馬克斯克魯維酵母菌調控酒精飲料風味物質代謝機制的研究進展
2024-02-28 03:10:56侯雨辰烏日娜曹愷欣王雨生安飛宇武俊瑞
中國釀造 2024年1期
侯雨辰,烏日娜,曹愷欣,王雨生,安飛宇,賀 揚,武俊瑞*
(1.沈陽農業大學 食品學院 遼寧省食品發酵技術工程研究中心 沈陽市微生物發酵技術創新重點實驗室,遼寧 沈陽 110866;2.啤酒生物發酵工程國家重點實驗室,山東 青島 266100;3.青島啤酒股份有限公司,山東 青島 266100)
酒精飲料深受廣大消費者青睞。風味品質是決定酒精飲料價值的重要依據,而酒精飲料發酵所用的酵母菌在其風味物質形成過程中發揮著極其重要的作用[1]。目前常用的傳統酵母菌主要為釀酒酵母(Saccharomyces cerevisiae),但研究發現,生產中S.cerevisiae與馬克斯克魯維酵母菌(Kluyveromyces marxianus)等野生酵母菌混合發酵可大大提高酒精飲料的風味品質[2]。
馬克斯克魯維酵母菌是一種在乳制品(開菲爾)中被分離得到的食品級安全酵母,被用于發酵食品、產酶、生產果糖、生物修復以及生產各種細胞產品等,且在系統發育上與S.cerevisiae相關,與使用較廣泛的乳酸克魯維酵母(Kluyveromyces lactis)是姊妹種[3]。此外已有大量研究表明,K.marxianus可代謝多種碳糖(葡萄糖、半乳糖、乳糖和菊糖等),從而能夠對釀酒原料所含有的多種糖類進行反應。其生長速度迅速且能在38~45 ℃生存,具有突出的耐熱性和乙醇發酵能力,以及具有產生多種芳香化合物(主要是酯類和醇類)和酶的能力,能夠改善酒的風味和口感,提升品質,從而使這種微生物在食品加工業中成為一種合適的生物技術工具[4-5]。目前國內外研究者已經意識到這種微生物在產香方面的商業價值,對其代謝芳香化合物的機制進行了探究,然而,對其在實際應用中如何發揮最大發酵性能,使其產品的風味更豐滿的探究并不深入,很多相關調控技術仍未能研究清晰,大大影響了K.marxianus在釀酒行業的投入使用。本文擬分析現有K.marxianus發酵酒精飲料產主要風味物質的代謝機制,探討其釀酒發酵風味的調控技術以及闡述了K.marxianus在酒精飲料應用的發展趨勢。希望為通過影響發酵風味的調控技術解決酒精飲料風味不足的問題提供有益的借鑒與參考。
1 K.marxianus對酒精飲料特征風味物質的代謝機制
