張掖市山丹縣馬鈴薯枯萎病菌的鑒定
2024-02-26 09:11:20何樹(shù)文王彥崔凌霄許永鋒王澤浩楊成德
中國(guó)蔬菜 2024年1期
何樹(shù)文,王彥,崔凌霄,許永鋒,王澤浩*,楊成德*
(1 張掖市植保植檢站,甘肅張掖 734000;2 甘肅農(nóng)業(yè)大學(xué)植物保護(hù)學(xué)院,甘肅蘭州 730070)
馬鈴薯具有較高的經(jīng)濟(jì)價(jià)值和營(yíng)養(yǎng)價(jià)值(齊恩芳 等,2023),是我國(guó)主要的糧食作物之一,年種植面積超過(guò)600 萬(wàn)hm2(9 000 萬(wàn)畝),產(chǎn)業(yè)發(fā)展?jié)摿薮螅ɡ顕?guó)景 等,2023)。隨著馬鈴薯種植面積的日益增長(zhǎng),馬鈴薯病害問(wèn)題也日益加劇,其病害種類(lèi)不斷增多,發(fā)病程度逐年加重(羅愛(ài)花 等,2019)。其中,由鐮刀菌所引起的馬鈴薯枯萎病(Fusariumavenaceum)已成為馬鈴薯最重要的病害之一,其分布廣泛,在美國(guó)(Taylor & Blodgett,1937)、印度(Azil et al.,2021)、意大利等馬鈴薯種植區(qū)均有發(fā)生。在中國(guó),馬鈴薯枯萎病主要發(fā)生在內(nèi)蒙古、甘肅、河北和新疆等馬鈴薯主產(chǎn)區(qū)(安小敏 等,2017)。據(jù)報(bào)道,馬鈴薯枯萎病在我國(guó)各地的發(fā)病率可達(dá)15%~45%,易于流行且難以防治,嚴(yán)重時(shí)會(huì)直接導(dǎo)致植株整株死亡,嚴(yán)重制約我國(guó)馬鈴薯產(chǎn)業(yè)的健康發(fā)展(陳春艷 等,2014)。
甘肅省是我國(guó)馬鈴薯的主要產(chǎn)區(qū)之一,據(jù)王玉琴等(2014)報(bào)道,引起甘肅省武威市天祝縣馬鈴薯枯萎病的病原菌為燕麥鐮刀菌(F.avenaceum),但其他地區(qū)病原菌種類(lèi)尚不清楚。因此,本試驗(yàn)對(duì)甘肅省山丹縣馬鈴薯枯萎病病原菌進(jìn)行分離和致病性測(cè)定,利用顯微觀察其形態(tài)學(xué)特征,并結(jié)合分子生物學(xué)分析對(duì)病原菌進(jìn)行鑒定,以期對(duì)山丹縣馬鈴薯枯萎病的田間診斷和綜合防治提供依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
供試標(biāo)本為2020 年9 月于張掖市山丹縣采集的具有典型枯萎病癥狀的馬鈴薯病株。供試培養(yǎng)基為馬鈴薯葡萄糖瓊脂培養(yǎng)基(PDA):馬鈴薯200 g、葡萄糖15~20 g、瓊脂16~18 g、蒸餾水1 000 mL。
1.2 試驗(yàn)方法
