新型脫甲基酶抑制劑氯氟醚菌唑對水稻惡苗病致病菌藤倉鐮孢菌的抑菌活性
2024-03-11 08:49:18楊家偉KOKLANNOUDamalkSaintClaireSenakpon
農藥學學報 2024年1期
關鍵詞:水稻
陳 星, 楊家偉, KOKLANNOU Damalk Saint Claire Senakpon,孫 揚, 陳 雨*,
(1.安徽農業大學 植物保護學院,合肥 230036;2.作物有害生物綜合治理安徽省重點實驗室,合肥 230036)
水稻是世界上重要的糧食作物,是全球一半以上人口的主糧[1]。然而,水稻的產量和品質受到多種病原物的威脅。由藤倉赤霉復合種Gibberella fujikuroispecies complex 侵染引起的水稻惡苗病,是水稻生產中的重要種傳病害。該病害最早于1828年在日本首次發現,隨后在北美、歐洲、非洲等全球各水稻種植區均有發生[2-4]。水稻惡苗病在我國分布廣泛,主要集中于江西、湖南、廣東及長江中下游地區,可造成水稻產量損失10%~30%,嚴重時減產超過50%[5]。
藤倉赤霉復合種包括藤倉鐮孢Fusarium fujikuroi、層出鐮孢F.proliferatum、擬輪枝鐮孢F.verticilliodes及F.andiyazi,其中藤倉鐮孢菌是我國引起水稻惡苗病的主要致病菌,可造成幼苗徒長、葉片呈淡黃綠色,在成株期造成莖基部腐爛、分蘗減少,嚴重時整株枯死[6-8]。由于抗病品種的缺乏,防治水稻惡苗病仍以化學防治為主。目前,國內外用于防治水稻惡苗病的藥劑主要有苯并咪唑類 (多菌靈、甲基硫菌靈)、甾醇合成抑制劑類 (咪鮮胺、戊唑醇)、2-氰基丙烯酸酯類殺菌劑 (氰烯菌酯) 等[9-10]。但由于殺菌劑的長期單一及不合理使用,水稻惡苗病病原菌對常用殺菌劑產生了抗藥性[11-12]。此外,水稻惡苗病病原菌還會產生一些毒素,如伏馬菌素 (FBs)、赤霉酸 (GAs)、串珠鐮刀菌素等,這些真菌毒素具有神經毒性、細胞毒性及免疫毒性,嚴重威脅人類和其他動物的生命健康[5-6]。因此,水稻惡苗病的田間有效防控及抗性治理極為迫切。
氯氟醚菌唑 (mefentrifluconazole) 是巴斯夫公司于2018 年上市的第一個異丙醇三唑類殺菌劑,屬于新型脫甲基酶抑制劑(demethylase inhibitors,DMIs),主要作用細胞膜麥角甾醇生物合成過程中的C14-脫甲基酶,破壞病原菌細胞膜的結構[13-14]。氯氟醚菌唑可有效防治多種作物上的真菌病害,如黃瓜白粉病、番茄早疫病、芒果炭疽病等,并已在歐洲、美國等多個國家和地區登記使用[13,15-16]。