低溫β-1,4-內切葡聚糖酶的基因挖掘、異源表達及其酶學性質表征
2024-03-01 12:55:56圣弟青鈕成拓閆亞楠鄭飛云劉春鳳王金晶李崎
食品與發酵工業 2024年4期
圣弟青,鈕成拓,閆亞楠,鄭飛云,劉春鳳,王金晶,李崎
(工業生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122)
β-1,4-內切葡聚糖酶(EC3.2.1.4)是纖維素酶系中的一種,能夠隨機作用于纖維素無定形區中的β-1,4-糖苷鍵,將纖維素分子水解成長度不同的寡糖鏈[1]。纖維素酶的應用十分廣泛,在洗滌、生物質能源等行業具有廣泛的用途。在生物質能源中,β-1,4-內切葡聚糖酶可以在其他酶類的協同作用下降解纖維素為寡糖,從而最終轉化為生物乙醇[2]。在洗滌行業,β-1,4-內切葡聚糖酶添加在洗滌劑中可以提升洗滌劑的洗滌效率,同時提高棉質衣物的光亮度改善棉纖維的柔軟性,保護棉紡織物等作用[3-4],因此具有很高的商業價值。最早被應用在洗滌行業的β-1,4-內切葡聚糖酶來自BacillusKSM-635,首先由ITO[4]報道,最適溫度40 ℃,最適pH值為9.5,在pH 11.0保持70%以上活性,目前為花王公司生產的Biotex。因為β-1,4-內切葡聚糖酶在洗滌行業上的應用環境通常都比較苛刻,通常需要其具有低溫、耐堿以及抗表面活性劑等穩定的酶學性質。目前,對于β-1,4-內切葡聚糖酶的研究工作大部分集中在提高酶活力熱穩定性等方向研究,并應用在飼料加工和生物能源等領域[5-6],對于可應用在洗滌紡織行業的β-1,4-內切葡聚糖酶的研究相對較少,并且對應用在洗滌行業的β-1,4-內切葡聚糖酶通常需要具有低溫和耐堿性的酶學特征,因此,獲得具有低溫和堿性耐受能力的β-1,4-內切葡聚糖酶有利于提升其在洗滌行業的應用價值[7]。
β-1,4-內切葡聚糖酶的分布十分廣泛,在古細菌、細菌、真菌、昆蟲和動物胃腸道中均有報道[8-10]。然而,不同家族來源β-1,4-內切葡聚糖酶的酶學性質也有很大差異[11]。例如,真菌來源的β-1,4-內切葡聚糖酶通常具有較好的熱穩定性和催化活性,且能夠實現胞外分泌生產,在能源、飼料行業具有廣泛的應用[12]。但是,真菌酶的最適pH值一般呈中性或偏酸性,無法在洗滌行業進行應用。在食草動物體內也分離得到一些β-1,4-內切葡聚糖酶基因,但是性質差別較大,且較難分離利用[5]。與之相比,細菌來源β-1,4-內切葡聚糖酶一般具有較寬的pH范圍和溫度范圍,且具有易分離提取的特點,因此目前在洗滌行業中應用的β-1,4-內切葡聚糖酶大多來源于細菌。例如,最早應用于洗滌劑中的工業酶就分離自堿性環境的嗜堿芽孢桿菌[13-14]。
