河北省廊坊市牛主要病毒性腹瀉病原感染狀況檢測(cè)及牛冠狀病毒演化分析
2024-03-01 12:34:56李思遠(yuǎn)付新成袁雪松蔡旭航孫心如謝玲玲李基棕
畜牧獸醫(yī)學(xué)報(bào) 2024年2期
李思遠(yuǎn),付新成,袁雪松,毛 立,蔡旭航,孫心如,黃 金,謝玲玲,王 府,周 華,張 琪,李基棕,4*,李 彬,4*
(1.西北農(nóng)林科技大學(xué)動(dòng)物醫(yī)學(xué)院,楊凌 712100;2.江蘇省農(nóng)業(yè)科學(xué)院獸醫(yī)研究所,農(nóng)業(yè)農(nóng)村部獸用生物制品工程技術(shù)重點(diǎn)實(shí)驗(yàn)室,南京 210014;3.南京農(nóng)業(yè)大學(xué)動(dòng)物醫(yī)學(xué)院,南京 210095;4.江蘇大學(xué)生命科學(xué)學(xué)院 食品與生物工程學(xué)院,鎮(zhèn)江 212013;5.河北省廊坊市農(nóng)業(yè)農(nóng)村局,廊坊 065000;6.貴州省種畜禽種質(zhì)測(cè)定中心,貴陽(yáng) 550018;7.黔西市動(dòng)物疫病預(yù)防控制中心,黔西 551500)
牛冠狀病毒(bovine coronavirus, BCoV)屬于β冠狀病毒,在全球范圍廣泛流行。主要編碼5個(gè)結(jié)構(gòu)蛋白,血凝素酯酶(hemagglutinin-esterase,HE)、刺突蛋白(spike protein,S)、包膜蛋白(envelope protein,E)、膜蛋白(membrane,M)和核衣殼蛋白(nucleocapsid,N)。BCoV最初被認(rèn)為是腸道病毒,可以造成新生犢牛腹瀉[1-2]、牛冬季痢疾,但迄今為止,越來(lái)越多的臨床案例表明該病毒可以從呼吸道檢測(cè)出并導(dǎo)致各年齡段牛呼吸道癥狀[3-4],因此BCoV也是呼吸道病原的說(shuō)法越來(lái)越頻繁地被提及。BCoV具有很大的跨物種傳播潛力,CoV-OC43被認(rèn)為與BCoV相近,這兩種病毒均能以9-O乙酰唾液酸作為糖結(jié)合底物[5-6],受人白細(xì)胞抗原Ⅰ亞群(human leukocyte antigen,HLA-Ⅰ)影響[5,7-8],所以BCoV常被作于人呼吸道冠狀病毒(人冠狀病毒OC-43、SARS-CoV2、HKU1)的研究模型。除了感染牛以外,在其他動(dòng)物(如羊駝、羊、長(zhǎng)頸鹿等)中也檢出類BCoV,其中更包括了德國(guó)的人呼吸道株4403[9-12],具有溢出跨宿主傳播風(fēng)險(xiǎn)。
牛病毒性腹瀉/黏膜病病毒(bovine viral diarrhea virus, BVDV)是全世界牛常見(jiàn)的疾病之一。BVDV可以導(dǎo)致牛生育能力下降、產(chǎn)奶量下降、呼吸道癥狀、腹瀉、流產(chǎn)和產(chǎn)死胎等生殖功能異常[13-14]。在亞洲不同國(guó)家報(bào)道中,牛星狀病毒(bovine astroviruses, BoAstV)被證明與牛腹瀉有關(guān),但報(bào)道的發(fā)病率不一致[15-16]。牛輪狀病毒(bovine rotavirus virus, BRV)是牛腹瀉最常見(jiàn)的原因,能導(dǎo)致顯著的并發(fā)癥和死亡率,在新生犢牛中尤為明顯[17-18]。牛環(huán)曲病毒(bovine torovirus, BToV)主要引起小牛和成年牛腹瀉,還具有雙重組織嗜性,可以在呼吸道中檢出,并導(dǎo)致牛腹瀉,嚴(yán)重患病牛可至死亡[19-21]。牛諾瓦病毒(bovine norovirus, BoNoV)是引起動(dòng)物胃腸炎的重要病原[22]。嵌杯病毒科的牛紐布病毒(bovine nebovirus, BoNeV)可導(dǎo)致患病牛小腸絨毛萎縮,引起腹瀉[22-23]。哺乳動(dòng)物正呼腸孤病毒(mammalian orthoreovirus, MRV)和牛嵴病毒(bovine kobuvirus, BKoV)也被認(rèn)為是引起牛腹瀉的病原之一[24-25]。