酒精飲料的香氣是由多種揮發性風味物質共同決定的。這些風味物質是在各種酶的作用下通過酵母細胞的合成代謝和分解代謝途徑生成的,主要包括醇類、酯類、醛類、酸類、酚類等,其中高級醇和乙酸酯是影響酒精飲料特殊風味的兩種重要化合物,且不同的酵母菌風味物質代謝途徑的活性有一定的差異。高級醇提供“背景風味”,也是酯類形成的重要前體物質。乙酸酯及其他酯類會影響酒精飲料的果香和花香風味。目前研究者對K.marxianus的代謝途徑研究不夠充分,而對釀酒酵母研究較為詳細,可為研究K.marxianus的基因、蛋白質、代謝途徑奠定基礎[6]。已知K.marxianus生產的芳香化合物被集中研究的有2-苯乙醇(2-phenethyl alcohol,2-PE)、2-苯乙基乙酸酯(2-phenethyl acetate,2-PEA)和乙酸乙酯[7]。
1.1 K.marxianus代謝合成苯乙醇途徑
2-苯乙醇是一種酵母衍生化合物,已被證明會影響發酵飲料的香氣和風味,呈玫瑰狀香氣,還可作為酒用香料額外添加到各類酒精飲料中[8]。酵母菌的2-PE生物合成主要包括莽草酸途徑(shikimate pathway)和埃爾利希途徑(Ehrlichpathway),其中最能有效合成的是Ehrlich途徑(圖1)。對于通過發酵生產2-PE,研究主要集中在S.cerevisiae和K.marxianus。這兩種酵母菌均能通過莽草酸途徑從糖從頭合成2-PE,還可通過埃爾利希途徑從L-苯丙氨酸(L-phenylalanine,L-Phe)生產更高含量的2-PE[9-10]。
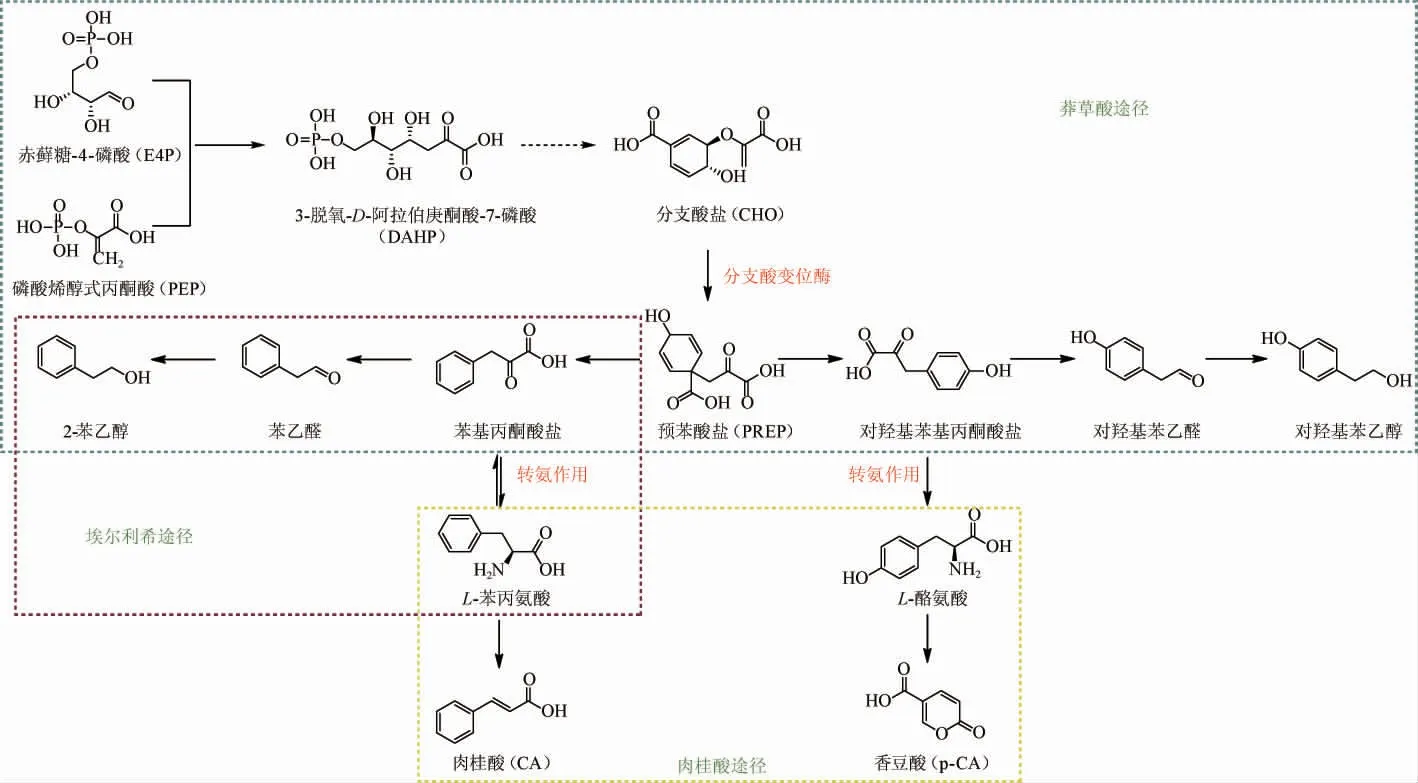
圖1 馬克斯克魯維酵母菌代謝合成苯乙醇途徑Fig.1 Metabolic pathway of phenylethanol synthesis by Kluyveromyces marxianus
1.1.1 莽草酸途徑
莽草酸途徑是由磷酸烯醇式丙酮酸(phosphoenolpyruvate,PEP)和赤蘚糖-4-磷酸(erythrose-4-phosphate,E4P)縮合形成3-脫氧-D-阿拉伯庚酮酸-7-磷酸(3-deoxy-D-arabinoheptulosonic acid-7-phosphate,DAHP)而啟動并通過一系列的酶促反應最終生成醇類[11]。DAHP通過一系列反應會生成分支酸鹽,再由分支酸變位酶進行催化生成預苯酸鹽。在該途徑中存在的兩種芳族2-含氧酸分別為苯基丙酮酸鹽或對羥基丙酮酸鹽是由預苯酸鹽轉化得來的,可通過進行轉氨作用分別產生L-苯丙氨酸和L-酪氨酸,也可通過2-含氧酸脫羧酶的酶促反應生成苯乙醛或對羥基苯乙醛,進而再轉化成苯乙醇或對羥基苯乙醇。然而,該途徑涉及的反應復雜且存在多種抑制作用,產生的2-PE含量很低,因此不是酵母菌產2-PE的主要途徑[12]。
1.1.2 埃爾利希途徑