基于高通量測序方法的更新與計算方法的發展,通過對已獲得基因組的挖掘已經成為發現新型天然酶的一種技術手段,而生物信息學的發展能夠進一步解析新發現酶的相關信息[15]。截止到2022年12月,已有241 015 745條基因序列被測序分析并記錄在GenBank中,雖然其中有很多基因序列暫時沒有被注釋所編碼的蛋白質特性與功能,但是很多氨基酸序列已被解析出功能和特征,為具有獨特酶學特性的新酶挖掘提供了寶貴的資源。目前通過基因挖掘獲得新酶,并對其酶學性質進行表征已成為獲得新酶的一個重要研究方向[16-17]。SANANGELANTONI等[18]通過基因挖掘獲得了一種熱穩定性高且底物特異性強的新型β-螺旋植酸酶;周茂志等[19]通過基因組數據挖掘獲得了9個新的扁桃酸消旋酶基因及活性蛋白,并成功異源表達出來源于放射性土壤桿菌Agrobacteriumradiobacter的ArMR 酶。通過基因挖掘手段也可以獲得新型β-1,4-內切葡聚糖酶,LIN等[20]通過基因挖掘成功從MicrobacteriumkitamienseS12中挖掘得到一種最適溫度為35 ℃的新型冷適應性β-1,4-內切葡聚糖酶,其在0~5 ℃能夠保持30%~40%的相對酶活;DHUME等[21]從MrakiarobertiiA2-3挖掘得到的耐寒內切葡聚糖酶的最適溫度為15 ℃,而比酶活力到17.38 U/mg,具有在洗滌行業實際應用的潛力。
因此,本論文通過基因挖掘手段篩選得到一種新的低溫β-1,4-內切葡聚糖酶基因,并在EscherichiacoliBL21中實現異源可溶性表達;在此基礎上,對新酶進行了表達和純化,并表征了其酶學性質,為其在洗滌和紡織行業的應用奠定基礎。
1 材料與方法
1.1 材料與設備
通過基因挖掘得到的β-1,4-內切葡聚糖酶基因(Genbank No.WP_246130614)命名為EgDC,由安生達(蘇州)公司經過密碼子優化后合成,并連接在pET-22b(+)載體,在重組蛋白N端連接His-tag標簽。DNA Marker、E.coliBL21(DE3)感受態細胞、高保真酶、限制性內切酶、T4連接酶等分子試劑,大連TaKaRa公司;Ni-NTA純化樹脂預裝柱、Bradford法蛋白濃度測定試劑盒、SDS-PAGE快速制備試劑盒等,生物工程(上海)有限公司;氯化鈉、甘油、胰蛋白胨、酵母粉、用作底物的羧甲基纖維素鈉(Carboxymethylcellulose sodium,CMC-Na)等,國藥集團化學試劑有限公司。
紫外-可見光分光光度計,上海MAPADA 儀器有限公司;FiveEasy pH 計,Mettler Toledo(上海)有限公司;冷凍離心機,德國Eppendorf 公司;恒溫水浴鍋,金壇市宏華儀器廠;Powerpac Basic 型蛋白電泳儀,美國BIO-RAD 公司;電子天平,奧豪斯儀器(常州)有限公司;SPX-250 生化培養箱,上海躍進醫療器械廠;MR-96T型智能酶標儀,騁克儀器(上海)有限公司;全自動高壓蒸汽滅菌鍋,日本HIRAYAMA株式會社;PL2002電子天平,METTLER TOLEDO公司;10 μL-1 mL不同型號移液槍,德國Eppendorf 公司;Sonics Vibra-Celltm超聲破碎儀,美國Sonics公司;GeIDoc-ItTM 凝膠成像系統,美國UVP公司;MICROL 17臺式高速離心機,美國Thermo Scientific公司。
1.2 實驗方法
1.2.1 基因挖掘和序列分析方法