本研究對(duì)我國(guó)河北省廊坊市以BCoV為主的腹瀉病原進(jìn)行流行病學(xué)調(diào)查,對(duì)BCoV遺傳進(jìn)化分析,為該地這9種病的發(fā)病、流行情況及疫病防控提供科學(xué)數(shù)據(jù)。
1 材料與方法
1.1 樣品
從我國(guó)河北廊坊地區(qū)14個(gè)養(yǎng)殖場(chǎng)中采集了323份腹瀉奶牛糞樣。
1.2 主要試劑
RNA提取試劑盒、逆轉(zhuǎn)錄酶、Green Taq Mix、2×Phanta Max Master Mix (Dye Plus) 均購(gòu)自南京諾唯贊生物科技有限公司,瓊脂糖購(gòu)自北京擎科生物科技有限公司,膠回收試劑盒購(gòu)自O(shè)mega Bio-Tek公司,磷酸鹽緩沖液(PBS)購(gòu)自塞維爾生物科技有限公司。
1.3 主要儀器
PCR擴(kuò)增儀和離心機(jī):德國(guó)Eppendorf公司;凝膠成像系統(tǒng):上海天能公司。
1.4 樣品處理
將采集的糞樣每份轉(zhuǎn)移部分至2 mL離心管中,其余凍存至-80 ℃冰箱。每個(gè)EP管中加入1 mL 經(jīng)高壓滅菌的PBS溶液進(jìn)行震蕩充分混勻,離心后棄去沉淀取上清用0.22 mm濾器過(guò)濾后流液備用。
1.5 引物合成
參考文獻(xiàn)[26-32]合成BCoV、BVDV、BoAstV、BRV、BToV、BNoV、BoNeV、MRV、BKoV相關(guān)片段的引物(表1、2)。

表1 病毒性腹瀉病原檢測(cè)引物信息

表2 BCoV S和HE引物信息
1.6 病毒RNA提取及轉(zhuǎn)錄
參照說(shuō)明進(jìn)行提取總RNA后進(jìn)行反轉(zhuǎn)錄,反轉(zhuǎn)錄產(chǎn)物cDNA凍存于-80 ℃冰箱備用。
1.7 RT-PCR擴(kuò)增
根據(jù)表1檢測(cè)引物,采用20 μL體系:10 μL Mix,0.4 μL 上游引物F,0.4 μL 下游引物R,1 μLcDNA,8.2 μL ddH2O。使用降落PCR方法擴(kuò)增片段,反應(yīng)程序:預(yù)變性95 ℃ 5 min;95 ℃ 15 s,48~55 ℃ 15 s,72 ℃ 30 s,共35個(gè)循環(huán);徹底延伸72 ℃ 10 min。PCR產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳,觀察結(jié)果。
根據(jù)S和HE特異性引物擴(kuò)增BCoV陽(yáng)性樣品,采用25 μL體系:12.5 μL Mix,1 μL 上游引物F,1 μL 下游引物R,2 μL cDNA,8.5 μL ddH2O。使用降落PCR方法擴(kuò)增目的片段,程序?yàn)?預(yù)變性95 ℃ 5 min;95 ℃ 15 s,48~55 ℃ 15 s,72 ℃ 1 min 共35個(gè)循環(huán);徹底延伸72 ℃ 10 min。得到產(chǎn)物使用1%瓊脂糖凝膠電泳,觀察結(jié)果。
1.8 膠回收驗(yàn)證及進(jìn)化樹(shù)構(gòu)建
按膠回收試劑盒說(shuō)明書(shū)回收目的條帶膠塊,由上海生工生物工程股份有限公司測(cè)序,通過(guò)測(cè)序結(jié)果進(jìn)行序列比對(duì)。同時(shí)將提取的RNA送上海探普生物公司進(jìn)行病毒基因組下一代測(cè)序。
使用MEGA 11軟件進(jìn)行BCoV的HE、S、N、M、E、ORF1a和全序列基因核酸序列比對(duì),并使用最大似然法構(gòu)建進(jìn)化樹(shù)。