據報道,氯氟醚菌唑對多種鐮孢菌屬病原菌具有不同的抑制效果。氯氟醚菌唑對西紅花球莖腐爛病菌尖孢鐮孢具有顯著的抑制效果[17],但對腐皮鐮刀菌抑制效果最差[15]。為明確氯氟醚菌唑對水稻惡苗病病原菌藤倉鐮孢菌的生物學活性,本研究明確了安徽省藤倉鐮孢菌對氯氟醚菌唑的敏感性分布,分析了氯氟醚菌唑對病原菌菌絲形態、孢子形態、細胞膜完整性和通透性的影響,同時測定了氯氟醚菌唑對藤倉鐮孢菌麥角甾醇和毒素生物合成的影響,以期為水稻惡苗病的田間防控及氯氟醚菌唑的科學合理使用提供理論依據。
1 材料與方法
1.1 供試材料
1.1.1 供試菌株 從安徽省不同地理區域采集的發病水稻幼苗和種子中共分離得到102 株水稻惡苗病致病菌 (表1),通過單孢分離純化,再經形態特征和分子生物學序列比對后鑒定為藤倉鐮孢菌F.fujikuroi[18],將菌株轉接至PDA 斜面培養基中于4 ℃保存,備用。2 株擬輪枝鐮孢菌F.verticillioides和1 株層出鐮孢菌F.proliferatum,由江蘇省農業科學院農產品質量安全與營養研究所提供。

表1 藤倉鐮孢菌菌株采集信息Table 1 Information of F. fujikuroi strains collected
1.1.2 供試藥劑 氯氟醚菌唑 (mefentrifluconazole)原藥和400 g/L 氯氟醚菌唑懸浮劑 (mefentrifluconazole 400 g/L SC),由巴斯夫股份公司提供,其中氯氟醚菌唑原藥用甲醇溶解。450 g/L 咪鮮胺水乳劑 (prochloraz 450 g/L EW) 和25%氰烯菌酯懸浮劑 (phenamacril 25% SC),分別由青島海納科技有限公司和江蘇省農藥研究所股份有限公司提供。
1.1.3 供試培養基 馬鈴薯葡萄糖瓊脂 (PDA) 培養基:稱取去皮馬鈴薯200 g,煮沸后過濾,隨后加入20 g 葡萄糖和20 g 瓊脂,攪拌溶解后加水定容至1 L;不添加瓊脂為PDB 液體培養基。水瓊脂 (WA) 培養基:稱取20 g 瓊脂,無菌水定容至1 L。酵母浸出粉胨葡萄糖 (YEPD) 培養基:10 g/L酵母提取物,20 g/L 蛋白胨,20 g/L 葡萄糖,加水定容至1 L。上述所有培養基均進行121 ℃高壓滅菌鍋滅菌20 min 后備用。
1.1.4 供試儀器設備 SW-CJ-2FD 超凈工作臺,上海博迅醫療生物儀器股份有限公司;Motic M150光學顯微鏡,合肥南達科學儀器有限公司;HITACHI S-4800 掃描電子顯微鏡,日本日立公司;LC-20AD XR 高效液相色譜儀,中國島津企業管理有限公司;OHAUS ST3100C 電導率儀,上海奧豪斯儀器有限公司。