酵母生成高級醇主要通過Ehrlich途徑。該途徑是由埃利希在1906年首次提出的,且證明了高級醇是由氨基酸的分解代謝合成的[13]。K.marxianus通過轉化不同的氨基酸底物,可以轉化成不同的高級醇,如亮氨酸、蛋氨酸和L-苯丙氨酸轉化成它們各自的雜醇:異戊醇、甲硫醇和2-苯乙醇[14]。如圖1所示,K.marxianus生產的2-PE來源于L-Phe經氨基酸轉氨酶生成的苯基丙酮酸。苯基丙酮酸的脫羧是埃利希途徑的開始,經由α-酮酸脫羧酶去除碳酸基形成苯乙醛,最后由氧化還原酶還原成2-PE[15]。然而,當有除L-Phe外其他更容易消化的氮源存在時,L-Phe更容易通過肉桂酸途徑代謝,從而會限制2-PE的形成[16-17]。
1.2 K.marxianus代謝合成乙酸酯及乙醇途徑
大量研究表明,酯類物質是酒類風味與品質的關鍵化合物之一,其中乙酸乙酯是最常見的酯,為白酒的四大酯之一,同時在葡萄酒的果香感知中起著關鍵作用[13]。在釀酒條件下,乙酸酯多由非釀酒酵母產生,是以生物合成為主[18]。K.marxianus在酯的生產方面有巨大的潛力,但在食品領域關于其生產乙酸酯的報道提及較少,同時該菌的風味積累代謝不如S.cerevisiae那樣被人所知[19]。K.marxianus代謝產生最重要的酯是2-PEA、乙酸異戊酯和乙酸乙酯,分別由2-PE、異戊醇和乙醇合成[6]。
K.marxianus的代謝合成乙醇及乙酸酯途徑具體表現為碳源底物經酵母菌的糖酵解途徑生成丙酮酸鹽,在丙酮酸鹽脫羧酶的催化下生成乙醛,又通過脫氫酶生成乙醇經酯交換最終形成乙酸乙酯,而其他乙酸酯的生成則是由Ehrlich途徑生成的高級醇酯化生成的(圖2)[6]。另外,在醇類物質進行酯交換時,主要是乙醇或其他雜醇與乙酰輔酶A通過乙醇乙酰基轉移酶(alcohol acetyltransferase,AATase)活性進行反應的,因此決定該菌生成乙酸酯的影響因素包括底物醇的種類和含量、乙酰輔酶A(coenzyme A,CoA)的水平以及AATase的表達[20]。
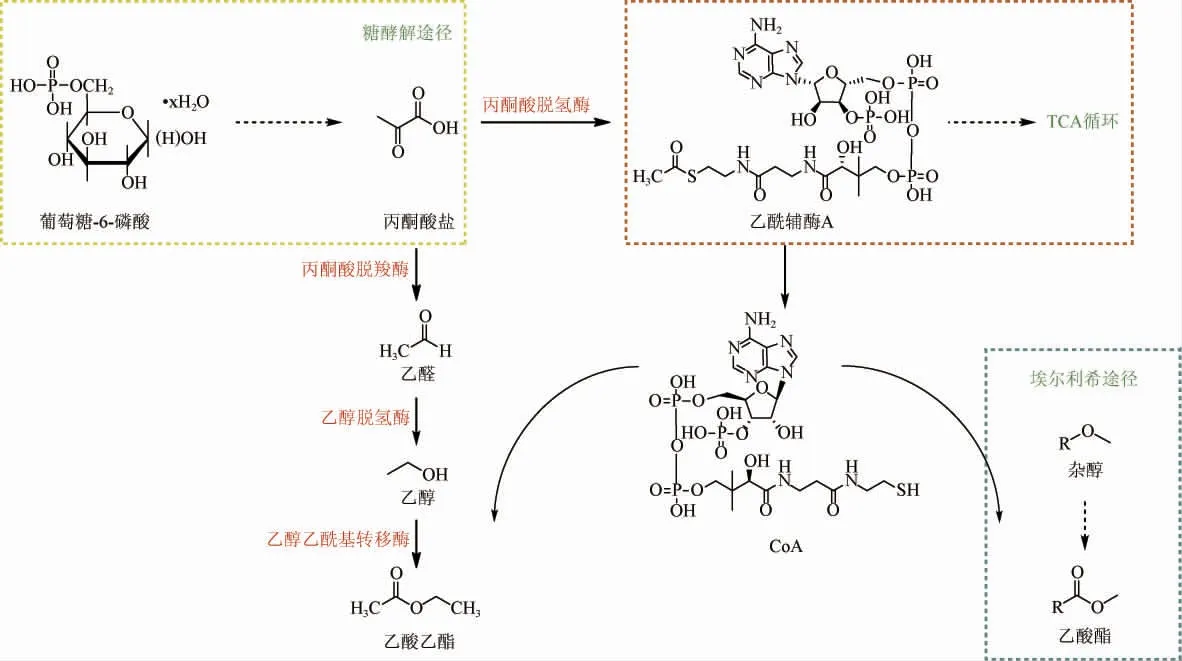
圖2 馬克斯克魯維酵母菌代謝合成乙醇及乙酸酯途徑Fig.2 Metabolic pathway of ethanol and acetate synthesis by Kluyveromyces marxianus
合成的芳香化合物是每個微生物的特定代謝所固有的,合成代謝過程中涉及到的酶即使在不同類型中也具有相同的活性。K.marxianus和釀酒酵母均含有編碼AATases的基因,其可變表達是確定發酵后最終形成酯的重要因素,并且組成AATase的基因主要是醇乙酰基轉移酶編碼基因ATF1和ATF2[21],而乙酰輔酶A主要有兩種途徑合成:①通過糖酵解(glycolytic pathway,EMP)途徑生成的丙酮酸鹽進入線粒體由丙酮酸脫氫酶(pyruvate dehydrogenase,PDH)促進生成的,然后可以進入三羧酸循環(tricarboxylic acid cycle,TCA);②丙酮酸鹽通過丙酮酸脫羧酶催化生成乙醛,再由乙醛脫氫酶形成乙酸,最后可通過乙酰輔酶A合酶生成[22]。在蘋果酒發酵中對比于S.cerevisiae,K.marxianus能降低乙酰輔酶A合成從乙酸鹽和乙酰輔酶A進入TCA的代謝通量,從而增加乙酸乙酯和相關酯的合成[23]。