2 結(jié) 果
2.1 RT-PCR檢測(cè)
在檢測(cè)323份樣品中BCoV總體陽(yáng)性檢出率為14.24% (46/323),BVDV為1.85% (6/323),BRV為57.89% (187/323),BoAstV為0.31% (1/323),BoNeV為10.84% (35/323),BoNoV為4.33% (14/323),MRV為2.48% (8/323),BToV為4.64% (15/323),BKoV為11.76% (38/323)。該地區(qū)BCoV檢出率并不高,而B(niǎo)RV高達(dá)57.89%,檢出率最低的是BoAstV,僅有0.31%。各個(gè)牛場(chǎng)不同病原陽(yáng)性檢出率差異較大,BVDV檢出率為0.00%~17.65%,BRV檢出率為0.00%~100%,BCoV 檢出率為0.00%~45.45%,BKoV檢出率為0.00%~42.47%,BNeV檢出率為0.00%~62.50%,BNoV檢出率為0.00%~30.00%,BToV檢出率為0.00%~13.33%,MRV檢出率為0.00%~11.76%,BoAstV檢出率為0.00%~3.33%。其中BRV在每個(gè)牛場(chǎng)中均能檢出,在7個(gè)奶牛場(chǎng)中檢出率為100.00%。而B(niǎo)CoV也在12個(gè)奶牛場(chǎng)中檢出,僅次于BRV。
2.2 混合感染檢測(cè)
總共有22種混合感染類型,最多同時(shí)感染5種病毒。以BRV和BoNeV混合感染檢出率最高,高達(dá)28.95% (22/76),BCoV混合感染比率為 36.84%(28/76) (表3)。

表3 混合感染狀況分析結(jié)果
2.3 BCoV的HE和S基因序列擴(kuò)增
陽(yáng)性樣本采用特異性引物擴(kuò)增HE和S片段,可分段擴(kuò)增出1株HE和S全長(zhǎng)序列,命名為HBLF2302株(圖1)。

M.DL2000 DNA 相對(duì)分子質(zhì)量標(biāo)準(zhǔn); 1. HBLF2302檢測(cè)片段;2. HA;3. HB;4. HE;5. S1;6. 35;7. 3536;8. 36;9. 3637;10. S2;11. S3;12. DL5000 DNA 相對(duì)分子質(zhì)量標(biāo)準(zhǔn)M.DL2000 DNA maker; 1. HBLF2302 detected segment;2. HA;3. HB;4. HE;5. S1;6. 35;7. 3536;8. 36;9. 3637;10. S2;11. S3;12. DL5000 DNA maker
2.4 BCoV結(jié)構(gòu)蛋白遺傳進(jìn)化分析
根據(jù)NCBI數(shù)據(jù)庫(kù)及DNAstar 7.0中的Megalign比對(duì),HBLF2302S基因與我國(guó)華東地區(qū)MZ711 357.1株相似性最高,達(dá)到99.5%。HBLF2302HE基因,與我國(guó)西南地區(qū)獲得的MN982 191.1序列相似性最高,可達(dá)99.53%,序列中無(wú)12個(gè)堿基插入。HBLF2302E、M、N基因與我國(guó)其它地區(qū)測(cè)得的序列相似性均在99.7%以上,差異較小。
從NCBI數(shù)據(jù)庫(kù)獲得的部分序列S序列進(jìn)行比對(duì),S進(jìn)化樹(shù)結(jié)果表明HBLF2302屬于GⅡb基因型,為國(guó)內(nèi)BCoV流行的基因型,值得注意的是,HY24株為我國(guó)牦牛源毒株,p95為我國(guó)臺(tái)灣省于HRT-18細(xì)胞上測(cè)得,屬于GⅠa基因型。不僅如此BCoV-GX-NN190313和BJ232犬冠狀病毒處于獨(dú)立分支,提示國(guó)內(nèi)BCoV可能存在新基因型。韓國(guó)流行株主要以GⅡa基因型為主,但也出現(xiàn)有GⅡb基因型。表明不同基因型跨地區(qū)傳播的風(fēng)險(xiǎn)在增加。可能存在新基因型的出現(xiàn),有待進(jìn)一步測(cè)序驗(yàn)證。HE、E、M和N基因在不同基因型中未見(jiàn)有明顯分化,表明在不同基因型中除了S差異較大具有分型價(jià)值外,其余四個(gè)結(jié)構(gòu)蛋白可能不可作為分型依據(jù)。