1.2 試驗方法
1.2.1 藤倉鐮孢菌對氯氟醚菌唑的敏感性測定
分別采用菌絲生長速率法和孢子萌發法[5]測定。
1.2.1.1 菌絲生長速率法 在滅菌的PDA 培養基中加入氯氟醚菌唑,使其終濃度分別為0.05、0.1、0.2、0.4、和0.8 μg/mL。將藤倉鐮孢菌在PDA 平板上培養5 d 后,從菌落邊緣打取直徑為5 mm 的菌餅,轉接至上述含不同質量濃度氯氟醚菌唑的PDA 平板中央。以僅添加有同樣體積溶劑的PDA平板作為對照,每處理重復3 次。于25 ℃黑暗培養6 d 后,采用十字交叉法測量各處理的菌落直徑,按公式 (1) 計算各濃度藥劑處理對菌絲生長的抑制率(Im)。
式中:DC為對照組菌落生長直徑,cm;DT為處理組菌落生長直徑,cm;D0為菌餅直徑,cm。
1.2.1.2 孢子萌發抑制試驗 將藤倉鐮孢菌在PDA 平板上培養5 d 后,從菌落邊緣打取直徑為5 mm 的菌餅,轉接至100 mL PDB 液體培養基中,于25 ℃、175 r/min 搖培3 d。離心收集分生孢子,使用無菌水調整分生孢子濃度至105個/mL。取100 μL 孢子懸浮液涂布于含不同濃度氯氟醚菌唑的WA 平板上,于25 ℃黑暗培養8 h 后,鏡檢觀察孢子萌發情況。以芽管長度達到孢子直徑的1/2 及以上視為萌發,記錄孢子萌發數,以不含藥劑的WA 平板為空白對照,每板記錄300 個分生孢子萌發情況,每處理重復3 次。按公式 (2) 計算各濃度藥劑處理對孢子萌發的抑制率 (Is)。
式中:Sc為對照組菌株的產孢量;Ss為處理組菌株的產孢量。
利用DPS 軟件,計算得出藥劑質量濃度的lg 值 (x) 及抑制率的幾率值 (y) 之間的線性回歸方程和相關系數,求出EC50值。
1.2.2 氯氟醚菌唑對藤倉鐮孢菌菌絲和孢子形態的影響 采用掃描電子顯微鏡[19]觀察氯氟醚菌唑對藤倉鐮孢菌菌絲和孢子形態的影響。隨機選取3 株藤倉鐮孢菌,分別接種至含有0.05 μg/mL 氯氟醚菌唑的PDA 平板中央,于25 ℃黑暗培養5 d后,切取菌落邊緣大小為5 mm、厚度為1 mm 的菌塊,置于5%戊二醛溶液中固定過夜。按1.2.1.2節的方法收集藤倉鐮孢菌分生孢子,使用生理鹽水調整濃度至105個/mL,分別加入終濃度為相應EC50及EC90值的氯氟醚菌唑藥液,處理6 h 后,離心收集孢子于5%戊二醛溶液中固定過夜。隨后使用0.1 mol/L 的磷酸緩沖液沖洗3 次,相繼通過體積分數為30%、50%、70%、80%、95%、100%乙醇和丙酮進行洗脫,臨界點干燥后使用掃描電子顯微鏡進行觀察。