2 K.marxianus發酵酒精飲料的風味調控技術
基于K.marxianus具有代謝多種碳糖、突出的耐熱性和乙醇發酵能力以及生成醇類、酯類等風味物質的特性,將其應用于酒精飲料來達到改善風味的作用。為能使這些優良特性被充分的利用,明悉其釀酒發酵風味的調控技術顯得尤為重要。在實際生產中,酒精飲料的風味調控因素包括在發酵過程中的接種方式、發酵溫度以及最容易被忽視的營養素的添加等。
2.1 通過K.marxianus接種工藝調控發酵酒精飲料風味
2.1.1 接種方式對風味的調控
K.marxianus在釀酒的研究中分為單菌發酵和混菌發酵兩種方式。然而K.marxianus單菌接種在實際生產中使用較少,大多是以與S.cerevisiae或乳酸菌混合發酵的形式進行的。但對于馬圭果汁和龍舌蘭汁這些含糖種類多且不易被S.cerevisiae利用的原料,采用K.marxianus單菌發酵的效果更佳[24]。另外,K.marxianus的代謝通量是針對乙醇發酵而不是可以支持細胞生長的乙酰輔酶A合成,而在S.cerevisiae中,乙醇發酵和乙酰輔酶A合成途徑均具有高度活性,因此K.marxianus單菌發酵的時間要比S.cerevisiae發酵酒精飲料所需的時間長[23]。
混菌發酵可以在提高發酵酒風味特性的同時高效率完成發酵過程,在生產中較為常見。采用混合發酵時,K.marxianus在發酵前期積累各種風味物質,隨著發酵的進行會受到S.cerevisiae代謝產物的抑制作用、營養物質競爭作用等,其活菌數量會迅速下降[25]。然而兩種菌的風味物質的積累會產生獨特的風味,因此在利用K.marxianus生產不同酒精飲料時,工廠要根據發酵原料的不同選擇適當的接種方式。
2.1.2 混菌發酵接種順序和初始比例對風味的調控
采用混菌發酵方式時,發酵動力學的差異取決于不同菌種之間的接種順序和初始比例,其會強烈影響發酵過程中酵母菌種的優勢和持久性[26]。恰當的接種方式和初始比例才能使每種酵母菌均能發揮出最大的發酵性能,增加芳香化合物的種類和含量。
目前關于非釀酒酵母和釀酒酵母菌株主要有兩種接種方法:①同時接種,即在發酵前同時接種兩種酵母菌。該接種方式在黃酒發酵的應用中較為常見;其接種比例在1∶1和1∶20的共發酵試驗中可以完成發酵,而在較高接種比例下可能存在發酵緩慢或停滯的風險[27]。劉夢[28]研究發現,K.marxianus與釀酒酵母1∶1同時接種發酵的黃酒口感最佳。②順序接種,通常先接種非釀酒酵母,在發酵前期充分積累風味物質,間隔一段時間后接種釀酒酵母。