2.5 BCoV S蛋白三級(jí)結(jié)構(gòu)分析
通過(guò)SWISS-MODEL同源建模及PyMDL分析,發(fā)現(xiàn)氨基酸157號(hào)區(qū)別于其他參考序列157V和157I,854I較為少見(jiàn),318號(hào)位為獨(dú)特突變位點(diǎn),在HBLF2302株中處于選擇。其中157 T和318V位于S1,854I位于S2。318號(hào)位點(diǎn)在HBLF2302上由318I突變?yōu)?18V時(shí),與631C形成氫鍵(圖2)。

A. 318I局部結(jié)構(gòu);B. 318V局部結(jié)構(gòu)A. Local structure of 318I of S; B. Local stucture of 318V of S
2.6 BCoV ORF1a序列及分析
通過(guò)氨基酸比對(duì)結(jié)果表明,在目前上傳至NCBI中的BCoV中國(guó)株的開(kāi)放閱讀框1a(Open reading frame ORF1a)基因中834、857和889號(hào)位為區(qū)別于其它地區(qū)的GⅡb株獨(dú)特的位點(diǎn)。在其他序列中834W、857 L和889R號(hào)位均為一致,而在中國(guó)株中834W為834R,857 L為857 S,而在889R為終止密碼子(圖3)。從遺傳進(jìn)化樹(shù)上可以看出,除了牦牛源MH810 163.1 HY24仍與GⅠa型傳統(tǒng)株處于同一分支外,中國(guó)株獨(dú)立于同為GⅡb型的日本株和美洲株獨(dú)立成簇(圖4)。由此可見(jiàn),在ORF1a上,同一基因型的BCoV毒株存在一定差異性,并且中國(guó)株呈現(xiàn)出獨(dú)特特征。

圖3 ORF1a 834、857和889號(hào)位氨基酸序列比對(duì)結(jié)果Fig.3 Amino acid sequence alignment of ORF1a 834, 857 and 889

圖4 基于BCoV ORF1a的進(jìn)化樹(shù)分析Fig.4 Phylogenetic tree analysis based on ORF1a of BCoV
2.7 BCoV 全基因遺傳進(jìn)化分析
根據(jù)同源性分析結(jié)果,HBLF2302與從內(nèi)蒙古地區(qū)獲得的序列OP924 545.1相似性最高,在進(jìn)化樹(shù)上也與OP924 545.1進(jìn)化距離接近。全基因組序列FJ938 067.1 4408株與基于S基因的進(jìn)化樹(shù)具有差異,在基于S基因進(jìn)化樹(shù)中與GⅠb基因型歸為一簇,而在全基因組序列中與GⅠa基因型歸為一簇(圖5)。
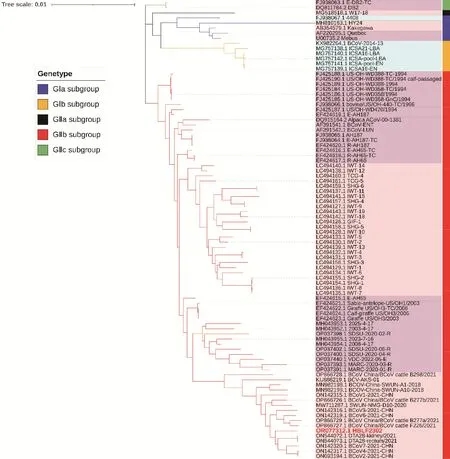
圖5 基于全基因組的BCoV遺傳演化分析Fig.5 Genetic evolution analysis of BCoV based on whole genome
3 討 論
