干預(yù)自噬調(diào)控p62-Keap1/Nrf2-GPX4通路對結(jié)直腸癌細(xì)胞鐵死亡及奧沙利鉑耐藥的影響
2024-03-07 07:43:16王苗苗張睿哲溫菲菲許曉陽吳淑華
臨床與實(shí)驗(yàn)病理學(xué)雜志 2024年2期
徐 磊,武 寒,王苗苗,張睿哲,溫菲菲,許曉陽,吳淑華
結(jié)直腸癌是全球第三大常見癌癥,也是癌癥相關(guān)死亡的第二大原因[1]。術(shù)后復(fù)發(fā)和轉(zhuǎn)移是結(jié)直腸癌患者死亡的重要原因,化療依然是術(shù)后重要的輔助治療。奧沙利鉑(OXA)是治療結(jié)直腸癌的一線化療藥物[2]。因腫瘤具有耐藥性和長期使用產(chǎn)生不良反應(yīng),導(dǎo)致不足40%的晚期結(jié)直腸癌患者受益于OXA[3]。因此,探究結(jié)直腸癌OXA耐藥的分子機(jī)制對改善OXA對晚期結(jié)直腸癌患者的預(yù)后具有重要意義。
近年研究發(fā)現(xiàn),鐵死亡與腫瘤耐藥密切相關(guān),鐵死亡是一種依賴于細(xì)胞內(nèi)鐵積累和脂質(zhì)過氧化的細(xì)胞程序性死亡形式[4]。谷胱甘肽過氧化物酶4(glutathione peroxidase 4, GPX4)是鐵死亡的重要調(diào)節(jié)因子,干預(yù)以GPX4為代表的脂質(zhì)過氧化物酶途徑促使腫瘤細(xì)胞發(fā)生鐵死亡可能成為逆轉(zhuǎn)腫瘤耐藥的新思路[5]。研究表明,GPX4表達(dá)與自噬適配器p62、Kelch樣Ech相關(guān)蛋白1(Kelchike Ech-associated protein 1, Keap1)、核轉(zhuǎn)錄因子E2相關(guān)因子2(nuclear factor erythroid 2-related factor 2, Nrf2)構(gòu)成的p62-Keap1/Nrf2細(xì)胞氧化應(yīng)激信號通路密切相關(guān)[6]。p62作為重要的選擇性自噬接頭蛋白,是重要的自噬激活標(biāo)志物。自噬被認(rèn)為是維持細(xì)胞穩(wěn)態(tài)的關(guān)鍵過程,LC3是公認(rèn)的自噬標(biāo)志物[7]。目前,越來越多的研究表明自噬參與了鐵死亡調(diào)節(jié)[8]。本課題組前期實(shí)驗(yàn)結(jié)果顯示,結(jié)直腸癌中存在鐵死亡現(xiàn)象,其與Keap1/Nrf2-GPX4表達(dá)有關(guān),且與患者預(yù)后密切相關(guān)[9]。本實(shí)驗(yàn)通過細(xì)胞培養(yǎng)、藥物干預(yù),觀察干預(yù)結(jié)直腸癌細(xì)胞自噬對p62-Keap1/Nrf2-GPX4-鐵死亡及耐藥的影響,探討自噬調(diào)控鐵死亡抑制腫瘤細(xì)胞增殖、侵襲和逆轉(zhuǎn)OXA耐藥性的可行性及其分子機(jī)制,為抑制結(jié)直腸癌的發(fā)生、發(fā)展并逆轉(zhuǎn)耐藥提供新思路。
1 材料與方法
1.1 材料
1.1.1細(xì)胞系 人結(jié)腸癌細(xì)胞系HCT-8、COLO205、SW480、SW620、HCT-116及OXA耐藥細(xì)胞株HCT-116/OXA均購自中國科學(xué)院典型培養(yǎng)物保藏委員會細(xì)胞庫,胎牛血清和RPMI-1640培養(yǎng)基購自美國Gibco公司。
1.1.2試劑 β-actin、LC3、p62、Keap1、Nrf2抗體均購自美國Abcam公司;GPX4抗體購自Affinity公司;辣根過氧化物酶標(biāo)記的羊抗兔多克隆IgG二抗購自福州邁新公司;細(xì)胞增殖及細(xì)胞計(jì)數(shù)試劑盒-8(cell counting kit-8, CCK-8)購自日本同仁化學(xué)研究所;OXA、雷帕霉素(Rapa)、氯喹(CQ)、Erastin、Fer-1和FerroOrange熒光探針均購自GlpBio公司;細(xì)胞內(nèi)自噬染色測定試劑盒[單丹磺酰尸胺(dansylcadaverine, MDC)法]、Western blot凝膠試劑盒、RIPA細(xì)胞裂解液等試劑盒和丙二醛(Malonic dialdehyde, MDA)、還原型谷胱甘肽(glutathione, GSH)檢測試劑盒均購自上海碧云天公司,鐵離子檢測試劑盒購自Solarbio公司;吉姆薩染液購自北京索萊寶公司;Transwell雙層細(xì)胞培養(yǎng)板購自美國Corning公司。
