaSeq 平臺PE250 進行雙端測序。
1.3 測序數(shù)據(jù)處理
使用QIIME2 2021.2 版本對原始數(shù)據(jù)進行質量控制, 去除接頭, 利用DADA2 插件去噪, 并生成擴增子序列變體(ASV)表。以每個ASV 中豐度最高的作為代表序列, 在SILVA138 數(shù)據(jù)庫中比對確定其所屬類群。去除葉綠體(Chloroplast)、線粒體(Mitochondria)及未被鑒定的ASVs, 共獲得了2933 個ASVs。
1.4 水環(huán)境細菌病原數(shù)據(jù)庫分析
將實驗樣品中每個ASV 的代表序列在水環(huán)境細菌病原數(shù)據(jù)庫(DPiWE, dayuz.com) (董鵬生等,2021)進行BLAST 比對、注釋, 參數(shù)閾值E-value<1×10-6, 序列同源性>98.5%, 最后輸出病原菌比對信息, 共比對獲得病原菌169 個ASV。由于數(shù)據(jù)庫注釋信息的差異, 所得病原菌序列以病原菌庫的注釋信息為準。
1.5 抗性基因檢測
采用Real-time qPCR 方法對常見的8 種ARGs(表1)進行定量分析。qPCR 在羅氏LightCycler480 上使用AceQ qPCR SYBR Green Master Mix (Vazyme)進行, 反應體系: 10 μL 2×SYBR real-time PCR premixture、0.8 引物(10 μmol/L)、1 μL 模板DNA, 滅菌雙蒸水補足20 μL。PCR 反應條件: 95 °C 5 min;95 °C 15 s, 55~62 °C 30 s, 72 °C 30 s, 40 個循環(huán), 之后進行熔解曲線分析。通過Ct 值和標準曲線計算基因拷貝數(shù), 以每毫升養(yǎng)殖源水的拷貝數(shù)來測定ARGs 的濃度。

表1 抗生素抗性基因及RT-PCR 引物Tab.1 Antibiotic resistance genes and RT-PCR primers
1.6 統(tǒng)計分析
通過Bray-Curtis 距離進行非度量多維尺度分析(NMDS)分析, 展示漂白粉消毒后水體細菌群落的演替規(guī)律。基于Bray-Curtis 距離的相似性分析(ANOSIM)分析各組細菌群落的相似性。通過R 中的“pheatmap”軟件包繪制熱圖。使用“psych”軟件包計算類群相關性矩陣, 保留皮爾森相關性系數(shù)|r|>0.6, 且P<0.05 的數(shù)據(jù)矩陣, 使用Gephi(v0.9.2)構建微生物群落的共現(xiàn)網(wǎng)絡, 并進行加權隨機模塊化網(wǎng)絡分析。利用SPSS 20.0 在用單因素方差分析(one-way ANOVA)分析樣品間ARGs 豐度的差異(P<0.05)。
2 結果
2.1 消毒前后對蝦養(yǎng)殖源水水體理化因子變化
實驗期間, 對蝦養(yǎng)殖源水溫度在27.5~32.9 °C,鹽度在漂白粉消毒后小幅升高, 高濃度組始終高于低濃度組; 溶解氧、化學需氧量濃度在消毒后(D0)迅速下降并保持穩(wěn)定; pH、硝酸鹽濃度在消毒后(D0)升高隨后保持穩(wěn)定; 亞硝酸鹽濃度在消毒后(D0)升高,之后大幅震蕩, 氨氮濃度則在消毒后緩慢上升; 活性磷酸鹽濃度在消毒后出現(xiàn)分化, 低濃度消毒組始終高于高濃度組(圖1)。