基因挖掘出發序列在碳水化合物活性酶數據庫pfam官網(http://pfam.xfam.org/)進行檢索;在NCBI數據庫(https://www.ncbi.nlm.nih.gov/)進行blast對比搜索;利用Mega X 程序基于連接法構建進化樹并在iTOL網站(https://itol.embl.de/)進行圖片展示;利用clustal W程序對得到β-1,4-內切葡聚糖酶氨基酸序列進行多序列比對;采用ESPript 在線服務器(http://espript.ibcp.fr/ESPript/ESPript/)進行可視化結果分析。
1.2.2 重組β-1,4-內切葡聚糖酶的構建
重組大腸桿菌的構建通過安生達(蘇州)公司進行挖掘得到的β-1,4-內切葡聚糖酶基因密碼子優化、合成后在其N段連接his-tag標簽,并連接至pET-22b(+)質粒。通過雙酶切驗證重組質粒正確性后,通過化學轉化的方法將其轉化進入E.coli感受態細胞中,并涂布于含硫酸卡那霉素的LB固態平板。在此基礎上,通過將克隆轉接至液體LB培養基后提取質粒,并通過雙酶切和基因測序驗證確定陽性克隆。
1.2.3 重組大腸桿菌的誘導表達及純化
從平板挑取陽性轉化子接種至含100 μg/mL氨芐青霉素的TB液體培養基,于37 ℃、40×g下振蕩培養至OD600值為0.7~0.8后,加入終濃度1 mmol/L 異丙基-β-D-硫代半乳糖苷(isopropyl-beta-D-thiogalactopyranoside,IPTG)誘導表達,并設置不添加IPTG誘導的對照組,在20 ℃培養48 h。取培養菌液40 mL于4 000×g離心5 min收集菌體,加入30 mL Tris-HCl緩沖液(20 mmol/L,pH 7.0)重懸菌體,并調節OD600值為5。在冰浴條件下通過超聲波破碎儀(500 W/750 W,美國Sonics公司)破碎細胞。破碎條件如下:300 W,超聲3 s、間隔7 s,30 min。破碎后得到的液體為全細胞裂解液,經4 000×g離心10 min后收集上清液,即為粗酶液。
β-1,4-內切葡聚糖酶的純化采用Ni-NTA預裝重力柱純化(上海生工生物工程有限公司)。通過梯度洗脫確定目標蛋白洗脫緩沖液咪唑濃度后,在最優咪唑濃度下洗脫目標蛋白,并采用截留分子質量為3 000 Da的蛋白超濾管濃縮洗脫液,經PD-10脫鹽柱換鹽后,最終獲得純化后蛋白樣品。純化后蛋白樣品通過SDS-PAGE進行分析。
1.2.4 β-1,4-內切葡聚糖酶的酶活力測定
β-1,4-內切葡聚糖酶的酶活力測定方法參考3,5-二硝基水楊酸試劑(3,5-dinitrosalicylic acid,DNS)法[22]。取1.5 mL CMC-Na底物溶液于EP管中,40 ℃恒溫水浴5 min,加入0.5 mL酶液,充分搖勻后于40 ℃恒溫水浴鍋中準確反應60 min,最后加入1 mol/L NaOH溶液作為反應終止劑。隨后加入2 mL DNS試劑置于沸水浴中反應5 min,流水冷卻后在分光光度計測定OD540值。
酶活力單位定義:在pH 7.0、40 ℃條件下,每分鐘催化水解羧甲基纖維素鈉生成1 μmol還原糖所需酶量為1個酶活力單位[23]。
1.2.5 β-1,4-內切葡聚糖酶的最適溫度和溫度穩定性測定