1.2 方法
1.2.1細(xì)胞培養(yǎng) HCT-8、COLO205、SW480、SW620、HCT-116和耐藥細(xì)胞株HCT-116/OXA接種于含5%胎牛血清的RPMI-1640培養(yǎng)基中,置于37 ℃ 5%CO2培養(yǎng)箱中培養(yǎng)。OXA耐藥的HCT-116/OXA細(xì)胞在含5 μmol/L OXA的培養(yǎng)基中持續(xù)培養(yǎng)1周維持抗性。每隔2天換液1次,每3天用0.25%胰蛋白酶消化傳代,取對數(shù)生長期的細(xì)胞進(jìn)行后續(xù)實(shí)驗(yàn)。
1.2.2Western blot法 收集細(xì)胞提取蛋白,采用BCA法測定蛋白濃度。各組取30 μg蛋白進(jìn)行凝膠電泳分離,并將其轉(zhuǎn)印至PVDF膜上,經(jīng)10%脫脂奶粉封閉2 h。一抗4 ℃孵育過夜,二抗室溫孵育1 h,滴加ECL發(fā)光液,成像系統(tǒng)曝光。利用Image J軟件分析目標(biāo)圖像。
1.2.3自噬和鐵死亡干預(yù)實(shí)驗(yàn) 選取HCT-116/OXA耐藥細(xì)胞株,分為自噬激活劑Rapa和自噬抑制劑CQ干預(yù)組,又根據(jù)對鐵死亡的干預(yù)分為不同亞組:Rapa組、Fer-1組、Rapa+Fer-1組、CQ組、Erastin組、CQ+Erastin組及相應(yīng)的對照組。根據(jù)預(yù)實(shí)驗(yàn),設(shè)誘導(dǎo)自噬及鐵死亡改變的最低藥物濃度為干預(yù)劑濃度,在各組細(xì)胞系中分別加入Rapa(0.10 μmol/L)、CQ(10.00 μmol/L)、Erastin(10.00 μmol/L)、Fer-1(1.00 μmol/L)和完全培養(yǎng)基各200 μL,培養(yǎng)24 h。確定干預(yù)成功后,用于后續(xù)實(shí)驗(yàn)。
1.2.4MDC染色 應(yīng)用MDC染色檢測細(xì)胞系中自噬體的形成。制備細(xì)胞懸液,以每孔2×105個細(xì)胞接種于6孔板內(nèi),藥物干預(yù)后吸棄培養(yǎng)液,向每孔中加入1 mL MDC染液,溫箱孵育30 min后置于熒光顯微鏡下觀察,高倍鏡下隨機(jī)選取5個視野,計(jì)數(shù)自噬激活細(xì)胞數(shù)所占百分比,取其平均值。
1.2.5Fe2+、MDA和GSH含量測定 按照試劑盒說明書,使用相應(yīng)試劑盒測定結(jié)直腸癌細(xì)胞中Fe2+、MDA和GSH的含量。酶標(biāo)儀檢測各指標(biāo)吸光度值,并計(jì)算其在細(xì)胞內(nèi)的含量。
1.2.6FerroOrange熒光探針檢測活細(xì)胞內(nèi)Fe2+含量 將HCT-116/OXA細(xì)胞計(jì)數(shù)后以每孔2×105個細(xì)胞接種于6孔板,藥物干預(yù)后吸棄培養(yǎng)液,向每孔加入500 μL經(jīng)無血清培養(yǎng)稀釋的FerroOrange工作液(1 μmol/L),置于培養(yǎng)箱中孵育30 min。熒光顯微鏡下觀察各組細(xì)胞的熒光強(qiáng)度。
1.2.7CCK-8實(shí)驗(yàn) 應(yīng)用CCK-8實(shí)驗(yàn)對細(xì)胞系進(jìn)行OXA藥物毒性測試。制備細(xì)胞懸液,設(shè)置實(shí)驗(yàn)組和對照組,以每孔1×103個細(xì)胞接種于96孔板,每組設(shè)計(jì)4個復(fù)孔。在實(shí)驗(yàn)組和對照組中分別注入200 μL含不同濃度OXA的培養(yǎng)基和不含藥物的培養(yǎng)基,培養(yǎng)24 h。棄置液體后注入10 μL CCK-8染液,37 ℃下溫育1 h后于酶標(biāo)儀450 nm處測量吸光度(OD)值,計(jì)算細(xì)胞存活率/抑制率。