圖1 消毒前后對蝦養(yǎng)殖源水水質理化因子變化Fig.1 Changes of physicochemical factors of shrimp source water before and after bleaching powder disinfection
2.2 消毒前后對蝦養(yǎng)殖源水的細菌群落組成
消毒后, 高、低濃度組細菌類群門水平組成均發(fā)生了顯著的變化, 擬桿菌門(Bacteroidota)的相對豐度較消毒前均顯著升高(P<0.05), 高濃度組變形菌門(Proteobacteria)的相對豐度則顯著下降(P<0.05) (圖2a)。消毒前(DC), 高濃度組養(yǎng)殖源水的主要門類包括變形菌門(53.71%)、擬桿菌門(27.15%)、放線菌門(Actinobacteriota, 9.32%)、Patescibacteria (1.84%)及藍細菌門(Cyanobacteria, 1.46%)等; 消毒后余氯衰減至0.1 mg/L 時(D0), 優(yōu)勢菌門為變形菌門(25.54%)、擬桿菌門(73.91%)。而低濃度組在消毒前(DC)的主要門類為變形菌門(74.74%)、擬桿菌門(12.76%)、藍菌門(7.34%)、放線菌門(2.76%)及Patescibacteria (1.55%)等; 消毒后(D0), 優(yōu)勢菌門為變形菌門(69.26%)、擬桿菌門(29.19%)。消毒后第1~2 天, 高濃度組的優(yōu)勢菌門以變形菌門(79.90%、51.17%)、擬桿菌門(19.83%、44.85%)為主, 而低濃度組則以變形菌門為優(yōu)勢菌, 相對豐度達到了98.78%、95.78%。至第3天, 高濃度組的優(yōu)勢菌門均以變形菌門(91.58%)、蛭弧菌門(Bdellovibrionota) (5.17%)、擬桿菌門(2.19%)為主, 低濃度組的優(yōu)勢菌門為變形菌門(87.92%)、蛭弧菌門(6.34%)、擬桿菌門(2.79%)、Patescibacteria(1.67%)。

圖2 消毒前后對蝦養(yǎng)殖源水細菌群落相對豐度圖Fig.2 Relative abundance of bacterial community in shrimp source water before and after bleach disinfection
從屬水平看(圖2b), 消毒前高低濃度組養(yǎng)殖源水的優(yōu)勢菌屬包括Thalassococcus、Nautella、unclassified Stappiaceae、Candidatus_Aquiluna、Erythrobacter、Synechococcus_CC9902、Hymenobacter、Phaeodactylibacter,以上菌屬的總相對豐度在高、低濃度組分別占比27.80%、29.61%。消毒后第0~2 天, 高濃度組中薄層菌屬(Hymenobacter, 19.77%~73.88%) 和Nautella(5.26%~55.98%)交替占據(jù)優(yōu)勢, 第3 天, 薄層菌屬豐度迅速下降。而低濃度組中薄層菌屬(29.06%)僅在第0 天占據(jù)優(yōu)勢,Nautella(37.40%~78.34%)則在第1~2天維持優(yōu)勢。消毒后第2 天, 高、低濃度組的小紅卵菌屬(Rhodovulum)豐度開始升高, 第3 天在兩組中均占據(jù)優(yōu)勢(24.08% 、 14.52%), 其次是Nautella(22.00%、12.12%)。
