不同溫度和脅迫時長下藍云杉的光合響應
2024-04-14 12:51:30歐陽芳群鄧軍育石青松李玉嶺
林業科學研究 2024年2期
歐陽芳群,崔 夏,張 輝,鄧軍育,石青松,李玉嶺,孫 猛*
(1. 植物遷地保護國家林業和草原局重點實驗室,北京 100093;2. 北京市花卉園藝工程技術研究中心,北京 100093;3. 北京市植物園管理處,北京 100093)
云杉屬(Picea)樹種多,分布廣,在陸地生態系統中占有特殊生態位,是高寒地區的生態建群種和重要用材樹種。根據聯合國政府間氣候變化專門委員會的第六次評估報告(https://www.ipcc.ch/report/ar6/wg2/),按照目前世界各國的減排措施,到本世紀末全球平均氣溫將上升2.7 ℃,遠高于《巴黎協定》控制全球氣溫上升的目標,將導致災難性的氣候變化。樹木年輪記錄隨著溫度的升高白云杉(Picea glauca(Moench) Voss)徑向生長降低[1]。分布在歐洲中部的歐洲云杉(Picea abies(L.) H. Karst.)因為夏季溫度超過其可忍受的臨界值而出現死亡[2]。云杉生長非常依賴其生長環境[3-4],全球持續暖化(www.epa.gov/climate-research)和降水量減少將不利于云杉生長[5-8],對云杉種的適合度和生產力將有嚴重影響[9]。
高溫脅迫會影響植物各種生理生化過程,其中光合作用是植物對溫度變化最敏感的生理過程[10],這會導致光系統II(PSII)受體的還原-氧化發生變化,并降低兩個光系統(PSⅠ和PSII)中光合電子傳遞效率[11]。植物光合作用的響應取決于熱脅迫的強度和持續時間[12]。中度熱脅迫引起光呼吸增加和氣孔導度降低,導致光合作用短暫降低;嚴重高溫脅迫或熱激下,由于類囊體膜完整性喪失和光系統失活, 葉片光合電子傳輸能力首先發生降低[13]。如果高溫脅迫時間超過植物耐熱閾值,便會導致光合機構發生不可逆的損害,進而對植物光合碳同化過程產生長遠影響。嚴重的傷害能顯著降低樹木的生長和抵抗病蟲害的能力,導致其死亡[14]。因此高溫脅迫對植物的傷害及其防御機制研究已越來越受到重視[15]。葉綠素熒光作為探測植物光合作用的“無損”探針,能夠有效反映幾乎所有光合作用過程的變化。高溫42~50 ℃處理白云杉30 min,其初始熒光(F0)隨溫度升高而平方增加,而最大熒光(Fm)、可變熒光(Fv)、PSII 的最大光化學效率(Fv/Fm)和光化學熒光淬滅 (qP)隨脅迫溫度升高而降低,在42~43 ℃ 處理后光系統II(PSII)失去了磷酸化的能力,44~46 ℃處理后嚴重限制了卡爾文循環利用NADPH 和ATP 的能力[16]。青杄(Picea wilsoniiMast.)在高溫35 ℃處理4 個月后Fv/Fm顯著低于25 ℃處理[17]。歐洲云杉在遭受了44 ℃短時熱激下,Fv/Fm沒有明顯變化,而52 ℃短時熱激下,PSII 的功能已明顯抑制,Fv/Fm降為對照組的63%,電子傳遞鏈的放氧復合體遭到了嚴重破壞,電子傳遞能力受到嚴重抑制[18]。和對照組相比,42 ℃熱處理小葉楊(Populus simoniiCarrière)3、6 h 后F0、Fv/Fm和qP沒有發生顯著變化,而處理12 和24 h 后Fv/Fm和qP顯著降低,但是F0增加[19]。因此植物對熱脅迫的響應不僅和脅迫的溫度有關,還依賴于時間長短。
高溫脅迫下,植物并非消極被動,能在生理生化及分子水平上發生一定變化來渡過脅迫期。但目前對云杉屬樹種的光系統對高溫脅迫的響應和適應機制尚待揭示。藍云杉(Picea pungensEngelm.)因其針葉顏色湛藍,觀賞價值高,是優良的園林景觀樹種。本課題組研究發現從濕潤地區引進的北美藍云杉表現出比鄉土種青海云杉(Picea crassifoliaKom.)更好的生長勢[20],推測其可能具有較好的適熱性。本研究以藍云杉樹種為對象,通過模擬高溫環境,研究藍云杉在不同溫度和不同脅迫時長下的植株形態變化、針葉相對含水量差異和PSI、PSII 的熒光參數響應熱脅迫的特點,分析熱脅迫解除后的恢復能力,為藍云杉在景觀生態等領域的應用和適熱性遺傳改良提供理論參考。
1 材料方法
供試的藍云杉苗木來自甘肅省小隴山林業實驗局林業科學研究所自育播種苗,種子來源美國亞利桑那州,苗齡均為5 a。在高溫脅迫處理前和恢復期,苗木生長在塑料大棚溫室里,平均溫度25 ℃,濕度75%,苗木生長在營養缽(直徑22 cm,高度22 cm)中,基質是體積比7:3 的腐殖質土和珍珠巖。75%的遮陽網用于夏季防曬。
(1)不同溫度高溫脅迫試驗:用恒溫恒濕實驗箱(HS-408)設定3個溫度處理(45、50、55 ℃)和常溫對照(25 ℃,CK,0 h)。處理時間為6 h(8:00 am—2:00 pm),每溫度處理4 株;
(2)不同耐熱時長高溫脅迫試驗:用恒溫恒濕實驗箱(HS-408)設定1 個溫度處理(45 ℃)和常溫對照(CK,0 h)。處理時間6 h(8:00 am—2:00 pm),12 h(6 h 重復2 d),18 h(6 h 重復3 d)和24 h(6 h 重復4 d)。每溫度處理4 株。
處理結束后0 d、恢復7 d 和恢復 28 d 統計植株葉色變化、落葉情況等。分別測定每株苗木的一年生枝條中段 2 cm 的針葉鮮質量和干質量(0.1 g)。計算葉片相對含水量:相對含水量=(鮮質量-干質量)/鮮質量 × 100%;每個溫度、耐熱時長處理下各選 3 株苗木測定,每株選擇 1 枝條。
每株苗木選擇1 份5~6 個完全展開成熟葉(并排用膠帶粘好),用HandPEA 儀器測定葉綠素熒光誘導動力學曲線(OJIP 曲線)和PSII 熒光參數[21](表1)。每處理3 個生物學重復。OJIP曲線是標準化后進行繪制的,標準化公式為Vt=(Ft-F0)/(Fm-F0)。式中:Vt表示任一時間點的熒光標準值,Ft表示任一時間點的熒光值。