1.2.1 病原菌分離及致病性測(cè)定 采用組織分離法對(duì)采集到的馬鈴薯病株進(jìn)行病原菌分離。在超凈工作臺(tái)中,用無(wú)菌手術(shù)刀切取0.5 cm 長(zhǎng)的馬鈴薯病株根部組織,并于0.1%升汞中消毒1 min,無(wú)菌水沖洗3 次,用無(wú)菌濾紙吸干組織表面水分后置于PDA 培養(yǎng)基,25 ℃培養(yǎng)5~7 d,單孢純化后編號(hào)并于4 ℃條件下保存?zhèn)溆谩?/p>
將單孢分離后的分離物置于PDB 液體培養(yǎng)基,25 ℃、180 r · min-1培養(yǎng)5 d,采用灌根法將其接種于生長(zhǎng)30 d 的健康馬鈴薯根莖處,以接種無(wú)菌水為對(duì)照,重復(fù)3 次,保濕48 h,連續(xù)觀察馬鈴薯植株發(fā)病情況。待植株發(fā)病后再次進(jìn)行組織分離,將分離物的菌落形態(tài)和孢子形態(tài)等與接種菌株進(jìn)行比較。
1.2.2 形態(tài)學(xué)鑒定 于25 ℃條件下培養(yǎng)5~7 d 后觀察菌落的形態(tài)、顏色等,測(cè)量直徑,在顯微鏡(10 × 40)下觀察菌絲、分生孢子、分生孢子梗和厚垣孢子等特征,并進(jìn)行拍照。
1.2.3 分子生物學(xué)鑒定 收集25 ℃條件下培養(yǎng)5~7 d 的病原菌菌絲,利用OMEGA 真菌DNA 提取試劑盒(D3195-01,美國(guó)Omega 生物技術(shù)公司),按說(shuō)明書(shū)步驟提取病原菌DNA。
選取通用引物ITS1(5′-TCCGTAGGTGA ACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATT GATATGC-3′)對(duì)病原菌DNA 進(jìn)行擴(kuò)增,方法參考相關(guān)文獻(xiàn)并經(jīng)優(yōu)化后應(yīng)用于本試驗(yàn)。PCR 擴(kuò)增體系(32 μL)為:Master Mix 10 μL,DNA 模板1 μL,ITS1 0.5 μL,ITS4 0.5 μL,ddH2O 20 μL;擴(kuò)增程序?yàn)椋?5 ℃預(yù)變性5 min;95 ℃變性30 s,54 ℃退火30 s,72 ℃延伸1 min,共30 個(gè)循環(huán);72 ℃延伸7 min,4 ℃保存。
利用鐮刀菌屬特異性引物EF728M(5′-CAT CGAGAAGTTCGAGAAGG-3′) 和Tef1R(5′-GCCATCCTTGGAGATACCAGC-3′)對(duì)病原菌DNA 進(jìn)行PCR 擴(kuò)增。反應(yīng)體系(25 μL)為:模板DNA 4 μL,10 μmol · L-1上、下游引物各1 μL,2× Easy Taq PCR Super Mix 12.5 μL,ddH2O 6.5 μL。PCR 擴(kuò)增程序?yàn)椋?5 ℃預(yù)變性5 min;94 ℃變性45 s,60 ℃退火45 s,72℃ 延伸2 min,35 個(gè)循環(huán);72 ℃延伸10 min。