1.2.8聯(lián)合OXA干預(yù)HCT-116/OXA細(xì)胞株生物學(xué)活性實(shí)驗(yàn) 選取聯(lián)合OXA干預(yù)實(shí)驗(yàn)細(xì)胞株(分組同1.2.3),分別在6孔板中加入2 mL自噬干預(yù)劑或鐵死亡干預(yù)劑(藥物濃度同1.2.3)及等量生理鹽水預(yù)處理6 h,之后吸出培養(yǎng)液,取2 mL自噬干預(yù)劑或鐵死亡干預(yù)劑及OXA[OXA濃度為半數(shù)抑制濃度(IC50)=141.94 μmol/L]混合液加入實(shí)驗(yàn)組中,OXA對照組中加入等量生理鹽水,培養(yǎng)24 h后,進(jìn)行生物學(xué)活性實(shí)驗(yàn)。
1.2.9細(xì)胞侵襲試驗(yàn) Transwell小室模型置于24孔板中,制備濃度為每毫升5×105個細(xì)胞懸液。并于小室上層加入100 μL細(xì)胞懸液,下室加入600 μL含20%的胎牛血清培養(yǎng)基,每組細(xì)胞重復(fù)2個小室,連續(xù)培養(yǎng)24 h。用棉簽輕輕擦除上室側(cè)細(xì)胞和基質(zhì)膠,甲醇溶液固定30 min后用吉姆薩染液染色,鏡下觀察穿透細(xì)胞數(shù)。每個腔室隨機(jī)選取3個視野,計(jì)算平均值。
1.2.10平板克隆實(shí)驗(yàn) 制備細(xì)胞懸液,每組細(xì)胞分別以每孔500~1 000個細(xì)胞接種于6孔板內(nèi),并加入2 mL完全培養(yǎng)基,培養(yǎng)約1周,至產(chǎn)生肉眼可見的克隆球時結(jié)束。甲醇固定30 min后用吉姆薩染液染色,沖洗染色并風(fēng)干,拍照并計(jì)算各組的克隆形成數(shù)。
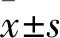
2 結(jié)果
2.1 不同細(xì)胞系中LC3、p62、GPX4的表達(dá)Western blot法檢測結(jié)果顯示(圖1A),LC3的表達(dá)量在HCT-8細(xì)胞中最低,在SW620細(xì)胞中最高(圖1B);p62的表達(dá)量在HCT-8細(xì)胞中最高,在SW620細(xì)胞中最低(圖1C);GPX4的表達(dá)量在HCT-8細(xì)胞中最高,在COLO205細(xì)胞中最低(圖1D)。選擇LC3中等表達(dá)量的HCT-116細(xì)胞株進(jìn)行后續(xù)實(shí)驗(yàn)。

圖1 結(jié)直腸癌細(xì)胞自噬關(guān)鍵蛋白和鐵死亡關(guān)鍵蛋白的表達(dá)水平:A.Western blot法檢測不同結(jié)直腸癌細(xì)胞系中LC3、p62、GPX4的表達(dá);B.LC3的表達(dá)量柱狀統(tǒng)計(jì)圖;C.p62的表達(dá)量柱狀統(tǒng)計(jì)圖;D.GPX4的表達(dá)量柱狀統(tǒng)計(jì)圖;*P<0.05,**P<0.01
2.2 結(jié)直腸癌敏感、耐藥細(xì)胞株中自噬和鐵死亡相關(guān)因子的表達(dá)選擇HCT-116細(xì)胞株,并委托原公司將其誘導(dǎo)成OXA耐藥細(xì)胞株HCT-116/OXA進(jìn)行后續(xù)實(shí)驗(yàn)。CCK-8實(shí)驗(yàn)檢測兩組細(xì)胞對OXA的敏感性,結(jié)果顯示:HCT-116細(xì)胞的IC50為9.87 μmol/L,耐藥細(xì)胞株HCT-116/OXA的IC50為141.94 μmol/L,明顯高于親本細(xì)胞株,差異有統(tǒng)計(jì)學(xué)意義(P<0.05,圖2A),提示耐藥誘導(dǎo)成功。采用Western blot法檢測兩組細(xì)胞中LC3、p62、Keap1、Nrf2、GPX4的表達(dá)。結(jié)果顯示,HCT-116/OXA細(xì)胞中p62、Nrf2、GPX4蛋白表達(dá)明顯高于HCT-116細(xì)胞株,而LC3、Keap1蛋白表達(dá)則顯著降低(P<0.05,圖2B、C)。試劑盒檢測兩株細(xì)胞中Fe2+、GSH、MDA的含量,結(jié)果顯示,HCT-116/OXA細(xì)胞中Fe2+、GSH、MDA的含量均高于HCT-116細(xì)胞(P<0.05,圖2D~F)。