在ASV 水平上(圖3), 源水細菌群落中相對豐度最高的50 個ASVs 主要歸類于紅桿菌科Rhodobacteraceae(16 個)、交替單胞菌科Alteromonadaceae (4 個)、黃桿菌科Flavobacteriaceae (3 個)和Nitrincolaceae (3 個),其中21 個ASVs 在消毒后相對豐度下降, 22 個ASVs相對豐度升高。消毒后, ASV848 (Nautella)在高、低濃度組的相對豐度均較消毒前顯著上升, 歸類于薄層菌屬的ASV1561、ASV953 的相對豐度在第1 天顯著升高, 之后開始下降; 紅桿菌科的 ASV1480、ASV1249、ASV2053 僅在高濃度組顯著下降,ASV1764 (小紅卵菌屬)則在高濃度組中顯著上升。

圖3 消毒前后對蝦養(yǎng)殖源水中的優(yōu)勢ASVs 熱圖(相對豐度前50 個ASVs)Fig.3 Dominant ASVs of shrimp source water before and after bleaching powder disinfection (top 50 ASVs in relative abundance)
2.3 消毒前后細菌群落結構演替特征
基于Bray-Curtis 距離的NMDS 分析發(fā)現(xiàn)消毒后水體細菌群落與消毒前分區(qū)明顯, 而高、低濃度組未完全分離(圖4); ANOSIM 分析也發(fā)現(xiàn), 消毒前、后細菌群落差異顯著(P<0.05), 而消毒后高、低濃度之間差異不顯著(P>0.05)。
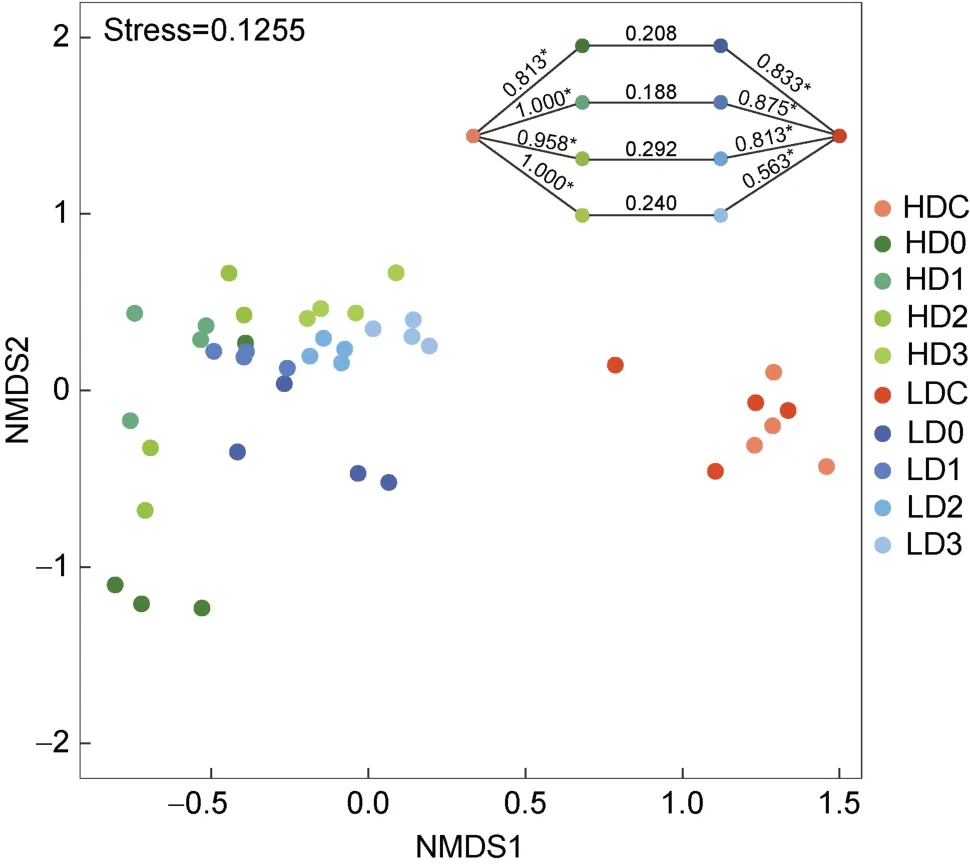
圖4 基于Bray-Curtis 距離的NMDS 分析水體細菌群落結構Fig.4 NMDS analysis of bacterial community structure in source water based on the Bray-Curtis distance
2.4 消毒前后水體細菌群落的共現(xiàn)模式
消毒前及高、低濃度組消毒后的水體細菌群落的共現(xiàn)網(wǎng)絡包含425、238、348 個節(jié)點, 分別由1 776、1 137、1 958 個邊連接(圖5a~5c, 表2), 說明消毒后節(jié)點減少, 低濃度消毒會增加群落間的相互作用。相比消毒前, 消毒后水體細菌群落共現(xiàn)網(wǎng)絡的模塊化系數(shù)、平均聚類系數(shù)降低, 而其他網(wǎng)絡拓撲學參數(shù)則升高; 消毒后, 除了網(wǎng)絡直徑外, 高濃度組共現(xiàn)網(wǎng)絡的平均聚類系數(shù)、平均路徑長度、平均度均低于低濃度組。高、低濃度組的網(wǎng)絡分別由6 個主要模塊構成,分別占整個網(wǎng)絡的69.74% (9.24%~13.87%)、79.38%(8.33%~18.39%), 而對照組模塊較為平均, 模塊Ⅰ到模塊Ⅵ僅占整個網(wǎng)絡的26.6% (4%~4.71%)。
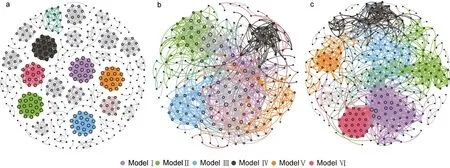
圖5 消毒前后水體微生物群落共現(xiàn)網(wǎng)絡Fig.5 Co-occurrence network of bacterial communities