表1 光系統熒光參數 Table 1 Fluorescence parameters of photosystem
同樣,每株苗木選擇1 份5~6 個完全展開成熟葉(并排用膠帶粘好),用熒光儀光纖型雙通道PAM-100 測量系統Dual-PAM-100 (Heinz Walz GmbH, Effeltrich, 德國)測定各處理下苗木PSI 和PSII 的葉綠素熒光活性[22]。測定的參數(表1)包括PSI 和 PSII 有效量子產量實際光化學效率Y(I)和 Y(II);PSI 和PSII 相對電子傳遞速率ETR(I)和ETR(II);受體側非光化學能量耗散產生的量子產量 Y(NA) 和由供體側限制引起的PSI 處非光化學能量耗散的量子產量Y(ND)。暗適應后最小和最大熒光產額F0和Fm。苗木暗適應20 min 后開始測定。
消費趨勢普遍代表著資本自由轉移,若非洲大陸利用好來自中國的市場,當地旅游業極有可能在GDP構成和創造就業等方面大大改變非洲。
(4)統計分析
采用SAS9.1 軟件GLM 模塊進行方差分析、多重比較。方差分析線性模型
式中:Xijk表示每株觀測值,μ表示試驗均值,Ti表示不同溫度或者耐熱時長效應(固定),eijk表示隨機誤差。用R 軟件作圖。
2 結果分析
2.1 不同溫度脅迫響應
2.1.1 表型響應 與對照相比,45 ℃處理6 h 后藍云杉表型未發生明顯變化(圖1A),且恢復28 d 時,仍與對照差別不大(圖1B);而50 ℃處理后藍云杉針葉變黃,55 ℃處理后藍云杉針葉顏色不但變黃,而且直接干枯掉落;恢復28 d 時,50 ℃和55 ℃處理的藍云杉整株針葉都變黃,基本干枯掉落(圖1)。