最適溫度測定方法如下:將酶液在不同溫度條件(15、20、25、30、35、40、45、50、55、60 ℃)下測定酶活力,將最高酶活力定義為100%,計算相對酶活力,其中測得酶活力最大值對應的溫度即為β-1,4-內切葡聚糖酶的最適溫度。
溫度穩定性測定方法如下:將酶液置于15、20、25、30、35、40、45、50、55、60 ℃保溫60 min,迅速冰浴冷卻至40 ℃后參考1.2.4節方法測定酶活力,以在相同溫度條件下處理后酶活力與未處理酶活力的比值表征酶的剩余酶活力,以最高酶活力定義為100%計算剩余酶活力。
1.2.6 β-1,4-內切葡聚糖酶的最適pH和pH穩定性測定
最適pH值測定方法如下:分別配制20 mmol/L醋酸/醋酸鈉緩沖液(pH 5.0~6.0)、20 mmol/L Tris-HCl緩沖液(pH 6.5~10.0)和20 mmol/L Gly-NaOH緩沖液(pH 10.0~12.0),并與羧甲基纖維素鈉底物等體積混合;將酶液在相應pH條件下測定酶活力,以最大酶活力為100%計算相對酶活力。
pH穩定性測定方法如下:將酶液置于上述緩沖液中于40 ℃恒溫處理1 h后,在最適溫度下測定酶活力,以相同pH條件下處理后酶活力與未處理酶活力的比值表征酶的剩余酶活力。
1.2.7 金屬離子、表面活性劑和化學試劑對β-1,4-內切葡聚糖酶酶活力的影響
在底物溶液中加入終濃度分別為1 mmol/L和10 mmol/L的金屬離子溶液(K+、Cu2+、Ba2+、Mg2+、Fe3+、Mn2+、Zn2+、Ca2+、Li+、Ni2+、Na+、Co2+)以及終質量濃度10 mg/L的表面活性劑和化學試劑[十二烷基硫酸鈉(sodium dodecyl sulfate, SDS)、十二烷基苯磺酸(dodecyl benzene sulfonic acid, LAS)、聚山梨醇酯-20 (Tween-20)、聚氧乙烯山梨糖醇酐單硬酸酯(Tween-60)、聚山梨酯-80 (Tween-80)、聚乙二醇辛基苯基醚(Triton)、二甲基亞砜(dimethyl sulfoxide, DMSO)、苯甲磺酰氟(phenyl methane sulfonyl fluoride, PMSF)、鹽酸胍(guanidine hydrochloride, GuHCl)、溴甲烷(bromomethane, MB)、丙酮(acetone, AC)、二硫蘇糖醇(dithiothreitol, DTT)、N-亞硝基嗎啉(N-nitrosomorpholine, NNM)、乙二胺四乙酸(ethylene diamine tetraacetic acid, EDTA)、尿素(urea)],再加入β-1,4-內切葡聚糖酶酶液在最適條件下反應60 min后測定酶活力,以未處理測得的酶活力為100%計算相對酶活。
1.2.8 β-1,4-內切葡聚糖酶的底物特異性分析
分別以CMC-Na、微晶纖維素、濾紙和棉等為底物,在最適條件下測定β-1,4-內切葡聚糖酶的酶活力,分析β-1,4-內切葡聚糖酶對不同底物的水解能力。
1.2.8 β-1,4-內切葡聚糖酶的催化動力學參數分析