牛群腹瀉的病原多樣,呈現(xiàn)出多重病原混合感染的復(fù)雜情況,在臨床中難以證實(shí)某種病原與腹瀉的直接相關(guān)性。本研究表明,廊坊地區(qū)BCoV檢出率為14.24%,其他9種腹瀉病原的檢出率為0.52%~57.89%。混合感染情況占35.37%。近年來(lái),BCoV流行率有著上升趨勢(shì),不同地區(qū)流行率各有差異,可能存在地方性流行的特點(diǎn)[33-34]。在吉林部分地區(qū)報(bào)道牛冠狀病毒陽(yáng)性檢出率為21.10%[31],云南部分地區(qū)為14.29%[32],在北疆地區(qū)檢出率為0%~38.89%[33],在四川部分地區(qū)為16.80%[34],而在高原地區(qū),腹瀉牛樣本檢出率高達(dá)69.05%,患呼吸道疾病的牛群中檢出率為72.45%[35]。由此可見(jiàn),在不同地區(qū)不同樣本牛冠狀病毒發(fā)病率存在差異,且在腹瀉牛群中檢出率較高。不僅如此,在國(guó)外報(bào)道中,巴西為68.7%[36],日本為57.7%[37],韓國(guó)為39.8%[38],土耳其為21.9%[39],伊朗為7.2%[40],在不同國(guó)家腹瀉牛群中檢出率也有所差異。本文中均為腹瀉樣本,牛冠狀病毒的檢出率為14.24%,可能是由于存在多種病原微生物混合感染,非單一因素或病原引起的牛群腹瀉或存在地區(qū)流行趨勢(shì),因此無(wú)法確切表明,牛冠狀病毒與腹瀉具有直接關(guān)聯(lián)。
按照目前BCoV的基因型劃分,不同地區(qū)存在著不同基因型的BCoV,美洲、東南亞和東亞流行株(除韓國(guó)外)主要為GⅡb,韓國(guó)主要為GⅡa,歐洲主要為GⅠb。此外還存少數(shù)以經(jīng)典株為主的為GⅠa和GⅡc基因型。但由于對(duì)牛冠狀病毒基因型劃分依據(jù)仍不統(tǒng)一,按目前劃分S基因與全基因序列存在差異。更值得注意的是BCoV全基因組序列差異和變異較小,不同地區(qū)流行的BCoV基因型主要可能是由貿(mào)易原因?yàn)橹鲗?dǎo),因此即使在地處亞洲的土耳其,在S基因上趨向于歐洲的GⅠb基因型。
不僅如此,關(guān)鍵的蛋白的突變和重組可能導(dǎo)致BCoV的易感性增加。S中的S1具有雙受體結(jié)合基序[37],S1的N端結(jié)構(gòu)域與9-O乙酰唾液酸結(jié)合[6],S1的受體結(jié)合結(jié)構(gòu)域(receptor-binding domain,RBD)被認(rèn)為與受體結(jié)合,并且大量選擇位點(diǎn)位于RBD處,可能改變其宿主譜進(jìn)而導(dǎo)致溢出。本研究獲取S基因的157號(hào)位點(diǎn)發(fā)生突變,該突變區(qū)別于157V和157I,318V為獨(dú)特突變位點(diǎn),位于S的NTD端,318號(hào)位點(diǎn)的突變改變了側(cè)鏈殘基間相互作用力,是否改變糖嗜性仍需進(jìn)一步研究。除此之外,本文通過(guò)序列比對(duì),發(fā)現(xiàn)在參考序列的中國(guó)株及HBLF2302株中的ORF1a上的834、857及889 aa存在獨(dú)特突變位點(diǎn),區(qū)別于其他地區(qū)的GⅡb基因型毒株并在889號(hào)位點(diǎn)出現(xiàn)終止密碼子,是否在后續(xù)報(bào)道毒株中存在普遍性用于更加細(xì)化解析牛冠狀病毒,仍需要大量測(cè)得序列來(lái)驗(yàn)證,并進(jìn)一步分析。
本研究對(duì)中國(guó)河北地區(qū)牛冠狀病毒及其他病毒性腹瀉病原進(jìn)行了流行病學(xué)調(diào)查,豐富了牛冠狀病毒及其他常見(jiàn)病毒性腹瀉病原的流調(diào)數(shù)據(jù),為進(jìn)一步研究牛冠狀病毒及牛腹瀉性病原提供了數(shù)據(jù)。
4 結(jié) 論
廊坊地區(qū)牛冠狀病毒檢出率為14.24%與其他不同地區(qū)存在差異,檢出率較低。測(cè)得的HBLF2302屬于GⅡb亞型流行株。在ORF1a發(fā)現(xiàn)區(qū)別于中國(guó)以外地區(qū)GⅡb型獨(dú)特的位點(diǎn),并在S蛋白NTD端上有獨(dú)特突變位點(diǎn)。