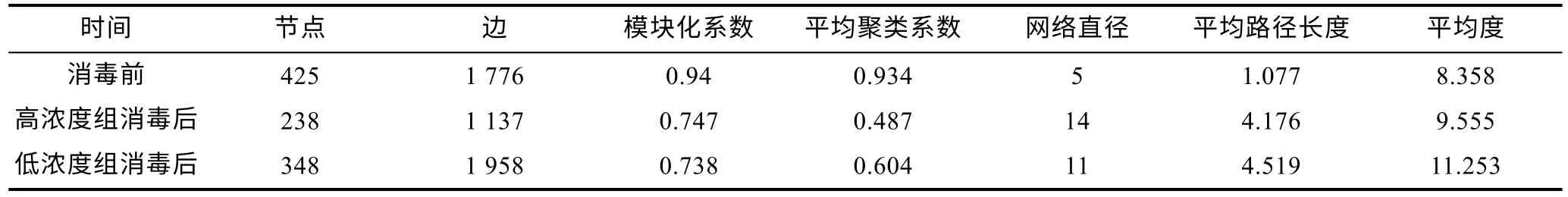
表2 共現(xiàn)網(wǎng)絡的拓撲學參數(shù)Tab.2 Topological parameters of co-occurrence networks of water bacterial communities before and after bleaching powder disinfection
2.5 消毒前后水體病原菌的組成
使用DPiWE 數(shù)據(jù)庫對消毒前后的水體細菌高通量測序數(shù)據(jù)進行序列比對和病原菌物種注釋, 消毒前水體病原菌有23 屬, 總序列數(shù)為6 461; 消毒后高濃度組病原菌有37 屬, 總序列數(shù)為2 912, 低濃度組有31 屬, 總序列數(shù)為6 584, 說明消毒后病原菌種類增加, 而高濃度組總序列數(shù)顯著下降。
對蝦養(yǎng)殖源水的優(yōu)勢病原菌(平均相對豐度大于0.01%)豐度占總病原菌豐度的95.59%, 其中50%為魚類病原, 50%為跨宿主共患病病原。消毒前, 優(yōu)勢病原菌為假埃希氏菌(Pseudescherichia, 1.56%)、微球菌屬(Micrococcus, 0.53%)、Thalassobius(0.23%)等; 消毒后, 高濃度組的優(yōu)勢病原菌Thalassobius(0.32%)、不動桿菌屬(Acinetobacter, 0.10%), 低濃度組則為Thalassobius(0.88%) 、 假 交 替 單 胞 菌 屬(Pseudoalteromonas, 0.13%)等。水中優(yōu)勢病原菌在消毒后具有一定時間上的差異, 而Thalassobius在高、低濃度組均逐漸占據(jù)絕對優(yōu)勢。值得一提的是, 弧菌屬(Vibro)病原菌的相對豐度在消毒后明顯下降, 但在源水中仍存在(圖6)。

圖6 消毒前后病原菌的相對豐度堆積圖Fig.6 Relative abundance of pathogenic bacteria before and after bleaching powder disinfection
2.6 消毒前后抗性基因的動態(tài)變化
本研究中, 4 大類8 個ARGs 在消毒前的養(yǎng)殖源水中均有檢出(圖7),tetA 是豐度最高的ARG, 達到107copies/mL,cfr是豐度最低的ARG (102copies/mL),其他ARGs 的豐度在103~104copies/mL 水平。消毒后, 余氯衰減至0.1 mg/L 時(D0), 高、低濃度組sul1、floR、cfr、tetQ 的豐度較消毒前(DC)顯著下降(P<0.05),而tetA、strB、cmlA、sul2 的豐度變化趨勢在高、低濃度組出現(xiàn)分化。sul1、floR 的豐度均在隨后的1~2 d內恢復到消毒前的數(shù)量級水平, 而cfr、tetQ 基因在實驗期間未能恢復到消毒前的數(shù)量級。

圖7 消毒前后8 種ARGs 的動態(tài)變化Fig.7 Changes of abundances of 8 ARGs before and after bleaching powder disinfection
3 討論