1.2.3 氯氟醚菌唑對藤倉鐮孢菌細胞膜通透性的影響 選取3 株藤倉鐮孢菌,在PDA 上培養5 d后,在菌落邊緣打取10 個直徑為5 mm 的菌餅,接種至100 mL YEPD 液體培養基中,于25 ℃、175 r/min 搖培48 h,每個菌株接種6 瓶。搖培48 h后,向其中3 瓶中加入終濃度為0.05 μg/mL 的氯氟醚菌唑藥液。以3 瓶不加藥劑的處理作為空白對照。繼續搖培48 h 后使用3 層紗布過濾、沖洗收集菌絲,風干。稱取0.3 g 菌絲,放入盛有20 mL蒸餾水的離心管中,分別在0、5、10、20、40、60、80、100、120、140、160 和180 min 時,使用電導率儀測定電導率。180 min 后將菌絲煮沸5 min,冷卻后測定最終電導率,計算出各時間點的相對電導率 (電導率/最終電導率)[20]。
1.2.4 氯氟醚菌唑對藤倉鐮孢菌細胞壁和細胞膜完整性的影響 選取3 株藤倉鐮孢菌,將其轉接至含有剛果紅 (congo red, CR) 質量濃度分別為0、0.05、0.1、0.2、0.4、0.8、1.2 和1.6 mg/mL 的加藥及不加藥的PDA 平板上,氯氟醚菌唑終濃度為0.05 μg/mL,以不含有剛果紅和氯氟醚菌唑的處理為對照組,評估氯氟醚菌唑對藤倉鐮孢菌細胞壁完整性的影響[20-21]。同樣,為測定氯氟醚菌唑對藤倉鐮孢菌細胞膜完整性的影響,將藤倉鐮孢菌接種至含有十二烷基硫酸鈉 (SDS) 質量濃度分別為0、0.05、0.1、0.2、0.4 和0.8 mg/mL 的加藥及不加藥的PDA 平板上,以不含有氯氟醚菌唑和SDS的處理為空白對照組,每處理重復3 次。于25 ℃黑暗培養6 d 后,用十字交叉法測定各處理的菌落直徑,按照公式 (1) 計算剛果紅及SDS 的實際抑制率。理論抑制率 (Ih) 按照公式 (3) 計算。
式中:RM為氯氟醚菌唑抑制率;RT為剛果紅/SDS 抑制率。
1.2.5 氯氟醚菌唑對藤倉鐮孢菌麥角甾醇含量的影響 隨機選取3 株藤倉鐮孢菌,分別接種至氯氟醚菌唑終濃度為0.05 μg/mL 的PDB 液體培養基中,于25 ℃、175 r/min 搖培5 d。收集菌絲,冷凍干燥后研磨成粉。取0.1 g 樣品于2 mL 無水乙醇中浸提過夜,隨后超聲處理40 min。靜置分層后,取1 mL 上清液過0.45 μm 有機濾膜,采用高效液相色譜 (HPLC) 法檢測麥角甾醇含量,流動相為100%甲醇,流速為1 mL/min。麥角甾醇保留時間以麥角甾醇標準品進行鑒定分析。試驗重復3 次。