圖1 藍云杉高溫處理6 h(A)和恢復28 d 時(B)的表型變化Fig. 1 Phenotypic changes in Picea pungens with high temperature treatment for 6 hours (A) and recovery for 28 days (B)
2.1.2 針葉相對含水量差異 處理6 h 后、恢復7、28 d 時,針葉相對含水量在溫度處理間差異均極顯著(P<0.01,表2)。經DUNCAN 多重比較,針葉相對含水量在處理后和恢復7 d 時,45 ℃處理和對照差異不顯著,50 ℃和45 ℃之間差異也不顯著,相對含水量在0.60~0.66 之間;而恢復28 d 時只有45 ℃和對照無顯著差異,相對含水量為0.64,50 ℃處理后降至0.25。55 ℃處理后,恢復7 d 和28 d 相對含水量在0.15~0.19,遠低于對照(圖2)。

圖2 針葉相對含水量在不同溫度處理下多重比較,小寫字母代表鄧肯分析結果Fig. 2 DUNCAN comparison of the relative needle water content under different temperature treatments, and the lowercase letters represent the DUNCAN analysis results

表2 針葉相對含水量方差分析Table 2 Variance analysis of the relative water content of needles
2.1.3 葉綠素熒光參數 不同溫度處理后6 h、恢復7、28 d 時,除Y(NA) 在恢復7 d 時不顯著外,針葉PSI 熒光參數(Y(I)、ETR(I)、Y(ND)、Y(NA))在溫度處理間均差異顯著(P<0.01,表3)。經DUNCAN 多重比較(圖3)發現,處理后6 h,Y(I)在對照下是0.36,顯著高于50 ℃(0.20)、45 ℃(0.14)和55 ℃(0)。ETR(I) 和Y(I) 的趨勢一致。恢復28 d,45 ℃處理后的Y(I)和ETR(I)與對照無顯著差異,顯著大于50 ℃和55 ℃處理。這兩種溫度下Y(I)、ETR(I)、Y(ND) 和Y(NA)的值均為0。