圖2 結(jié)直腸癌敏感、耐藥細(xì)胞株中自噬和鐵死亡相關(guān)因子的表達(dá):A.CCK-8實(shí)驗(yàn)檢測敏感和耐藥細(xì)胞對OXA的敏感性;B.Western blot法檢測兩組細(xì)胞中LC3、p62、Keap1、Nrf2、GPX4蛋白表達(dá);C.兩組細(xì)胞中LC3、p62、Keap1、Nrf2、GPX4蛋白表達(dá)量柱狀統(tǒng)計(jì)圖;D.兩組細(xì)胞中Fe2+含量;E.兩組細(xì)胞中GSH含量;F.兩組細(xì)胞MDA中含量;*P<0.05,**P<0.01,***P<0.001,****P<0.000 1
2.3 干預(yù)自噬和鐵死亡對HCT-116/OXA細(xì)胞鐵死亡相關(guān)因子表達(dá)的影響選擇HCT-116/OXA細(xì)胞株進(jìn)行自噬和鐵死亡干預(yù)實(shí)驗(yàn),應(yīng)用MDC染色法觀察自噬活性,采用Western blot法檢測LC3、p62、Keap1、Nrf2和GPX4表達(dá)水平。結(jié)果顯示:自噬激活劑組中,Rapa組和Rapa+Fer-1組與其余兩組相比,細(xì)胞內(nèi)熒光顆粒顯著增多,LC3表達(dá)升高,p62表達(dá)降低,提示自噬被激活;Keap1均呈高表達(dá),Nrf2、GPX4均呈低表達(dá),但Rapa+Fer-1組中GPX4的表達(dá)高于Rapa組(P<0.05)。而Fer-1組與對照組相比各項(xiàng)指標(biāo)差異均無統(tǒng)計(jì)學(xué)意義(P>0.05,圖3)。

圖3 Rapa和Fer-1干預(yù)結(jié)直腸癌細(xì)胞后LC3、p62、Keap1、Nrf2及GPX4蛋白表達(dá)水平:A.MDC染色檢測不同干預(yù)組中的自噬水平;B.不同干預(yù)組中細(xì)胞自噬發(fā)生率;C.Western blot法檢測不同干預(yù)組的蛋白表達(dá)水平;D.LC3蛋白表達(dá)柱狀統(tǒng)計(jì)圖;E.p62蛋白表達(dá)柱狀統(tǒng)計(jì)圖;F.Keap1蛋白表達(dá)柱狀統(tǒng)計(jì)圖;G.Nrf2蛋白表達(dá)柱狀統(tǒng)計(jì)圖;H.GPX4蛋白表達(dá)柱狀統(tǒng)計(jì)圖;*P<0.05,**P<0.01,***P<0.001,****P<0.000 1
在自噬抑制劑組中,與對照組和Erastin組相比,CQ組和CQ+Erastin組細(xì)胞內(nèi)熒光顆粒顯著增多,LC3、p62表達(dá)同步升高,提示自噬活性降低;Nrf2均呈高表達(dá),Keap1均呈低表達(dá)(P<0.05)。對照組與Erastin組相比,CQ組與CQ+Erastin組相比,蛋白表達(dá)水平差異均無統(tǒng)計(jì)學(xué)意義(P>0.05)。GPX4在Erastin組中的表達(dá)顯著低于其余三組,而在CQ組中的表達(dá)則高于其余三組(P<0.05,圖4)。以上結(jié)果提示干預(yù)自噬可能通過p62-Keap1/Nrf2-GPX4通路調(diào)控結(jié)直腸癌細(xì)胞的鐵死亡。