1.2.6 氯氟醚菌唑對水稻惡苗病菌毒素含量的影響 將2 株擬輪枝鐮孢菌、1 株層出鐮孢菌[18]和3 株藤倉鐮孢菌分別接種至含有其對應EC50濃度氯氟醚菌唑的PDA 平板上培養7 d,以不加藥的處理為對照。將培養基粉碎后,稱取4 g 置于30 mL質量分數為80%的乙腈中,于180 r/min 搖培2 h,隨后將上清液過濾至離心管中。吸取4 mL 上清液氮吹至近干,用1 mL 質量分數為50%的乙腈復溶后過0.45 μm 濾膜,用于后續毒素檢測。按照仇劍波等[18]報道的方法對2 株擬輪枝鐮孢菌、1 株層出鐮孢菌和3 株藤倉鐮孢菌分別進行赤霉酸和伏馬菌素的檢測。
2 結果與分析
2.1 藤倉鐮孢菌對氯氟醚菌唑的敏感性
如圖1 所示,氯氟醚菌唑對藤倉鐮孢菌的菌絲生長和孢子萌發都具有良好的抑制效果,其對供試的102 株藤倉鐮孢菌菌絲生長的EC50值分布在0.0305~0.7579 μg/mL 之間,平均EC50值為(0.2469 ±0.0167) μg/mL,變異系數 (最高EC50值/最低EC50值) 為24.84。氯氟醚菌唑對供試84 株藤倉鐮孢菌孢子萌發的EC50值分布在0.1091~1.6870 μg/mL之間,平均EC50值為(0.6397 ± 0.0324) μg/mL。均未檢測到對氯氟醚菌唑具有抗性的藤倉鐮孢菌菌株。
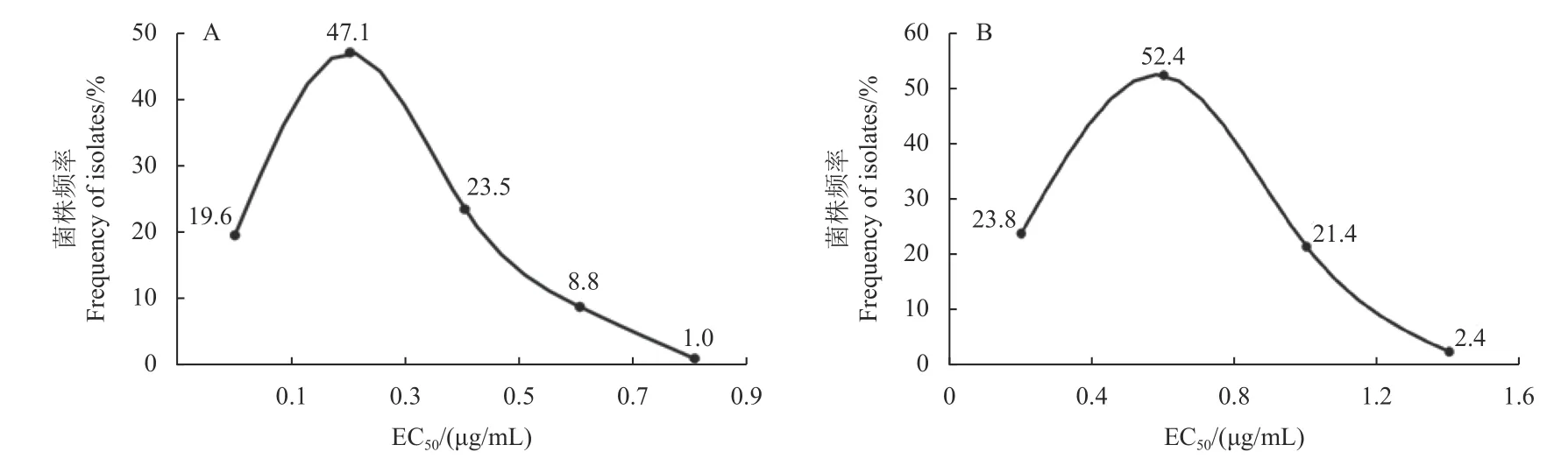
圖1 藤倉鐮孢菌菌絲 (A) 和孢子 (B) 對氯氟醚菌唑的敏感性分布Fig.1 Distribution of sensitivity of mycelia (A) and spores (B) of F. fujikurori to mefentrifluconazole
2.2 氯氟醚菌唑對藤倉鐮孢菌菌絲和孢子形態的影響
掃描電子顯微鏡觀察結果顯示,在0.05 μg/mL氯氟醚菌唑處理條件下,藤倉鐮孢菌菌絲形態出現不正常的扭曲變形,菌絲皺縮破裂,對照組呈現正常的菌絲形態 (圖2A)。氯氟醚菌唑處理組的孢子形態也出現皺縮的現象,且隨著藥劑質量濃度的增加,皺縮現象加劇。在EC90濃度的氯氟醚菌唑處理下,孢子變得扁平破裂,不再呈典型的孢子形態特征 (圖2B)。
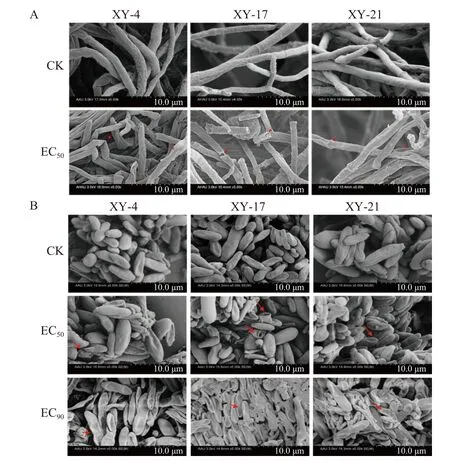
圖2 氯氟醚菌唑對3 株藤倉鐮孢菌(XY-4, XY-17 和XY-21)菌絲 (A) 和孢子 (B) 形態的影響Fig.2 Effect of mefentrifluconazole on the mycelial morphology (A) and spore morphology (B) of three F. fujikuroi strains (XY-4, XY-17 and XY-21)