取6 μL 擴(kuò)增產(chǎn)物,于1.2%瓊脂糖凝膠上進(jìn)行電泳檢測(cè),將具特異性條帶的擴(kuò)增產(chǎn)物送至西安擎科生物工程有限公司進(jìn)行測(cè)序。所得序列于NCBI中進(jìn)行Blast 比對(duì),用MEGA 7.0 軟件采用鄰接法(Neighbor-joining)構(gòu)建系統(tǒng)發(fā)育樹(shù)(崔凌霄 等,2018)。
2 結(jié)果與分析
2.1 田間發(fā)病癥狀
患病植株下層葉片最先出現(xiàn)萎蔫,主要表現(xiàn)為葉片邊緣卷曲、呈褐色,隨后上部幼嫩葉片出現(xiàn)萎蔫等癥狀(圖1-a),切開(kāi)病株維管束呈現(xiàn)褐色(圖1-b)。

圖1 馬鈴薯枯萎病癥狀
2.2 病原菌分離及致病性測(cè)定
經(jīng)組織分離,于張掖市山丹縣馬鈴薯枯萎病病株上共分離純化得到3 株真菌分離物,編號(hào)分別為1-1、3-2 和3-6。
將3 株分離物進(jìn)行盆栽致病性測(cè)定。結(jié)果表明,接種50 d 后,接種分離物1-1 的馬鈴薯植株發(fā)病嚴(yán)重,葉片萎蔫、卷曲,呈褐色,病株維管束切開(kāi)后呈褐色,發(fā)病較嚴(yán)重時(shí)出現(xiàn)整株枯死等癥狀(圖2-a);接種分離物3-2 和3-6 的馬鈴薯植株均出現(xiàn)明顯的枯萎癥狀,但發(fā)病癥狀較分離物1-1 引起的癥狀輕,主要表現(xiàn)為植株下部老葉枯萎、凋落(圖2-b、c);接種無(wú)菌水的對(duì)照均無(wú)發(fā)病癥狀(圖2-d)。將發(fā)病后的馬鈴薯再次進(jìn)行組織分離,均得到與接種物培養(yǎng)性狀相同的分離物,依據(jù)科赫氏法則,確定分離物1-1、3-2 和3-6 均為引起張掖市山丹縣馬鈴薯枯萎病的致病菌。

圖2 病原菌致病性測(cè)定
2.3 病原菌鑒定
2.3.1 形態(tài)學(xué)鑒定 將致病菌株1-1、3-2 和3-6接種于PDA 平板,25 ℃培養(yǎng)7 d。其中,菌株1-1生長(zhǎng)較慢,菌落表面中心由白色漸變?yōu)榧t色,邊緣呈白色不規(guī)則狀(圖3-a),未觀察到分生孢子;菌株3-2 菌絲密集,菌落為白色(圖3-d),大型分生孢子呈鐮刀狀,3~5 個(gè)分隔,其中一端彎曲程度大于另一端,大小為(50~75)μm ×(4~5)μm,小型分生孢子橢圓形(圖3-e);菌株3-6 菌絲紅色,生長(zhǎng)后期菌落中央表面呈黃色(圖3-g),菌絲有隔,厚垣孢子圓形(圖3-i),未觀察到分生孢子。