含氯消毒劑在海水養(yǎng)殖疾病預防中的應用已被養(yǎng)殖業(yè)者廣泛接受, 但由于水體中不同細菌對消毒劑的敏感性存在差異, 消毒后水體的細菌群落結構的恢復動態(tài)值得關注。研究表明, 氯消毒會改變飲用水的細菌群落組成, 降低群落結構多樣性(Holingeret al, 2014; Daietal, 2020)。Becerra-Castro 等(2016)研究發(fā)現(xiàn)紫外線、臭氧及光催化臭氧化等多種方式消毒城市污水和地表水, 3 d 后的兩種類型水體的變形菌門豐度明顯上升。Duan 等(2020)報道漂白粉消毒對蝦育苗用水5 d 后, 變形菌門和擬桿菌門豐度上升, 放線菌門豐度下降。本研究中, 漂白粉消毒后養(yǎng)殖源水的細菌群落與消毒前差異顯著, 但高低濃度組之間無顯著差異。消毒后(D0), 高濃度組變形菌門豐度明顯下降, 擬桿菌門占據(jù)主導地位, 而低濃度組變形菌門豐度無明顯變化, 說明高濃度漂白粉對變形菌門的殺滅效果更強, 存在劑量依賴效應。消毒后第3 天,高、低濃度組細菌群落組成趨于一致, 變形菌門豐度占據(jù)優(yōu)勢, 表明消毒后變形菌門類群可快速恢復。共現(xiàn)網(wǎng)絡的復雜性代表了不同樣本微生物群落之間的根本差異(Shietal, 2016), 網(wǎng)絡的拓撲學參數(shù)可以表征微生物之間的相互作用(Barberánetal, 2012)。漂白粉消毒后, 細菌網(wǎng)絡節(jié)點數(shù)量減少, 模塊化程度降低,說明消毒降低了細菌的共現(xiàn)網(wǎng)絡復雜程度。低濃度組共現(xiàn)網(wǎng)絡節(jié)點之間的連接最為復雜, 并且網(wǎng)絡中心性最高, 表明消毒后低濃度的水體細菌群落穩(wěn)定性高于高濃度組。
從屬水平看, 消毒后(D0)養(yǎng)殖源水優(yōu)勢屬的數(shù)量減少, 但總相對豐度增加, 與Wan 等(2022)的研究結果一致。屬水平的變化趨勢與ASV 水平相似, 主要由單個或幾個優(yōu)勢ASV 的豐度變化主導。消毒后0~2 d, 薄層菌屬和Nautella在占據(jù)主導地位, 高濃度組薄層菌屬豐度高于低濃度組; 第3 天, 小紅卵菌屬在高低濃度組均占據(jù)優(yōu)勢, 其次是Nautella豐度, 且高濃度豐度均高于低濃度組。有研究認為, 薄層菌屬具有輻射抗性, 能適應多種極端環(huán)境(Collinsetal,2000)。Nautella常見于海水養(yǎng)殖環(huán)境及對蝦腸道中(李秋芬等, 2011; Sakamietal, 2014; Zhengetal,2016), 但尚未發(fā)現(xiàn)對凡納濱對蝦的致病性。小紅卵菌屬是一類光養(yǎng)細菌, 分泌多種殺菌活性物質, 對水產動物有益生作用(Looetal, 2013; Changetal, 2019;Kogaetal, 2021)。
DPiWE 是一個基于高通量測序數(shù)據(jù)分析水體環(huán)境病原菌群落特征和溯源的綜合數(shù)據(jù)庫, 提供了人類介水傳染病、水產動物疾病及跨宿主共患病等病害的細菌病原的分類學和宿主信息(董鵬生等, 2021)。通過DPiWE 數(shù)據(jù)庫比對, 養(yǎng)殖源水中優(yōu)勢病原菌為埃希氏菌、微球菌屬及Thalassobius等, 與象山大黃魚網(wǎng)箱養(yǎng)殖區(qū)水體中優(yōu)勢病原菌(弧菌屬、黏著桿菌屬Tenacibaculum、埃希氏-志賀氏菌屬Escherichia-Shigella和不動桿菌屬)不同, 且總體豐度偏低(Houet al, 2021), 說明本實驗源水環(huán)境受到水產養(yǎng)殖活動影響的程度低。