2.3 氯氟醚菌唑對藤倉鐮孢菌細胞膜通透性的影響
氯氟醚菌唑處理下供試3 株藤倉鐮孢菌細胞膜透性的變化結果如圖3 所示:在0.05 μg/mL 氯氟醚菌唑處理條件下,XY-4、XY-17 和 XY-21 的電導率明顯高于未處理對照組,且隨著時間的延長,電導率均表現上升趨勢。表明氯氟醚菌唑可破壞藤倉鐮孢菌的細胞膜,導致胞內電解質的外滲,從而造成緩沖液中電導率增加。
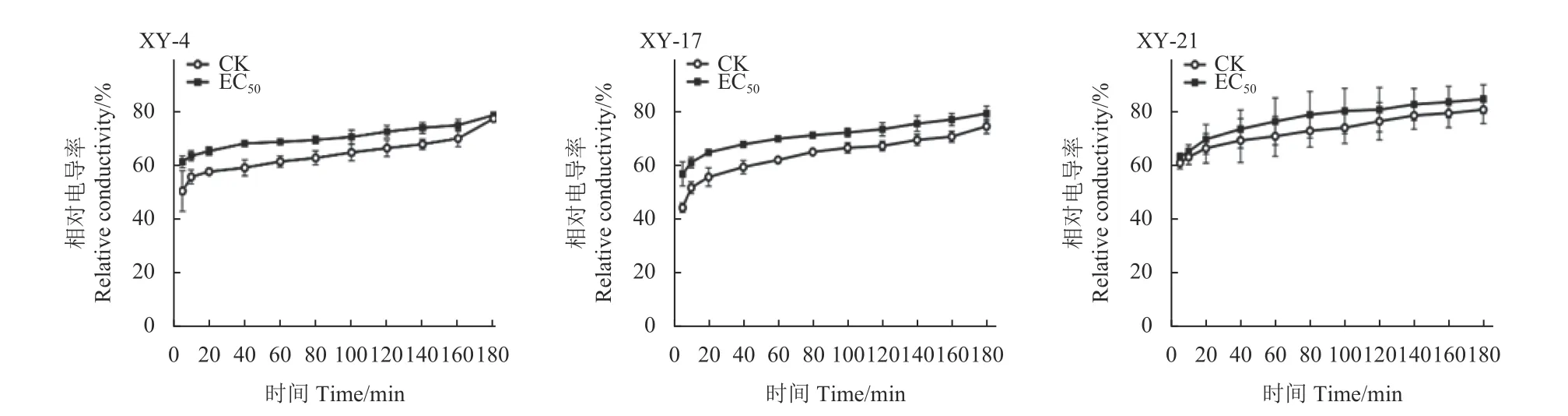
圖3 氯氟醚菌唑對3 株藤倉鐮孢菌細胞膜通透性的影響Fig.3 Effect of mefentrifluconazole on the membrane permeability of three F. fujikuroi strains (XY-4, XY-17 and XY-21)
2.4 氯氟醚菌唑對藤倉鐮孢菌細胞壁和細胞膜完整性的影響
測定結果表明,0.05 μg/mL 氯氟醚菌唑和0.05~1.6 mg/mL 剛果紅混合處理對藤倉鐮孢菌的菌絲生長抑制率高于其理論值,且隨著剛果紅質量濃度的增加,抑制率增加幅度顯著高于其理論抑制率 (圖4A)。而SDS 作為一種離子表面活性劑,可破壞細胞膜的完整性。氯氟醚菌唑與0.05~0.8 mg/mL SDS 混合處理對藤倉鐮孢菌菌絲生長抑制率明顯高于理論抑制率 (圖4B)。表明氯氟醚菌唑對藤倉鐮孢菌細胞壁和細胞膜的完整性具有破壞作用。
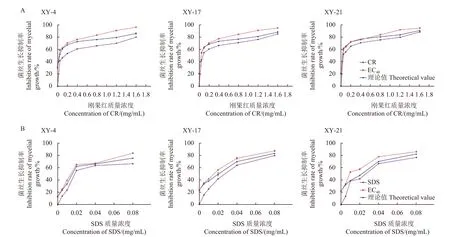
圖4 氯氟醚菌唑對3 株藤倉鐮孢菌細胞壁 (A) 和細胞膜 (B) 完整性的影響Fig.4 Effect of mefentrifluconazole on the cell wall (A) and the cell membrane (B) integrity of three F. fujikuroi strains
2.5 氯氟醚菌唑對藤倉鐮孢菌麥角甾醇生物合成的影響
測定結果如圖5 所示,氯氟醚菌唑可顯著抑制藤倉鐮孢菌麥角甾醇的生物合成。在0.05 μg/mL氯氟醚菌唑處理條件下,XY-4、XY-17 和XY-21 3 株藤倉鐮孢菌的麥角甾醇含量均顯著低于未處理組。
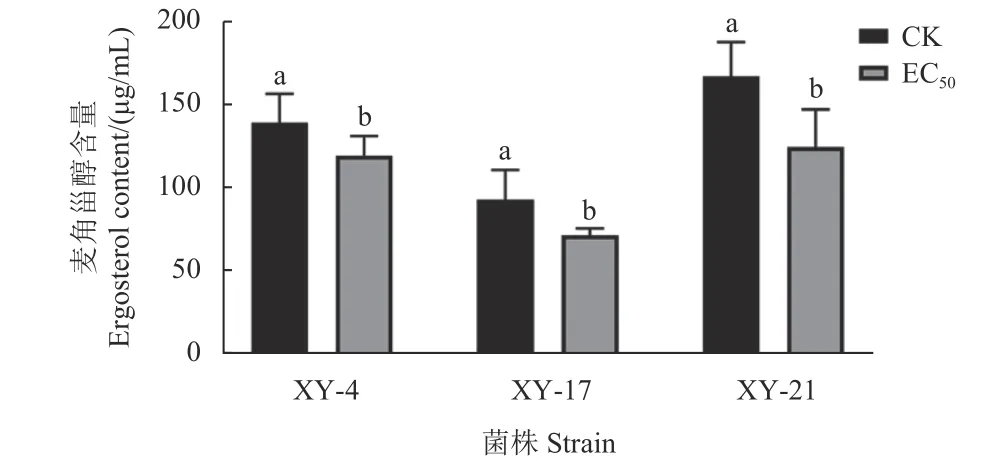