順序接種這種方式能夠延長非釀酒酵母在整個發酵體系中的存活時間,縮短酒精發酵完成的時間,并能更有效地改善酒的品質和感官特性,因此在進行混菌發酵時順序接種是最常使用的方法,大多數間隔48 h為最佳[29-30]。然而,使用兩次接種會增加釀酒廠制酒的操作時間和操作步驟,期間易造成污染,在操作時也要著重注意污染的問題。特別的是,K.marxianus的耐熱性有利于在高溫下減少前期接種造成污染的機會,因此在進行接種時,釀酒廠可考慮在高溫下進行操作[31]。
2.2 通過K.marxianus發酵溫度調控發酵酒精飲料風味
酵母菌的耐熱性是酒精飲料工業發酵的關鍵特性。在一定程度上,升高溫度能提高酵母菌胞內酶的活性、膜的流動性和生長代謝速率,能夠加速風味物質的合成,形成不同種類和數量的酒類風味代謝產物[32]。以白酒為例,在入池發酵過程中由于微生物生長繁殖會產生生物熱,發酵的實際溫度最高可上升到45 ℃左右[33]。在發酵過程中,酵母的過早死亡可能導致高殘余糖的停滯發酵,那么在眾多可進行酒精發酵的非釀酒酵母菌株中,耐熱性可以作為這些酵母菌的篩選指標特性。
FU X F等[34]研究發現,K.marxianus在30~45 ℃的溫度下葡萄糖消耗量和乙醇產量在發酵的早、中期幾乎沒有差異,而在45 ℃的溫度下的乙醇合成會在發酵晚期停滯。K.marxianus具有能在4~52 ℃的溫度范圍內生長(即耐熱性高)的特性,有利于其在更高溫度下進行同步糖化和發酵等步驟,也可防止熱敏感的微生物生長,減少發酵過程中微生物的污染[4]。LI P S等[35]在高溫發酵條件下對K.marxianus進行代謝組學的分析,發現該菌在高溫發酵停滯后大部分代謝途徑停止,但上調代謝物的途徑富集分析顯示與核黃素和氨基酸(甘氨酸、絲氨酸、蘇氨酸、丙氨酸、天冬氨酸和谷氨酸)代謝相關的途徑顯著富集,表明高溫條件下通過Ehrlich途徑由不同氨基酸生成高級醇的相關代謝會顯著加強。這種情況可通過令K.marxianus經受溫度逐漸升高的重復長期培養,來提高其在高溫發酵產乙醇的能力[36]。
2.3 通過K.marxianus營養需求調控發酵酒精飲料風味
酵母需要多種營養物質來維持其生存、生長和持續代謝活動。營養物質的組成和濃度對酵母菌進行酒精發酵和芳香物質的代謝有非常重要的作用。碳源是酵母菌產生CO2以及生物質或其他非酒精副產物的源頭,目前已被大量研究,但氮源、維生素和其他微量元素也值得關注,這些營養物質的缺失會導致停滯或發酵緩慢。K.marxianus和S.cerevisiae之間存在對各種營養物質的競爭作用,且對營養物質的偏好不同。ROLLERO S等[37]證明了K.marxianus對氮源以及其他營養物質具有物種依賴性競爭,還強調了控制營養物濃度的重要性。因此,研究其他營養物質種類或濃度對K.marxianus合成風味物質的影響是有必要的。
2.3.1 氮源