2.3.2 基因序列分析鑒定 提取3 株病原菌的基因組DNA,利用ITS 通用引物擴(kuò)增并測(cè)序,通過(guò)Blast 相似性分析后,選擇同源性大于95%的序列進(jìn)行系統(tǒng)發(fā)育樹(shù)構(gòu)建。結(jié)果表明,分離自張掖市山丹縣馬鈴薯的3 株病原菌中,菌株3-2 與木賊鐮刀菌(F.equiseti)聚于同一分支,菌株1-1 和3-6 與三線鐮刀菌(F.tricinctum)聚于同一分支(圖4)。利用EF-1α 特異性引物對(duì)3-2 和1-1 進(jìn)行擴(kuò)增并測(cè)序,下載同源性較高的序列構(gòu)建系統(tǒng)發(fā)育樹(shù),其結(jié)果與ITS 結(jié)果相一致(圖5)。結(jié)合形態(tài)特征,最終將3-2 鑒定為木賊鐮刀菌,1-1 和3-6 鑒定為三線鐮刀菌,其中木賊鐮刀菌引起的馬鈴薯枯萎病為國(guó)內(nèi)首次報(bào)道,三線鐮刀菌引起的馬鈴薯枯萎病為甘肅省首次報(bào)道。

圖4 基于ITS 基因序列構(gòu)建病原菌1-1、3-2 和3-6 及其他相關(guān)菌株的系統(tǒng)發(fā)育樹(shù)
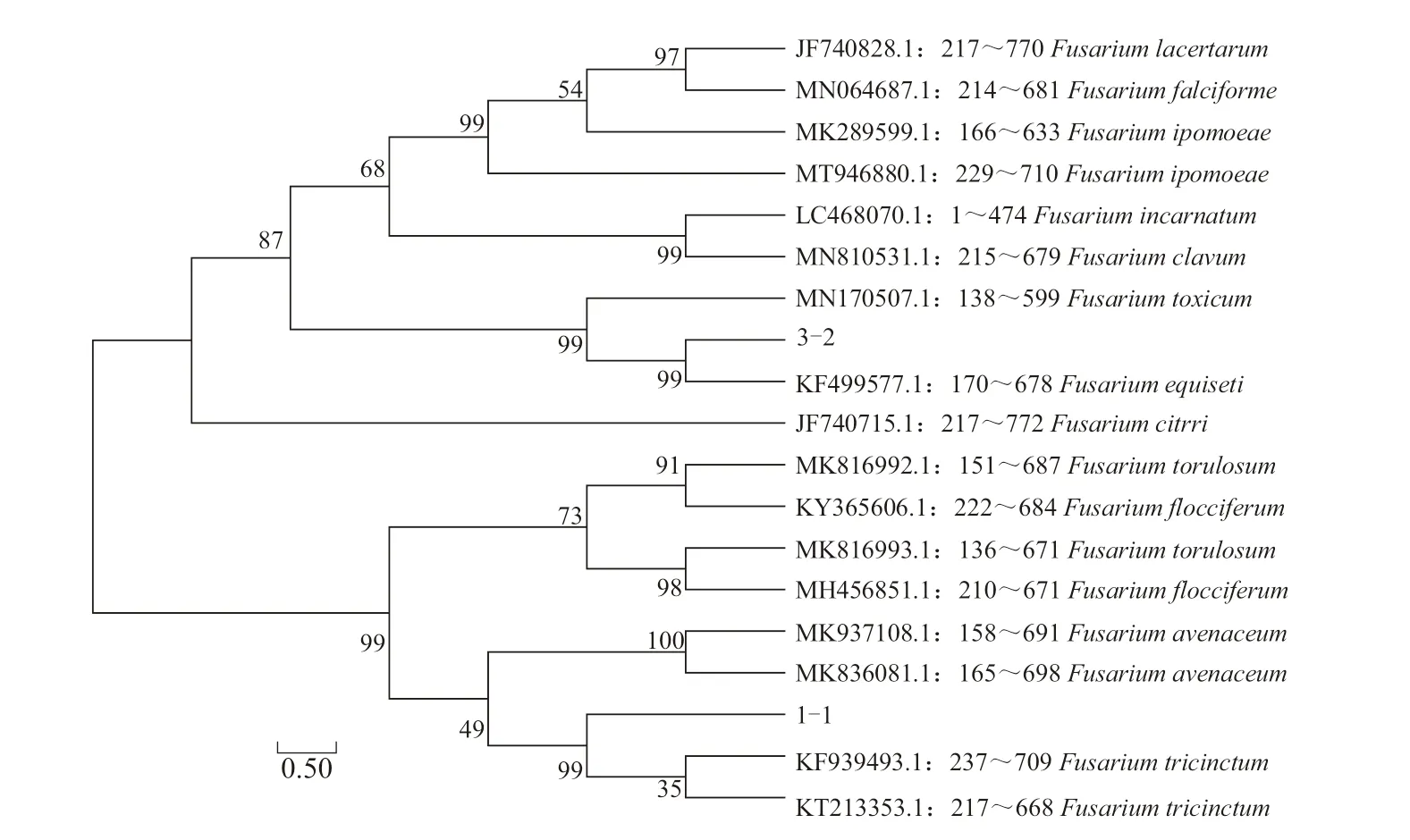
圖5 基于EF-1α 基因序列構(gòu)建病原菌1-1 和3-2 及其他相關(guān)菌株的系統(tǒng)發(fā)育樹(shù)
3 討論與結(jié)論
據(jù)國(guó)內(nèi)外報(bào)道,能夠引起馬鈴薯枯萎病的鐮刀菌共有8 種,分別為尖孢鐮刀菌(F.oxysporum)、茄病鐮刀菌(F.solani)、 接骨木鐮刀菌(F.sambucinum)、雪腐鐮刀菌(F.nivale)、串珠鐮刀菌(F.moniliforme)、三線鐮刀菌(F.tricinctum)、銳頂鐮刀菌(F.acuminatum)和燕麥鐮刀菌(F.avenaceum)(曲延軍,2014)。不同區(qū)域分離的菌株在致病力方面存在差異性,如賈瑞芳等(2023)報(bào)道的139 株鐮刀菌屬真菌均能引起馬鈴薯枯萎病,但不同種的鐮刀菌屬菌株在馬鈴薯上存在一定的致病力分化。鐮刀菌對(duì)環(huán)境的敏感性較強(qiáng),且遺傳變異較大,其同一屬內(nèi)不同菌株間可能存在著遺傳多樣性,進(jìn)而造成致病力存在差異性。其中,引起甘肅省武威市天祝縣馬鈴薯枯萎病的病原菌為燕麥鐮刀菌(F.avenaceum)(王玉琴 等,2014),其他地區(qū)病原菌種類(lèi)尚不明確。在本試驗(yàn)中,對(duì)甘肅省山丹縣馬鈴薯枯萎病病原菌進(jìn)行分離和致病性測(cè)定,共得到3 株具有致病性的鐮刀菌,包含1株木賊鐮刀菌(3-2)和2 株三線鐮刀菌(1-1 和3-6)。早期木賊鐮刀菌主要侵染馬鈴薯薯塊,能夠引起馬鈴薯薯塊的干腐病。2009—2010 年,對(duì)密歇根州馬鈴薯貯藏庫(kù)內(nèi)的薯塊進(jìn)行調(diào)查后發(fā)現(xiàn),在370 個(gè)發(fā)病薯塊中共分離得到228 株鐮刀菌,其中木賊鐮刀菌為第二大優(yōu)勢(shì)種,占分離總株數(shù)的19.3%(Gachango et al.,2012)。