將CMC-Na底物與20 mmol/L Tris-HCl緩沖液(pH 7.0)混合,配制成質量濃度分別為0.5、1.0、1.5、2、3、4、5、6、7、8、9、10、15、20、25 mg/mL的底物溶液,在最適條件下測定酶活力[24]。通過擬合Lineweaver-Burk雙倒數曲線計算Km和Vmax值,Vmax除以對應的底物濃度即為催化常數kcat值,進一步計算可得催化效率常數kcat/Km值。
1.2.8 β-1,4-內切葡聚糖酶的催化動力學參數分析
將純化后的β-1,4-內切葡聚糖酶液分別置于pH 8.0和pH 9.0環境,并保持溫度為40 ℃。每隔10 min加入羧甲基纖維素鈉底物測定酶活力,直到剩余酶活力達到初始酶活力的50%以下[25]。將β-1,4-內切葡聚糖酶在不同pH條件下酶活力降至一半所需要的時間定義為酶的半衰期。
2 結果與分析
2.1 β-1,4-內切葡聚糖酶序列挖掘
基于獲得耐低溫和耐堿β-1,4-內切葡聚糖酶的目標,通過pfam數據庫篩選得到一株BacillusKSM-635來源的低溫、耐堿β-1,4-內切葡聚糖酶基因(Genbank No.1G01_A);將該酶保守功能域氨基酸在NCBI數據庫進行blastp搜索,共獲得209個潛在內切葡聚糖酶基因序列。所篩選到的β-1,4-內切葡聚糖酶在活性區域具有高度的保守性,為了保證酶基因的新穎性,保留同源性在40%~75%的氨基酸序列,共篩選得到99株潛在氨基酸序列,并構建了系統進化樹。從圖1-a可以看出,基于進化距離以及來源分布,β-1,4-內切葡聚糖酶主要來源于芽孢桿菌屬,同時在梭狀芽孢桿菌、奇異球菌、棲熱袍菌、假單胞菌屬微生物中也鑒定得到部分新型酶基因。
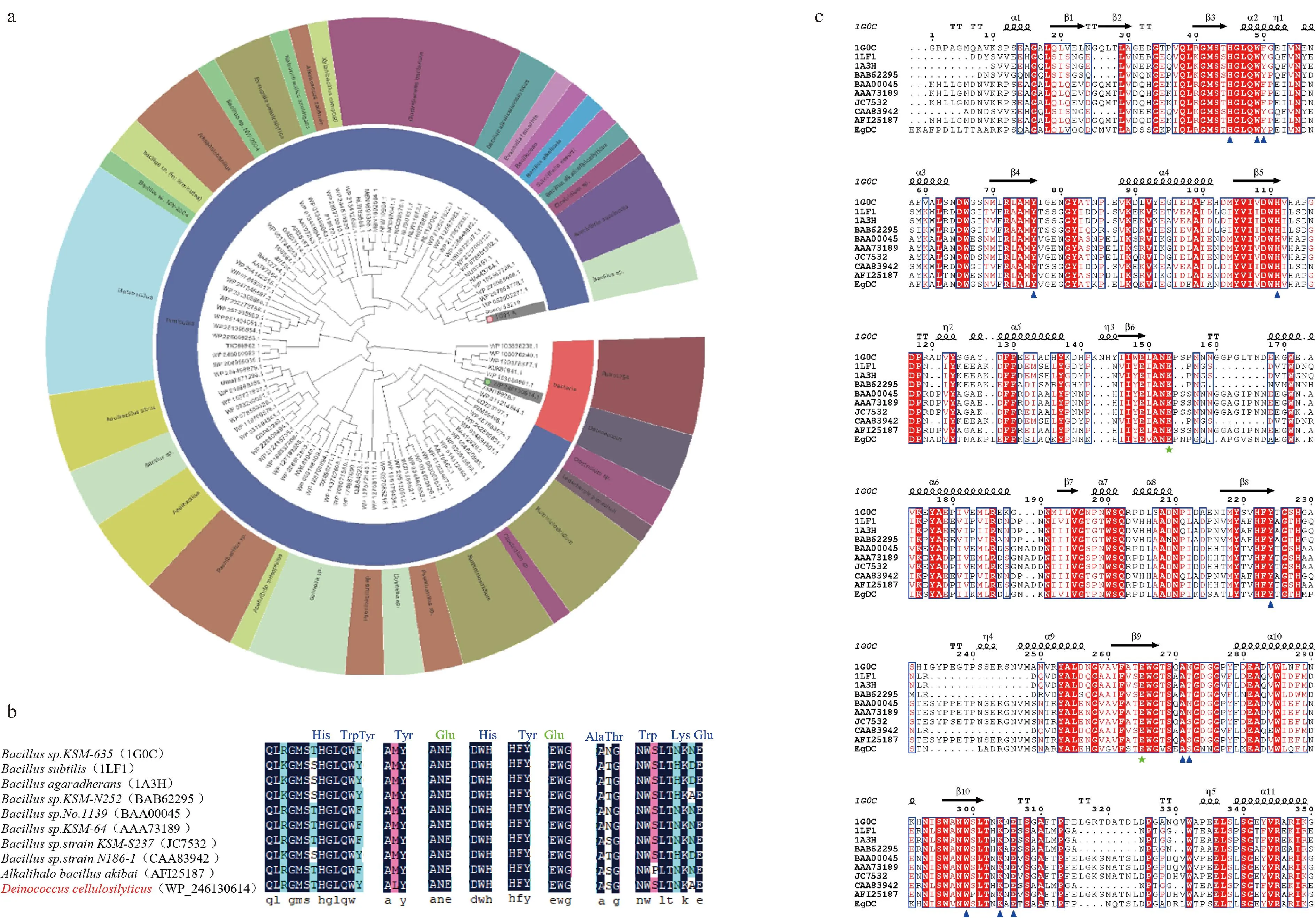
a-挖掘得到酶基因搭建的系統進化樹;b-潛在β-1,4-內切葡聚糖酶的保守氨基酸序列比對;c-WP_246130614編碼蛋白與其同源性較高的蛋白多序列比對結果圖1 β-1,4-內切葡聚糖酶的基因挖掘結果分析
為進一步篩選具有潛在工業應用價值的β-1,4-內切葡聚糖酶基因,首先去除具有組合纖維素酶功能域、缺少CBM結構域等不符合篩選特定保守域的序列;在此基礎上,基于棲熱袍菌、假單胞菌屬菌株特性,其來源β-1,4-內切葡聚糖酶通常不具有低溫、耐堿性的特征,因此也被排除。經過上述2輪篩選,共排除49個酶序列,共余49個酶序列。剩余序列通過分析序列氨基酸占比情況,并計算了各β-1,4-內切葡聚糖酶的理論最適pH值。為保證堿性條件下的應用,篩選理論最適pH值在7.0以上的序列,考慮到在洗滌行業中的低溫環境,分析低溫β-1,4-內切葡聚糖酶的氨基酸序列特點,篩選具有更少的內部作用力、鹽橋數量少和更多非極性殘基暴露的序列[26]。為了確定挖掘得到的β-1,4-內切葡聚糖酶基因的功能,將篩選剩下的內切葡聚糖酶氨基酸序列中的關鍵催化區域與BacillusKSM-635來源內切葡聚糖酶1G0C序列進行氨基酸序列比對。從圖1-b和圖1-c可以看出,篩選得到的β-1,4-內切葡聚糖酶氨基酸序列的催化功能域和底物結合位點(藍色)與已驗證的模板酶序列具有高度保守性,其具有相同的催化活性位點Glu155和Glu259(綠色),這表明篩選得到的內切葡聚糖酶基因具有潛在的β-1,4-內切葡聚糖酶催化能力。考慮到基因序列的可獲得性和操作性,選擇與內切葡聚糖酶1G0C氨基酸序列相似性為60.83%的Deinococcuscellulosilyticus來源內切葡聚糖酶EgDC(Genbank No.WP_246130614)進行下一步研究。