氮是酵母發酵所需的重要營養物質之一,可以增加生物量的產量,刺激糖的利用率,而氮的缺乏直接影響酵母的發酵性能,尤其是對雜醇生產的影響顯著[38]。因此,酵母同化氮化合物的可用性和性質是工藝管理的關鍵因素,可以通過添加不同的有機或無機來源來簡單控制。
有研究發現,K.marxianus與S.cerevisiae表現出相同的氮源偏好,比如可消耗γ-氨基丁酸或少量銨[39]。氮源對2-PEA的水平有強烈的影響,當氮源是銨時,可以起到抑制AATase表達或活性的作用,此時只有非常少量的2-PEA被合成但同樣由AATase活性合成的乙酸異戊酯沒明顯的N-效應,因為除2-PEA外,乙酸乙酯和其他重要的合成酯類的合成路線是碳代謝,而不是氮代謝;當使用葡萄糖/銨混合物時K.marxianus的生長速度會加快,另外使用酵母抽提物時,醇的含量會升高[20]。由于2-PE的天然合成水平相當低,由2-PE的代謝途徑可知通過添加外源L-Phe,尤其是作為唯一的氮源,也會導致2-PE水平大大增加。
2.3.2 維生素
已知增加某些維生素的濃度可以改善葡萄酒發酵過程中酵母的發酵動力學和細胞活力。與所研究的其他營養素相比,維生素B1(硫胺素)對K.marxianus的發酵性能影響最為顯著。硫胺素是一種特殊的水溶性維生素,在酒精飲料發酵過程中維持充足的外源添加水平對酵母發酵性能的有益影響已被充分報道[40]。酵母在某些環境條件下合成硫胺素的能力較差,如在葡萄酒發酵過程中,酵母需要足夠的外源硫胺素來誘導最大的生長和發酵速率。在模擬釀酒條件下硫胺素能提高K.marxianus的發酵性能,是這種酵母菌維持最大細胞活力和發酵性能的必要條件,在0.25 mg/L硫胺素的飽和閾值時,表現對該酵母菌發酵性能的最大影響[41]。另外,維生素B7(生物素)也可能影響K.marxianus的生長代謝。METHNER Y等[42]研究發現,K.marxianus無法在啤酒中釋放生物素,反而可能將麥汁原本所含的生物素用于自身的新陳代謝中,因此生物素也可能是K.marxianus的重要生長因子。
2.3.3 微量元素和應激因子
一些微量元素(比如鎂、鐵、鋅)和應激因子(真菌毒素、黃曲霉素等)對K.marxianus的發酵效率有一定的影響,能夠影響酵母生長、發酵速率、獲得的乙醇產量、酵母絮凝行為和發酵期間對外界壓力的耐受性。
L?SER C等[43]通過K.marxianus發酵乳清殘渣轉化生產乙酸乙酯,說明了鐵在這個過程中起重要作用,證明鐵吸收動力學對于理解和控制酯合成過程具有重要的實際意義。另外,GSCHAEDLER A等[44]在K.marxianus發酵蘋果酒的體系中添加鎂、鋅和氮后乙醇產量大量增加,證明這些營養物質是發酵所必需的,此外,又發現在不添加任何營養物質的情況下,K.marxianus是唯一能產生更多揮發性化合物的酵母。因此,從發酵酒的應用來看,微量元素也是影響K.marxianus發酵的較為重要的營養元素。