同樣,在?demi?和Torbal? 倉(cāng)庫(kù)中,從468 個(gè)發(fā)病薯塊中分離得到了41 株具有致病性的鐮刀菌,其中木賊鐮刀菌占比14.63%(Yikilmazsoy & Tosun,2021)。除此之外,木賊鐮刀菌也可造成馬鈴薯植株莖基部腐爛等癥狀,曾被認(rèn)為是造成德蘭士瓦馬鈴薯枯萎病的主要病原菌(Theron & Holz,1989)。而本試驗(yàn)中木賊鐮刀菌3-2 引起的馬鈴薯枯萎病為國(guó)內(nèi)首次報(bào)道。三線鐮刀菌是鐮刀菌屬中一類(lèi)重要的病原菌,目前在中國(guó)主要發(fā)生在內(nèi)蒙古(陳慧 等,2016)和青海(楊波 等,2019),而在甘肅省由三線鐮刀菌引起的馬鈴薯枯萎病為首次報(bào)道。陳慧等(2016)報(bào)道,三線鐮刀菌接種后于馬鈴薯生長(zhǎng)的中后期發(fā)病,發(fā)病初期植株下部葉片萎蔫;發(fā)展后期,葉片由下而上逐漸萎蔫枯死,與本試驗(yàn)結(jié)果相同。其中三線鐮刀菌3-6 引起的病害癥狀表現(xiàn)為馬鈴薯植株下部葉片枯萎,而三線鐮刀菌1-1 引起的病害癥狀為整個(gè)馬鈴薯植物萎蔫枯死,說(shuō)明同一屬的不同菌株間存在差異性,具有遺傳多樣性。曹宜等(2004)曾分析了22 株尖孢鐮刀菌遺傳物質(zhì)的差異性,利用分子生物學(xué)手段證明了不同基因的表達(dá)結(jié)果是造成尖孢鐮刀菌致病力存在差異的主要原因。因此,三線鐮刀菌中也可能存在著同樣的差異性基因表達(dá),導(dǎo)致其不同菌株間的致病性存在差異性。
木賊鐮刀菌菌株3-2 菌落呈淡米黃色、疏松、棉絮狀,分生孢子梗單生具分枝,大分生孢子呈鐮刀形,3~5 個(gè)分隔,與文獻(xiàn)中報(bào)道的木賊鐮刀菌的培養(yǎng)性狀相似(Gachango et al.,2012)。三線鐮刀菌菌株1-1 的菌落形態(tài)為菌落表面中心由白色逐漸變?yōu)榧t色,邊緣呈不規(guī)則狀;菌株3-6 的菌落形態(tài)為菌絲紅色,生長(zhǎng)后期菌落中心表面呈黃色,二者之間培養(yǎng)性狀存在差異性。但也有相關(guān)報(bào)道表明,鐮刀菌對(duì)環(huán)境的變化較敏感,容易受到溫度、酸堿度、海拔等因素的影響(O’Donnell et al.,1998;郝蓉蓉 等,2015)。因此,通過(guò)常規(guī)的形態(tài)學(xué)鑒定很難將鐮刀菌確定到種。本試驗(yàn)進(jìn)一步通過(guò)分子生物學(xué)手段將菌株1-1 和3-6 最終均鑒定為三線鐮刀菌,說(shuō)明分子生物學(xué)在復(fù)雜種的鑒定過(guò)程中具有重要的作用。
本試驗(yàn)明確了山丹縣馬鈴薯枯萎病病原菌的種類(lèi),可以為甘肅省山丹縣馬鈴薯枯萎病的田間診斷和綜合防治提供理論依據(jù)。