2.2 β-1,4-內切葡聚糖酶EgDC的異源表達及純化
將內切葡聚糖酶EgDC基因通過pET-22b(+)載體轉化進入E.coliBL21,成功搭建重組大腸桿菌。通過含pET-22b(+)-EgDC的重組大腸桿菌的發酵液和全細胞裂解液的SDS-PAGE分析研究了酶的表達情況。如圖2-a所示,重組大腸桿菌的發酵液與全細胞裂解液都在44.3~66.4 kDa均出現了條帶,與酶分子質量理論值58 kDa基本相符。與此同時,在24 h和48 h發酵液中出現了明顯條帶,證明內切葡聚糖酶EgDC成功實現了表達。在此基礎上,基于重組β-1,4-內切葡聚糖酶EgDC蛋白質C端帶有his-tag標簽,通過Ni2+親和層析對其進行純化。通過SDS-PAGE分析,純化后的EgDC顯示單一蛋白質條帶,表觀分子質量為58 kDa,與理論值相符。
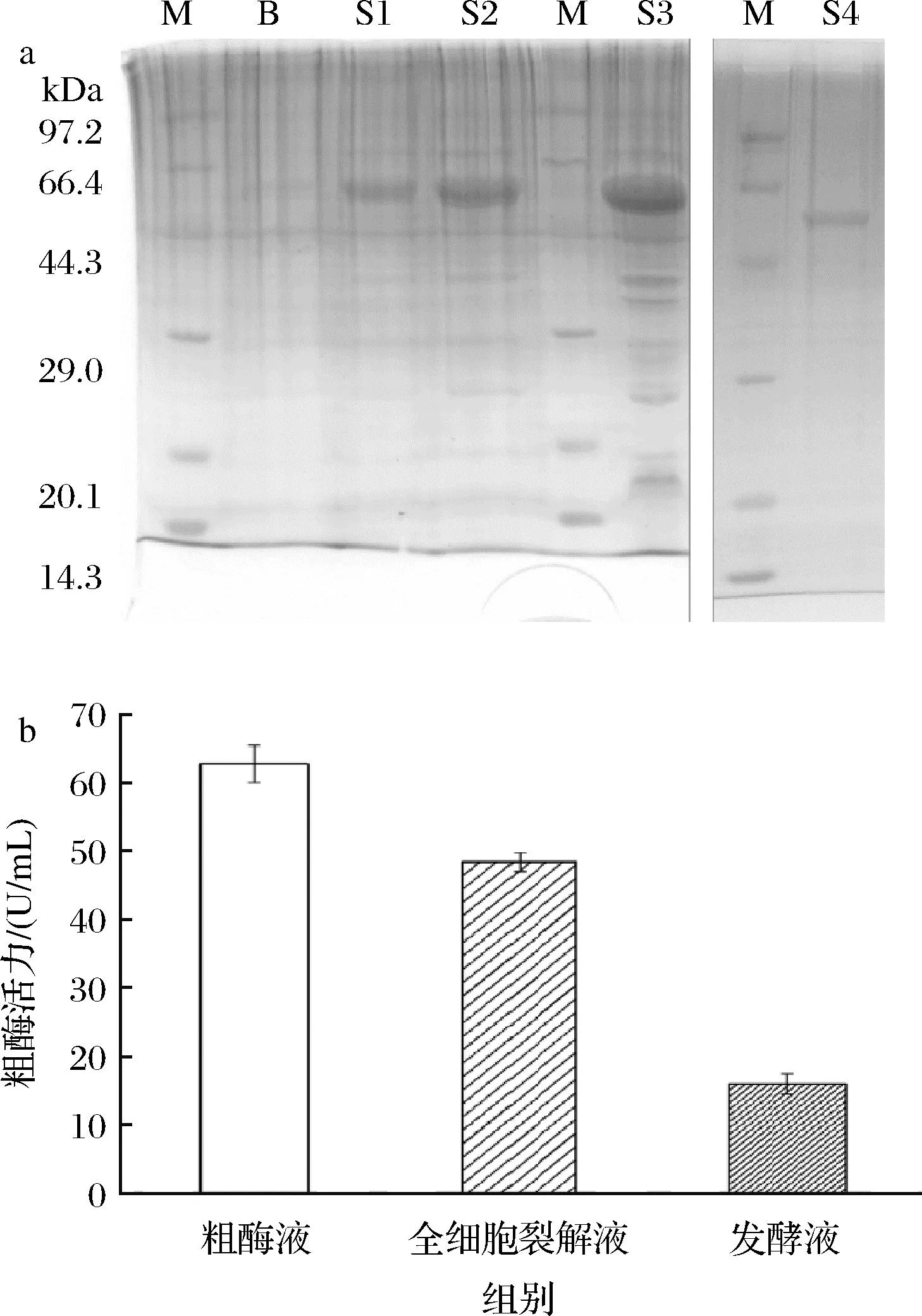
a-重組β-1,4-內切葡聚糖酶發酵液、全細胞裂解液和純化酶的SDS-PAGE分析;b-粗酶活力分布情況圖2 重組β-1,4-內切葡聚糖酶表達結果分析
為了進一步驗證重組大腸桿菌對β-1,4-內切葡聚糖酶EgDC的可溶性表達的程度,測定了含pET-22b(+)-EgDC的重組大腸桿菌誘導40 h后的發酵液、粗酶液和全細胞裂解液的總酶活力。如圖2-b所示,發酵40 h后粗酶液酶活力為(62.25±3.98) U/mL。其中,全細胞裂解液中測得酶活力為(49.74±1.65) U/mL,約占粗酶液酶活力的3/4;此外,少量重組酶被分泌到發酵液中[(16.55±1.01) U/mL]。由此可見,β-1,4-內切葡聚糖酶EgDC絕大部分存在于細胞周質,少部分分泌到胞外。
2.3 β-1,4-內切葡聚糖酶EgDC的酶學性質分析
2.3.1 β-1,4-內切葡聚糖酶的最適溫度和溫度穩定性
測定了β-1,4-內切葡聚糖酶EgDC的最適溫度和溫度穩定性。如圖3-a所示,β-1,4-內切葡聚糖酶EgDC的最適溫度為40 ℃,與之前報道的能夠在洗滌行業進行應用的內切葡聚糖酶的最適溫度(30~45 ℃)相符。但是,EgDC在20~40 ℃時依然保持70%以上的相對酶活力,這有利于其在低溫環境下的應用。在此基礎上,將EgDC在不同溫度下溫育1 h后測定其殘余酶活力。如圖3-b所示,EgDC的剩余酶活力隨著溫度的提高而逐漸降低。當溫度低于45 ℃時,溫育1 h后EgDC仍可以保持60%以上的活性;而當溫度高于50 ℃時,溫育1 h后EgDC的剩余酶活力下降較明顯。在60 ℃處理1 h時后EgDC的剩余酶活力僅剩10%~20%。
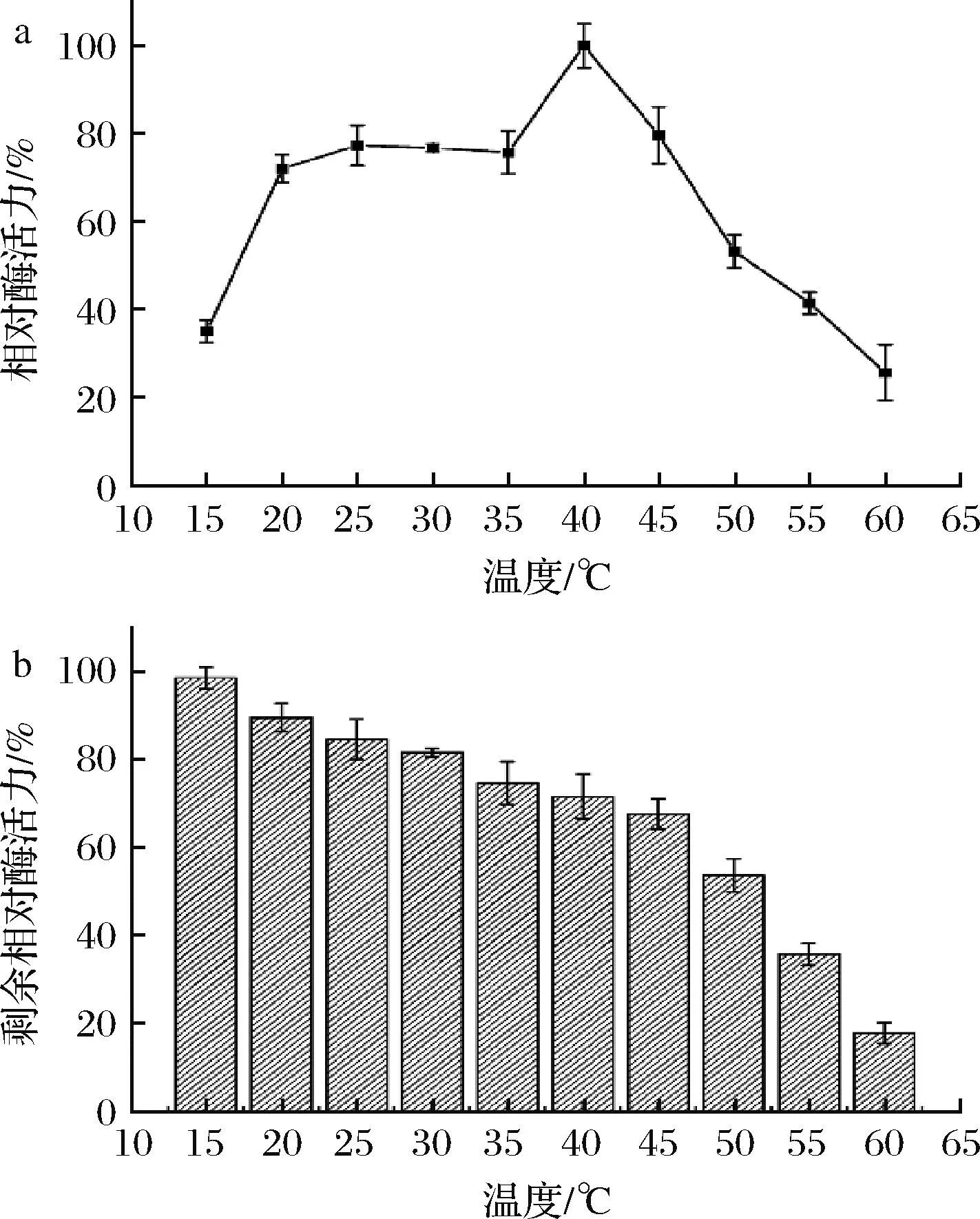
a-β-1,4-內切葡聚糖酶EgDC的最適溫度曲線;b-EgDC的溫度穩定性圖3 β-1,4-內切葡聚糖酶EgDC的溫度酶學特征