圖4 CQ和Erastin干預(yù)結(jié)直腸癌細(xì)胞后LC3、p62、Keap1、Nrf2及GPX4蛋白表達(dá)水平:A.MDC染色檢測不同干預(yù)組中的自噬水平;B.不同干預(yù)組中細(xì)胞自噬發(fā)生率;C.Western blot法檢測不同干預(yù)組的蛋白表達(dá)水平;D.LC3蛋白表達(dá)柱狀統(tǒng)計(jì)圖;E.p62蛋白表達(dá)柱狀統(tǒng)計(jì)圖;F.Keap1蛋白表達(dá)柱狀統(tǒng)計(jì)圖;G.Nrf2蛋白表達(dá)柱狀統(tǒng)計(jì)圖;H.GPX4蛋白表達(dá)柱狀統(tǒng)計(jì)圖;*P<0.05,**P<0.01,***P<0.001,****P<0.000 1
2.4 干預(yù)自噬和鐵死亡對HCT-116/OXA細(xì)胞Fe2+、GSH、MDA含量的影響應(yīng)用試劑盒檢測不同干預(yù)組中Fe2+、MDA、GSH的表達(dá)量,并采用FerroOrange熒光探針檢測各組細(xì)胞內(nèi)Fe2+的含量。組間比較結(jié)果顯示,在自噬激活劑組中,Rapa組Fe2+、MDA的含量均高于其余三組,GSH的含量則低于其余三組(P<0.05)。Rapa+Fer-1組Fe2+、MDA含量低于Rapa組,而GSH含量高于Rapa干預(yù)組(P<0.05)。Fer-1組與對照組相比,各指標(biāo)的表達(dá)差異均無統(tǒng)計(jì)學(xué)意義(P>0.05,圖5A~C),提示激活自噬可能引起結(jié)直腸癌細(xì)胞發(fā)生鐵死亡。

GControlRapaFer-1Rapa+Fer-1ControlCQErastinCQ+Erastin
在自噬抑制劑組中,CQ組Fe2+、MDA的含量低于其余三組,GSH含量高于其他組;Erastin組Fe2+、MDA含量高于其余三組,GSH含量則低于其他組(P<0.05);與Erastin組相比,Erastin+CQ干預(yù)組的Fe2+、MDA含量低,GSH含量高(P<0.05,圖5D~G),提示下調(diào)自噬活性可能抑制結(jié)直腸癌細(xì)胞鐵死亡的發(fā)生。
2.5 自噬和鐵死亡干預(yù)劑聯(lián)合OXA對HCT-116/OXA細(xì)胞株增殖活性的影響CCK-8實(shí)驗(yàn)檢測自噬和鐵死亡干預(yù)劑預(yù)處理后聯(lián)合OXA對HCT-116/OXA細(xì)胞株增殖活性的影響。結(jié)果顯示,自噬激活劑組聯(lián)合應(yīng)用OXA后,Rapa干預(yù)組對OXA的化學(xué)敏感性高于其余三組,細(xì)胞增殖活性明顯降低;Rapa+Fer-1組對OXA的敏感性低于Rapa組,但高于Fer-1組和對照組,組間相比差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。Fer-1組細(xì)胞活性與對照組相比,差異無統(tǒng)計(jì)學(xué)意義(P>0.05,表1)。

表1 CCK-8實(shí)驗(yàn)檢測Rapa和Fer-1聯(lián)合OXA干預(yù)后HCT-116/OXA細(xì)胞株的存活情況
聯(lián)合應(yīng)用OXA后,在自噬抑制劑組中,Erastin干預(yù)組細(xì)胞活性顯著降低于其余三組,而CQ+Erastin組雖較Erastin組細(xì)胞活性升高,但細(xì)胞活性低于CQ和對照組,組間相比差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。CQ組細(xì)胞活性與對照組相比,差異無統(tǒng)計(jì)學(xué)意義(P>0.05,表2)。

表2 CCK-8實(shí)驗(yàn)檢測CQ和Erastin聯(lián)合OXA干預(yù)后HCT-116/OXA細(xì)胞株的存活情況
2.6 自噬和鐵死亡干預(yù)劑聯(lián)合OXA對HCT-116/OXA細(xì)胞株侵襲能力的影響Transwell侵襲實(shí)驗(yàn)顯示,聯(lián)合應(yīng)用OXA后,自噬激活劑組中Rapa組遷移細(xì)胞數(shù)量明顯少于其余三組;而Rapa+Fer-1組遷移細(xì)胞數(shù)量較Rapa組增多,但低于其余2組,差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。Fer-1組與對照組相比,差異無統(tǒng)計(jì)學(xué)意義(P>0.05,圖6)。