圖5 氯氟醚菌唑對3 株藤倉鐮孢菌麥角甾醇含量的影響Fig.5 Effect of mefentrifluconazole on the ergosterol content of three F. fujikuroi strains
2.6 氯氟醚菌唑對水稻惡苗病菌毒素合成的影響
為明確氯氟醚菌唑對水稻惡苗病菌毒素生物合成的影響,測定了在氯氟醚菌唑處理下3 株F.fujikuroi、2 株F.verticillioides和1 株F.proliferatum毒素含量的變化。結果如表2 所示,在F.fujikuroi中僅檢測到赤霉酸,而由于F.verticillioides和F.proliferatum不合成赤霉酸[18],故僅檢測到伏馬菌素。經氯氟醚菌唑處理后,雖然處理組F.fujikuroi中赤霉酸含量稍有下降,但與對照組相比無顯著差異,而氯氟醚菌唑處理可顯著降低F.verticillioides和F.proliferatum伏馬菌素 (FB1、FB2、FB3) 含量。

表2 氯氟醚菌唑對3 種水稻惡苗病菌毒素產生的影響Table 2 Effect of mefentrifluconazole on the production of several toxins of Fusarium spp.
3 結論與討論
氯氟醚菌唑其分子結構中的異丙醇基團能夠使其與麥角甾醇生物合成酶更緊密地結合[22-23]。在我國,氯氟醚菌唑已被登記用于防治柑橘等果樹炭疽病、番茄早疫病、蘋果褐斑病、香蕉葉斑病、馬鈴薯早疫病、黃瓜靶斑病和黃瓜白粉病等(http://www.chinapesticide.org.cn/zwb/dataCenter)。此外,已有文獻報道氯氟醚菌唑對桃褐腐病菌、小麥葉枯病菌等都具有很好的抑制效果[14]。相較于其他DMI 殺菌劑,氯氟醚菌唑活性更高,安全性更強[24]。文獻報道氯氟醚菌唑對水稻紋枯病菌Rhizoctonia solani的室內抑菌活性優于氟環唑、丙硫菌唑和丙環唑等三唑類殺菌劑,且其對水稻紋枯病具有良好的保護和治療活性[25]。有研究表明,氯氟醚菌唑對多種鐮孢菌屬病原菌抑制效果不盡相同[15]。
本文探究了氯氟醚菌唑對藤倉鐮孢菌孢子和菌絲的抑菌活性,并研究了其抑菌作用方式及對鐮刀菌毒素的影響。結果表明,氯氟醚菌唑對藤倉鐮孢菌菌絲生長和孢子萌發的EC50值分別分布在0.0305~0.7579 μg/mL 和0.1091~1.6870 μg/mL之間,平均EC50值分別為(0.2469 ± 0.0167) μg/mL和(0.6397 ± 0.0324) μg/mL,優于同類型殺菌劑丙硫菌唑、丙環唑對藤倉鐮孢菌的抑制活性[25],由此說明氯氟醚菌唑可以有效抑制藤倉鐮孢菌生長,且對菌絲生長的抑制作用優于對孢子萌發的抑制,同時未發現抗性菌株。