2.3.2 β-1,4-內切葡聚糖酶的最適pH和pH穩定性
測定了β-1,4-內切葡聚糖酶EgDC的最適pH值和pH穩定性。如圖4-a所示,EgDC的最適pH值為6.5。在pH 6.0~8.0時,EgDC的相對酶活力能保持90%以上;當pH值>8.5時,EgDC的相對酶活呈現顯著下降趨勢,剩余酶活力低于60%。在EgDC的pH穩定性方面,EgDC在中性條件下的穩定性較好,其在pH 6.5~8處理1 h后依然能保持高于60%的剩余酶活力。在pH值>8.5時,EgDC的穩定性相對較差,其在處理1 h后的剩余酶活力在20%~40%(圖4-b)。
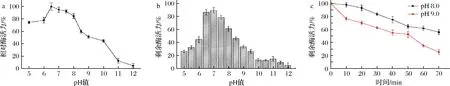
a-β-1,4-內切葡聚糖酶EgDC的最適pH值曲線;b-EgDC的pH穩定性;c-EgDC在pH 8.0和pH 9.0下的半衰期圖4 β-1,4-內切葡聚糖酶EgDC的pH酶學特征
同時測定了β-1,4-內切葡聚糖酶EgDC在pH 8.0和pH 9.0條件下的半衰期。如圖4-c所示,EgDC在pH 8.0條件下經70 min處理后的剩余酶活力依然在55%以上,說明EgDC具有一定程度的耐堿性。EgDC在pH 9.0條件下隨著處理時間延長,剩余酶活力呈現明顯下降趨勢,在50 min左右剩余50%的酶活力。
2.3.3 金屬離子、表面活性劑和化學試劑對β-1,4-內切葡聚糖酶的影響
測定了金屬離子對β-1,4-內切葡聚糖酶EgDC酶活力的影響。如圖5-a所示,1 mmol/L的Mn2+、Ca2+和10 mmol/L的K+、Ba2+、Mg2+、Li+、Na+對β-1,4-內切葡聚糖酶EgDC具有促進作用。其中,10 mmol/L的Ba2+對EgDC酶活力的促進作用最明顯,在其存在條件下酶活力可達初始酶的130%左右。在1 mmol/L時,大部分金屬離子(如K+、Ba2+、Mg2+、Ca2+、Na+)對EgDC酶活力具有一定的抑制作用,而當濃度增加到10 mmol/L時其抑制作用均變弱,甚至具有一定的促進作用。在所有金屬離子中,對EgDC酶活力抑制作用最強的為Cu2+和Zn2+。在含10 mmol/L Cu2+或Zn2+情況下,EgDC的相對酶活力低于初始酶的10%,甚至接近為0。在洗滌條件下,能夠耐受一定濃度的金屬離子可以有效提高β-1,4-內切葡聚糖酶在洗滌中的應用價值。
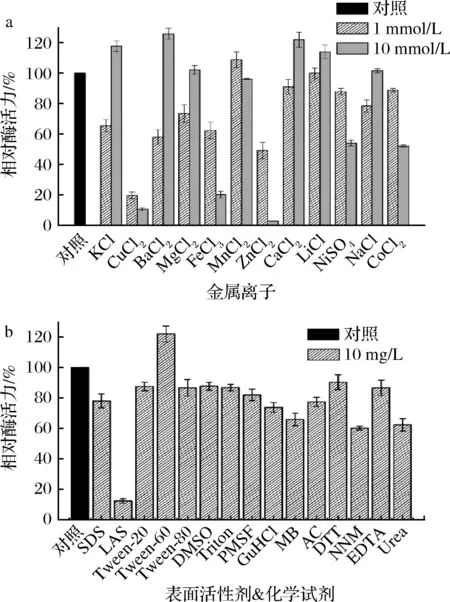
a-金屬離子對β-1,4-內切葡聚糖酶EgDC酶活力的影響;b-表面活性劑及有機試劑對β-1,4-內切葡聚糖酶EgDC酶活力的影響圖5 β-1,4-內切葡聚糖酶EgDC對金屬離子、表面活性劑和有機試劑的耐受性分析