圖6 Rapa和Fer-1聯(lián)合OXA對HCT-116/OXA細(xì)胞侵襲能力的影響:A.Transwell細(xì)胞侵襲實(shí)驗(yàn)結(jié)果;B.不同干預(yù)組的遷移細(xì)胞數(shù);??P<0.01,???P<0.001,????P<0.000 1
自噬抑制劑聯(lián)合應(yīng)用OXA后,與對照組相比,Erastin組、CQ+Erastin組和CQ組細(xì)胞遷移數(shù)量均減少,其中Erastin組最低,CQ+Erastin組遷移細(xì)胞數(shù)量高于Erastin組、低于CQ組,各組間相比,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05,圖7)。

圖7 CQ和Erastin聯(lián)合OXA對HCT-116/OXA細(xì)胞侵襲能力的影響:A.Transwell細(xì)胞侵襲實(shí)驗(yàn)結(jié)果;B.不同干預(yù)組的遷移細(xì)胞數(shù);?P<0.05,??P<0.01,????P<0.000 1
2.7 自噬和鐵死亡干預(yù)劑聯(lián)合OXA對HCT-116/OXA細(xì)胞株克隆形成能力的影響平板克隆實(shí)驗(yàn)結(jié)果顯示,自噬激活劑聯(lián)合應(yīng)用OXA后,Rapa組的細(xì)胞克隆形成數(shù)較其余三組明顯減少;Rapa+Fer-1組細(xì)胞克隆形成數(shù)量較Rapa組增多,但較其余兩組少,組間相比差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。Fer-1組與對照組相比,差異無統(tǒng)計(jì)學(xué)意義(P>0.05,圖8)。

圖8 Rapa和Fer-1聯(lián)合OXA對HCT-116/OXA細(xì)胞克隆形成能力的影響:A.克隆形成實(shí)驗(yàn)檢測細(xì)胞增殖;B.不同干預(yù)組細(xì)胞克隆形成數(shù);**P<0.01,***P<0.001,****P<0.000 1
自噬抑制劑聯(lián)合應(yīng)用OXA后,Erastin組、CQ+Erastin組和CQ組三組細(xì)胞克隆形成數(shù)量均低于對照組(P<0.05);其中Erastin組的細(xì)胞克隆形成數(shù)量最少,CQ+Erastin組高于Erastin組,但低于CQ組,各組間相比,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05,圖9)。

圖9 CQ和Erastin聯(lián)合OXA對HCT-116/OXA細(xì)胞克隆形成能力的影響:A.克隆形成實(shí)驗(yàn)檢測細(xì)胞增殖;B.不同干預(yù)組細(xì)胞克隆形成數(shù);*P<0.05,***P<0.001,****P<0.000 1
自噬抑制劑聯(lián)合應(yīng)用OXA后,Erastin組、CQ+Erastin和CQ組三組細(xì)胞克隆形成數(shù)量均低于對照組(P<0.05);其中Erastin組的細(xì)胞克隆形成數(shù)量最少,CQ+Erastin組高于Erastin組,但低于CQ組,各組間相比,差異均具有統(tǒng)計(jì)學(xué)意義(P<0.05,圖9)。
3 討論
腫瘤耐藥是多基因、多信號通路的復(fù)雜過程,與藥物攝入減少、泵出增加、DNA修復(fù)能力增強(qiáng)、凋亡抑制等相關(guān)[10]。OXA作為中晚期結(jié)直腸癌一線化療藥物,其主要作用機(jī)制是與細(xì)胞核內(nèi)DNA交聯(lián),激活凋亡途徑,導(dǎo)致細(xì)胞死亡[11]。此外,OXA還可以通過破壞細(xì)胞內(nèi)的氧化還原穩(wěn)態(tài),導(dǎo)致細(xì)胞死亡[12]。但部分中晚期結(jié)直腸癌患者對OXA產(chǎn)生的耐藥性已成為影響預(yù)后的重要因素,因此,逆轉(zhuǎn)OXA耐藥,更好地發(fā)揮其持久的抗癌作用,提高患者的生存率,成為臨床醫(yī)師和研究人員關(guān)注的熱點(diǎn)。