一些應激因子會影響酵母的發酵性能,如污染發酵培養基的真菌毒素。盡管許多酵母可以適應不利的生長條件,這種極端條件可能導致細胞死亡或生長減緩,還可能對產量和發酵指標產生不利影響[45]。JAKOPOVIC′? 等[46]研究了黃曲霉毒素B1、赭曲霉毒素A和玉米赤霉烯酮對釀酒酵母和K.marxianus形態、生長參數和代謝活性的影響發現,在發酵的早期階段,所有暴露于真菌毒素的酵母代謝活性和生物量生長都比對照低,但是發酵產物和生物量濃度在發酵結束時達到對照水平。
綜上所述,發酵酒精飲料風味調控技術是提升產品風味品質的關鍵,不僅局限于接種方式、比例和溫度等,也要注重額外營養素的添加,歸根結底就是要安全且能提高菌株本身新陳代謝。采取適當的調控技術有利于充分發揮K.marxianus菌株的發酵性能,可縮短發酵時間、節約成本,更有利于減少K.marxianus停留在實驗室階段而很少實際應用的局限,有利于其在酒精飲料的實際投入生產。
3 K.marxianus在酒精飲料的應用
3.1 在奶啤中的應用
奶啤是以牛乳為主要原料,由乳酸菌、酵母菌兩步發酵而成的一種低酒精度、具有碳酸沙口感且質感細膩的新型乳飲料[47]。生產所使用的酵母菌能影響醇、酸、酯類等主要風味物質的產生,是奶啤產品發酵的關鍵菌種[48]。目前奶啤市場存在風味單一的問題,因此企業一直在尋找代謝產物豐富、酒精產量低、適合直接發酵的乳源酵母菌種作為發酵替代菌株,來降低成本,簡化工藝,增加風味。何明迎[49]將K.marxianus應用到奶啤的發酵中,并對發酵工藝及穩定性進行優化,發現優化后的奶啤中的各種風味物質及其相對含量均顯著提高,尤其是乙酸酯類和醇類相對含量顯著增加,使香氣成分更豐富,風味品質得到明顯的提高。然而,奶啤的菌種優化和產香機制研究處于初步研究階段,仍需研究者進行進一步的探究。
3.2 在果酒中的應用
K.marxianus在葡萄酒發酵過程中具有高果膠酶活性,并且相比于釀酒酵母(S.cerevisiae)能產生更多的芳香化合物(如苯乙醇、乙酸異戊酯和乙酸乙酯等),使葡萄酒具有更多的果味或花香特征[50]。該菌不僅廣泛應用于葡萄酒,也被應用于其他果酒的發酵。陳穎等[51]研究發現,經過K.marxianus發酵的柿子酒可提高芳香化合物的含量,還可生成新的高級醇、酯和酮類化合物,從而提升酒的品質。國外研究者發現K.marxianus對于馬圭果汁具有高發酵能力,且發酵產生的乙醇濃度、乙醇產量、效率高于S.cerevisiae,被認為是手工生產馬圭果汁酒的合適工具[52]。另外,對K.marxianus和S.cerevisiae發酵的蘋果酒中的揮發性和非揮發性成分進行轉錄組分析,發現K.marxianus更能增加蘋果汁發酵過程中高級醇、脂肪酸和酯的含量,尤其是提高乙酸乙酯、己酸乙酯、癸酸乙酯等乙酸酯的產量[23]。這些研究均可證明K.marxianus在不同種果酒發酵的應用潛力之巨大,可增加其對不同品種水果發酵酒產品的開發,以便在果酒產業中對該菌種的拓展使用。