同時測定了化學試劑(包括有機試劑、表面活性劑和抑制劑)對EgDC酶活力的影響。如圖5-b所示,EgDC對大部分化學試劑表現出較好的耐受性,其中Tween-60對EgDC酶活力具有促進作用。在10 mg/L Tween-60條件下,EgDC酶活力達到初始酶的120%以上。盡管陰離子表面活性劑烷基苯磺酸鈉(sodium alkylbenzenesulfonate,LAS)對于EgDC酶活力具有較強的抑制作用,但是Tween-60、Tween-80、TritonX-100這些非離子表面活性劑對酶活力的影響非常小。在10 mg/L陰離子表面活性劑SDS存在情況下,EgDC也具有初始酶75%以上的活性,這說明β-1,4-內切葡聚糖酶EgDC具有一定的適應洗滌環境的能力,能夠與表面活性劑進行復配使用;此外,有機試劑(如甲苯、丙酮、二甲亞砜、尿素)對酶活力的影響不是很顯著;EDTA、PMSF和鹽酸胍等抑制劑對EgDC酶活力沒有較大的影響,說明EgDC催化關鍵區域無需絲氨酸或金屬離子。
2.3.4 β-1,4-內切葡聚糖酶的底物特異性分析
β-1,4-內切葡聚糖酶EgDC的底物特異性分析結果如表1所示。EgDC對可溶性底物羧甲基纖維素鈉的降解活性最高,達到(26.38±1.43) U/mg;此外,其對于微晶纖維素也有一定的活性[(12.31±1.23) U/mg]。EgDC在以濾紙或棉作為底物時的酶活力較低。這可能是由于EgDC結構中存在碳水化合物結合位點(carbohydrate binding module, CBM),對催化底物存在一定選擇性。

表1 β-1,4-內切葡聚糖酶EgDC的底物特異性Table 1 Substrate specificity of β-1,4-endoglucanase EgDC
2.3.5 β-1,4-內切葡聚糖酶的酶促動力學和半衰期
以羧甲基纖維素鈉為底物測定了β-1,4-內切葡聚糖酶EgDC的比酶活力和催化動力學參數。如表2所示,通過擬合Michaelis-Menten方程計算,EgDC在最適條件下的Km值和Vmax值分別為(0.043±0.000 5) mg/mL和(29.67±0.065) U/(mg·s),其kcat和kcat/Km的值分別為(22.14±0.49) s-1和(520.03±17.07) s-1/mg(表2)。與其他研究報道[14]相比,EgDC的Km值更低,而其kcat/Km值,說明EgDC對羧甲基纖維素鈉具有更高的親和力以及更好的的催化性能,說明該酶在洗滌行業具有潛在應用價值。
3 結論
本文通過基因挖掘獲得了一株D.cellulosilyticus來源的新低溫β-1,4-內切葡聚糖酶基因EgDC,并在大腸桿菌中實現了可溶性表達。EgDC的比酶活力為(26.38±1.43) U/mg,其最適溫度和最適pH值分別為40 ℃和6.5,且在低溫和中性環境下具有較好的穩定性。該酶對金屬離子、化學試劑以及表面活性劑均具有較強的耐受性。與其他報道[14,27]相比,EgDC具有較更高的kcat/Km值,說明具有更佳的催化性能。上述結果說明,這種新型低溫β-1,4-內切葡聚糖酶具有洗滌行業所需要的大部分性質,具有潛在的工業應用價值。近年來研究者對于生存環境比較特殊的極端微生物的研究較多,這也成為潛在的工業酶資源挖掘的主要方向,但是這種方法存在菌株篩選、目的酶基因難以獲得,極端微生物難以培養以及較低的開發效率等缺陷。該研究基于已有的序列進行生物信息學分析,實現通過基因挖掘獲得特異的酶基因,為以后針對不同行業的特殊需求挖掘新型工業酶提供了可選途徑。