鐵死亡是近年來發(fā)現(xiàn)的一種新型細(xì)胞死亡形式,是一種鐵依賴性脂質(zhì)過氧化物過度累積誘導(dǎo)細(xì)胞死亡的方式[4]。鐵死亡的發(fā)生機(jī)制復(fù)雜,主要通過內(nèi)、外兩種途徑啟動:外源性的轉(zhuǎn)運(yùn)蛋白依賴途徑和內(nèi)源性的酶調(diào)節(jié)途徑。其中,以抗氧化酶GPX4為主內(nèi)源性酶調(diào)節(jié)途徑最為經(jīng)典[13]。鐵死亡發(fā)生時,過量的鐵離子導(dǎo)致大量活性氧的產(chǎn)生與積累,GSH耗竭,GPX4活性下降,繼而氧化膜脂質(zhì),MDA升高,促使細(xì)胞膜完整性缺失,引發(fā)鐵死亡。GPX4是抗氧化體系中的關(guān)鍵分子,以GSH作為電子供體,使過氧化脂質(zhì)轉(zhuǎn)化為無毒的醇,減少對細(xì)胞損傷,從而抑制鐵死亡[14]。越來越多的證據(jù)表明,GPX4的降低是鐵死亡發(fā)生的關(guān)鍵標(biāo)志。Lu等[15]研究發(fā)現(xiàn),在腎透明細(xì)胞癌細(xì)胞中,抑制GPX4可激活鐵死亡,抑制細(xì)胞遷移和侵襲。Deng等[16]研究發(fā)現(xiàn),抑制GPX4表達(dá)可激活鐵死亡,增強(qiáng)肺腺癌細(xì)胞對順鉑的藥物敏感性。本研究結(jié)果顯示,不同結(jié)直腸癌細(xì)胞中GPX4的表達(dá)不同,雖然OXA耐藥細(xì)胞株中鐵離子及脂質(zhì)過氧化水平較高,但其GSH、GPX4表達(dá)水平顯著高于親本細(xì)胞株。作者認(rèn)為該現(xiàn)象符合腫瘤耐藥細(xì)胞的生物學(xué)特點(diǎn),高水平的鐵滿足了癌細(xì)胞的高代謝需求[17],而GPX4的增高又恰好增加了癌細(xì)胞的抗氧化應(yīng)激能力,抵抗鐵死亡。因此,通過降低GPX4表達(dá)引發(fā)腫瘤細(xì)胞鐵死亡,有望成為腫瘤耐藥治療的新靶點(diǎn)。
自噬是真核細(xì)胞內(nèi)高度保守的溶酶體介導(dǎo)的代謝途徑,主要負(fù)責(zé)清除錯誤折疊的蛋白質(zhì)、受損的細(xì)胞器等,對于維持細(xì)胞穩(wěn)態(tài)起重要作用[7]。作為重要的選擇性自噬接頭蛋白,p62被認(rèn)為是清除泛素化蛋白的重要自噬生物學(xué)標(biāo)志物,p62與泛素化的蛋白質(zhì)結(jié)合并以其為靶點(diǎn),通過自噬進(jìn)行降解,其含量隨自噬流的產(chǎn)生而降低[18]。Nrf2是一種堿性亮氨酸拉鏈轉(zhuǎn)錄因子,通過激活解毒酶、調(diào)控抗氧化基因和蛋白的表達(dá)來防止氧化損傷。GPX4已被確定為Nrf2的轉(zhuǎn)錄靶標(biāo),其表達(dá)可被Nrf2上調(diào)[19]。在正常生理情況下, Keap1與Nrf2結(jié)合并通過泛素-蛋白酶體途徑快速降解。而在氧化應(yīng)激狀態(tài)下,Keap1介導(dǎo)的Nrf2降解受到抑制,Nrf2被激活并釋放入核,發(fā)揮轉(zhuǎn)錄功能[20]。有研究發(fā)現(xiàn),自噬蛋白p62可以在Nrf2和Keap1的結(jié)合部位與Keap1競爭性結(jié)合,促進(jìn)Nrf2釋放,激活抗氧化系統(tǒng)。Li等[21]研究報道,激活p62-Keap1/Nrf2通路可減輕輻射所致肺損傷中的鐵死亡。然而,關(guān)于p62-Keap1/Nrf2-GPX4通路是否會影響結(jié)直腸癌鐵死亡的研究報道較少。本研究顯示,HCT-116/OXA細(xì)胞中鐵離子和脂質(zhì)過氧化水平均處于較高水平,且GSH、GPX4等抗氧化的活性同樣維持在較高水平。同時,p62、Nrf2、GPX4蛋白高表達(dá),提示HCT-116/OXA細(xì)胞的鐵死亡抑制和耐藥可能與p62-Keap1/Nrf2-GPX4通路處于異常激活狀態(tài)有關(guān)。因此,通過影響p62-Keap1/Nrf2-GPX4通路進(jìn)而促進(jìn)鐵死亡,有可能成為逆轉(zhuǎn)結(jié)直腸癌耐藥的新途徑。