劉亞慧等研究表明,氯氟醚菌唑對藤倉鐮孢菌的EC50值 < 0.172 μg/mL,本研究結果與其結果一致[15],但也有文獻報道,氯氟醚菌唑不能抑制尖孢鐮孢菌和灰葡萄孢菌的分生孢子萌發[16-17]。真菌細胞壁中幾丁質成分含量和菌絲對剛果紅的敏感性直接相關,而SDS 主要破壞細胞膜的完整性,在剛果紅或SDS 存在的情況下,氯氟醚菌唑對藤倉鐮孢菌的抑制效果高于理論抑制值,由此推斷氯氟醚菌唑可能通過降低藤倉鐮孢菌細胞壁中幾丁質含量,從而破壞細胞壁和細胞膜的完整性[20-21]。另外,氯氟醚菌唑作為一類廣譜三唑類殺菌劑,能夠降低細胞中麥角甾醇含量,破壞病原真菌細胞膜,引起細胞膜通透性的改變,從而導致細胞內容物泄漏[26-27]。在本研究中,氯氟醚菌唑可顯著降低藤倉鐮孢菌細胞膜中麥角甾醇含量,在0.05 μg/mL 氯氟醚菌唑處理條件下,不同菌株麥角甾醇含量均降低了25%以上,相對電導率顯著增加,菌絲和孢子形態發生改變,研究結果進一步證實了氯氟醚菌唑破壞病菌細胞膜的作用方式。
水稻惡苗病菌可產生包括赤霉酸、伏馬菌素、鐮刀菌酸等在內的多種毒素,這些毒素會污染糧食作物,造成人和動物急性或慢性中毒[18]。本研究結果表明,擬輪枝鐮孢和層出鐮孢菌主要產生伏馬菌素,而主要致病菌藤倉鐮孢產生的毒素以赤霉酸為主,這點與文獻報道[28-29]一致。同時本研究發現,氯氟醚菌唑可顯著降低擬輪枝鐮孢和層出鐮孢體外伏馬菌素的產生,但并不影響藤倉鐮孢菌中赤霉酸的合成,由此說明氯氟醚菌唑主要抑制伏馬菌素的生物合成。有研究結果表明,高濃度戊唑醇、苯醚甲環唑等殺菌劑會抑制毒素的生物合成,而低濃度會刺激毒素的生物合成[30-31],后續可考慮評估不同氯氟醚菌唑濃度對水稻惡苗病菌毒素生物合成的影響。
在農業生產中,使用殺菌劑進行種子處理是防治水稻惡苗病這類種傳病害最有效的措施。目前用于水稻惡苗病防治的殺菌劑主要有咯菌腈、咪鮮胺、戊唑醇等,但由于連年大量使用,抗藥性發生日趨嚴重,亟需篩選獲得用于水稻惡苗病防治的新型殺菌劑。氯氟醚菌唑殺菌譜廣,其分子中含有的異丙醇基團使其空間結構多變,可有效延緩田間抗藥性的發生,是用于病害防治的優異候選藥劑[32]。已有文獻報道,氯氟醚菌唑對灰霉病、辣椒炭疽病、西紅花球莖腐爛病等的保護活性優于治療活性[16-17,22],后續可進一步評價氯氟醚菌唑對水稻惡苗病的防治效果,為氯氟醚菌唑的田間合理使用提供理論依據。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