漂白粉消毒后病原菌總豐度降低, 但種類增加, 與病原菌對氯消毒的抗性有關(Songetal,2019; Wangetal, 2019b), 如消毒后新增鏈球菌Streptococcus、梭狀芽孢桿菌Clostridium、埃希氏-志賀氏菌等病原菌被普遍報道具有氯消毒抗性(Wangetal, 2021b)。高濃度組病原菌在消毒后初期豐度下降,而低濃度組在D0 時高于消毒前, 說明高濃度漂白粉對病原菌有更好地殺滅作用。源水病原菌組成與豐度存在明顯的動態(tài)演替, 實驗后期均以Thalassobius占主導地位, 有研究認為Thalassobius中包含多種魚類病原菌, 但未見對蝦類致病的報道(Kanehisa, 2018)。
本研究中, 養(yǎng)殖源水中豐度最高的ARG 是tetA,其次是cmlA 與floR, 與文獻報道對蝦養(yǎng)殖近海水源水主要為磺胺類ARGs (sul2)不同, 養(yǎng)殖源水中ARGs豐度與該水產養(yǎng)殖區(qū)常用的抗生素類型有關(夏濤濤等, 2022), 同時受人類活動、生活污水排入、降雨等多種因素影響(Shaoetal, 2018; Liuetal, 2020)。有研究表明, 氯消毒能有效去除水體中的ARGs (Yoonet al, 2017), 去除效率與有效氯濃度有關(Huangetal,2013)。本實驗中, 漂白粉消毒后(D0)僅對sul1、floR、cfr、tetQ 有一定的去除效果, 且在有效氯衰減后短時間內恢復至消毒前水平, 說明漂白粉消毒養(yǎng)殖源水不能有效去除ARGs (Tangetal, 2023)。值得注意的是, 消毒后高濃度組sul1、sul2、tetA、tetQ 等ARGs的豐度高于低濃度組, 分析原因可能與消毒后攜帶相關抗性基因的細菌快速再生有關(Sullivanetal,2017)。氯消毒對水體ARGs 的去除具有選擇性, 對引用水的研究發(fā)現(xiàn), 氯消毒除對sul1 有明顯的去除作用外, 對ampC、aphA2、blatem1、tetA、tetG、ermA、ermB 等ARGs 均有富集作用(Shietal, 2013)。Wan 等(2022)也報道氯消毒30 min 對多種ARGs 的豐度沒有影響。本實驗也有類似結果,tetA、cmlA 和strB 的豐度在消毒后無明顯變化, 但具體機制有待進一步研究。
4 結論
研究發(fā)現(xiàn), 漂白粉消毒改變了源水的細菌群落結構及共現(xiàn)網(wǎng)絡的復雜性。消毒后, 兩個濃度組源水的擬桿菌門豐度上升, 而60 mg/L 漂白粉組變形菌門的豐度顯著降低; 屬水平上, 薄層菌屬和Nautella在消毒后源水中占據(jù)優(yōu)勢; 至第3 天, 兩組的細菌群落組成在門和屬水平趨于一致。60 mg/L 漂白粉對病原菌豐度和恢復速度的抑制作用優(yōu)于20 mg/L。漂白粉消毒對養(yǎng)殖源水中sul1、floR、cfr、tetQ 有一定的去除作用, 但對其他 ARGs 幾乎無影響, 顯示了對ARGs 去除的選擇性, 且高低濃度組之間無差異。綜合分析認為, 相比20 mg/L 組, 60 mg/L 漂白粉消毒可以顯著降低變形菌門的豐度, 抑制病原菌快速增殖,可以作為凡納濱對蝦養(yǎng)殖中更好地消毒選擇。