3.3 在其他酒精飲料中的應用
K.marxianus也被應用于龍舌蘭酒、乳清酒和米酸酒。研究發現以不同種類的龍舌蘭物種為原料發酵蒸餾成酒時,S.cerevisiae不能發酵全部種類的龍舌蘭汁,而K.marxianus能夠全部發酵,且可以根據發酵的不同底物能產生高水平的乙醇、乙酸乙酯、高級醇和乙醛,比S.cerevisiae更適于某些龍舌蘭酒的發酵[25]。已有研究表明,K.marxianus具有通過β-半乳糖苷酶代謝乳糖的能力,是進行乳清發酵的理想候選菌株,其在發酵酸乳清時和S.cerevisiae表現出相似的發酵速率,且K.marxianus產生了特殊的化學香氣[53]。
無論是單菌還是混菌發酵試驗,K.marxianus會影響酒精飲料中具有芳香特征成分的種類與含量。然而,絕大多數研究還停留在芳香物質數量上的變化,對影響風味的產香機制研究還遠遠不夠,尤其是區別于不同種類、不同風格的酒精飲料關鍵風味物質的追蹤。
4 展望
K.marxianus作為一種能夠利用多種碳源,增加酒精飲料的酯香、醇香,同時具有優秀的發酵能力的食品級安全產香酵母菌,在各類酒精飲料的實際應用逐漸增多。目前研究者在代謝方面對K.marxianus產香機制的探索相對較少,應基于基因組、轉錄組等組學信息和代謝建模數據來提高其相關生產力,尤其要填補在基因、酶和其他風味物質代謝通路及調控方面的知識空白,以便更好地應于食品領域。在酒精飲料發酵風味的調控技術中,研究者往往容易忽視營養素的添加。確定對K.marxianus菌株發酵性能至關重要的營養素,并隨后確保足夠的營養素水平是酒精飲料風味品質提升的一大關鍵,但不同種類的酒精飲料其具體關鍵風味物質和發酵條件有一定的差異,如何快速找到有效調控產香機制的關鍵技術仍需進一步研究。另外,在混合發酵的實際應用中不同菌株之間的相互作用是復雜多樣的,因此利用組學技術分析蛋白、基因等表達差異研究K.marxianus與釀酒酵母或者乳酸菌之間代謝、生物量的相互作用機制是至關重要的。最后,利用K.marxianus能合成多種芳香化合物的特性,可考慮拓展其在生產香料和其他發酵食品的應用。未來應加強其在酒精飲料的實際應用,應充分挖掘該菌株的優良釀酒特性,并利用適當的調控技術使其發揮出最好的發酵性能,生產出風味品質更佳的酒類產品,以促進我國釀酒產業的良性發展。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
中國衛生(2014年12期)2014-11-12 13:12:52
食品工業科技(2014年9期)2014-03-11 18:15:31