自噬與鐵死亡關(guān)系密切,共同參與多種腫瘤的發(fā)生、發(fā)展,但目前兩者的關(guān)系尚不統(tǒng)一。Tang等[22]研究發(fā)現(xiàn),姜黃素通過激活自噬誘導(dǎo)非小細(xì)胞肺癌鐵死亡,增強(qiáng)非小細(xì)胞肺癌的治療效果。而在膠質(zhì)母細(xì)胞瘤干細(xì)胞中,抑制自噬可誘導(dǎo)鐵死亡的發(fā)生且增強(qiáng)細(xì)胞對替莫唑胺的敏感性[23]。然而,在結(jié)直腸癌中自噬對鐵死亡的影響尚未明確。本研究分別應(yīng)用自噬激活劑Rapa和自噬抑制劑CQ,觀察兩者對鐵死亡影響的分子機(jī)制。結(jié)果顯示,在結(jié)直腸癌HCT-116/OXA細(xì)胞中,應(yīng)用自噬激活劑Rapa后,LC3水平升高的同時伴隨p62水平降低、Keap1高表達(dá)、Nrf2低表達(dá),GPX4表達(dá)下降,鐵死亡生化指標(biāo)Fe2+、MDA含量上升,GSH含量下降,細(xì)胞存活率下降。應(yīng)用自噬抑制劑CQ后,結(jié)果與之相反。提示,干預(yù)自噬可通過p62-Keap1/Nrf2-GPX4通路影響鐵死亡。為進(jìn)一步證明自噬與鐵死亡的關(guān)系,本研究同步應(yīng)用了鐵死亡抑制劑Fer-1和激活劑Erastin。結(jié)果顯示,Rapa誘導(dǎo)的細(xì)胞死亡可被鐵死亡抑制劑Fer-1所抑制,而CQ抑制自噬則是通過提高抗氧化酶GSH和GPX4的表達(dá),從而抑制鐵死亡激活劑Erastin誘導(dǎo)的細(xì)胞鐵死亡。因此,本研究結(jié)果提示,自噬可通過p62-Keap1-Nrf2-GPX4影響鐵死亡,進(jìn)而影響細(xì)胞生存,干預(yù)此通路可能成為結(jié)直腸癌治療的新途徑。
許多研究表明,鐵死亡的激活有助于多種腫瘤的治療,在部分腫瘤中,鐵死亡可以增強(qiáng)化療藥物的療效。Du等[24]研究報道,雙氫青蒿素可通過誘導(dǎo)鐵死亡克服胰腺導(dǎo)管腺癌對順鉑的耐藥性。Song等[25]研究證實(shí),抑制GPX4可刺激細(xì)胞鐵死亡,增強(qiáng)三陰型乳腺癌對吉非替尼的敏感性。但干預(yù)自噬調(diào)控鐵死亡這一過程是否能逆轉(zhuǎn)耐藥腫瘤細(xì)胞對OXA的敏感性尚不清楚。本研究結(jié)果顯示,應(yīng)用自噬激活劑Rapa后誘導(dǎo)細(xì)胞發(fā)生鐵死亡,同時細(xì)胞對OXA的敏感性顯著升高,并且其增殖活性和遷移能力均顯著下降,而聯(lián)合應(yīng)用鐵死亡抑制劑Fer-1后OXA的作用效果顯著降低。聯(lián)合應(yīng)用鐵死亡激活劑Erastin后,細(xì)胞對OXA的敏感性升高,其增殖活性和遷移能力均顯著下降。作者認(rèn)為,在腫瘤細(xì)胞高鐵代謝的環(huán)境中,激活自噬可誘導(dǎo)鐵死亡的發(fā)生,進(jìn)一步增強(qiáng)了腫瘤細(xì)胞對OXA的敏感性。
綜上,本研究發(fā)現(xiàn)干預(yù)自噬可通過p62-Keap1/Nrf2-GPX4通路調(diào)控鐵死亡,從而逆轉(zhuǎn)結(jié)直腸癌對OXA耐藥性。因此,深入探索自噬與鐵死亡的關(guān)系,研究其在抗腫瘤治療中的潛在作用,為逆轉(zhuǎn)腫瘤耐藥提供新思路。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
昆明醫(yī)科大學(xué)學(xué)報(2022年1期)2022-02-28 07:43:40
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