圖3 光系統熒光參數在不同云杉種不同溫度處理下多重比較Fig. 3 DUNCAN comparation of chlorophyll fluorescence parameters of photosystem under different temperature treatments of different spruces
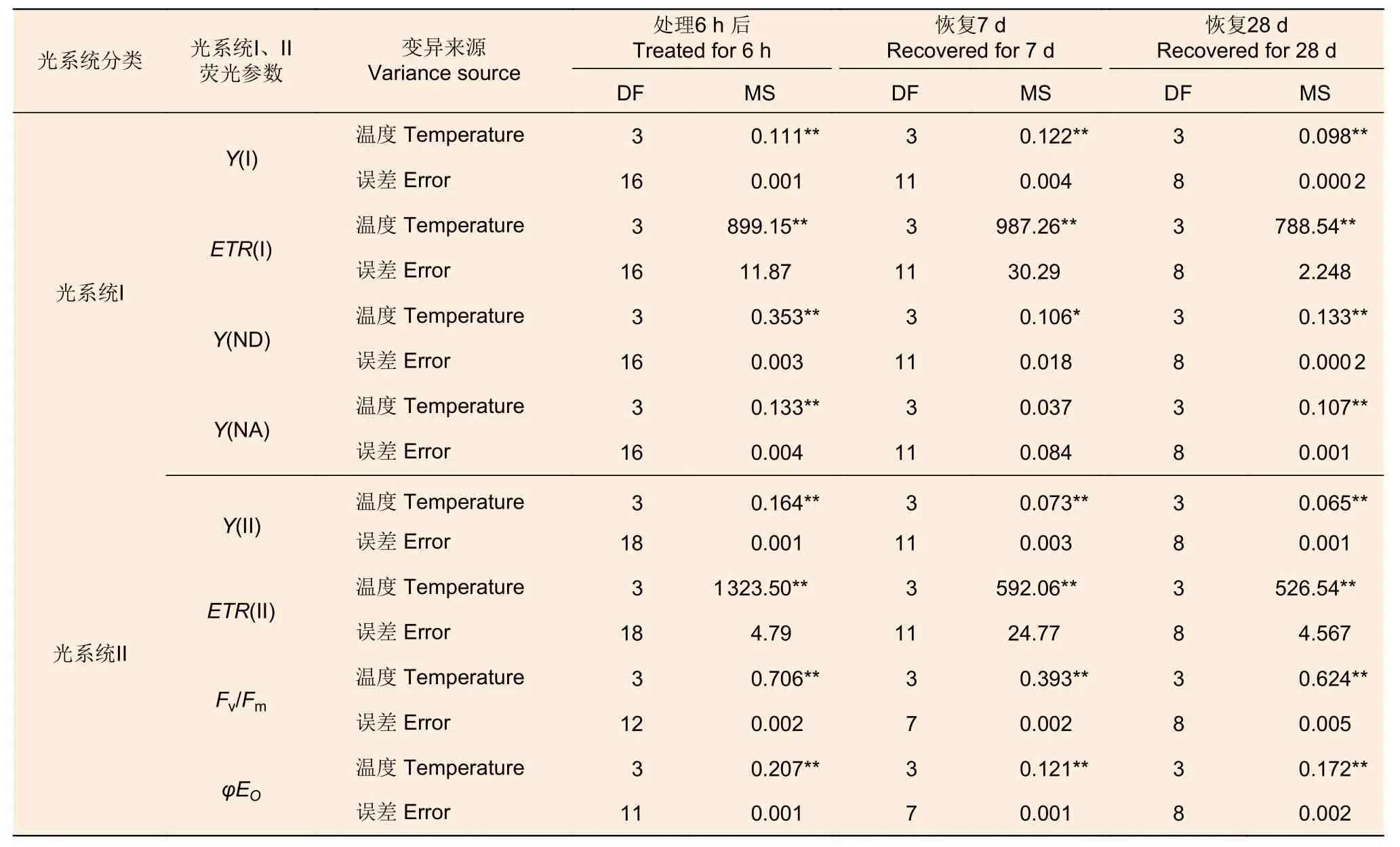
表3 光系統熒光參數方差分析 Table 3 Variance comparation of chlorophyll fluorescence parameters of photosystem
2.1.4 OJIP 曲線 藍云杉三種溫度處理6 h 后、恢復7 d 和恢復28 d 的經標準化后的OJIP 曲線(圖4)顯示45 ℃處理3 種云杉6 h 后OJ 相上升,且出現K 峰,至JI 相開始下降,到IP 相趨于平緩。和對照相比,恢復7 d 時候除了JI 相外其他基本恢復正常,到恢復28 d 時,OJIP 曲線基本恢復正常。而50、55 ℃處理6 h 后在恢復7 d 和恢復28 d 時,OJIP 曲線未恢復正常。

圖4 3 種溫度處理6 h 后(A)、恢復7 d(B)、恢復28 d(C)后的OJIP 曲線Fig. 4 OJIP curve of the three temperatures under CK, 45, 50 and 55°C for 6 h (A) and the recovery period 7 d (B) and 28 d (C).
2.2 藍云杉45 ℃不同耐熱時長響應
2.2.1 表型效應響應 和對照相比,45 ℃處理6、12、18、24 h 后藍云杉表型未發生明顯變化;恢復7 d 時,45 ℃處理6 h 依然變化不大,但是12、18 h 針葉顏色變黃,失水,24 h 處理幾乎整株針葉變黃;恢復28 d 時, 12、18 h 針葉繼續失水、變黃,24 h 處理整株針葉都變黃,且基本干枯掉落(圖5)。

圖5 藍云杉45 ℃處理后(A)、恢復7 d(B)、恢復28 d 時(C)的表型變化Fig. 5 Phenotypic changes in Picea pungens with high temperature treatment for 6 hours (A),recovery for 7 days and recovery for 28 days (B), from left to right
2.2.2 針葉相對含水量差異 針葉相對含水量在45 ℃不同耐熱時長處理后6 h、恢復7 d、28 d差異顯著或極顯著(P<0.05 或者P<0.01,表4),經DUNCAN 多重比較,針葉相對含水量在處理后時長18 h 處理下值為0.45,顯著低于其他時長處理和對照(0.62~0.66)。而恢復7 d 時,12、18 h處理后的相對含水量0.77~0.79,顯著高于對照、6 h 和24 h 處理(分別為0.67、0.64、0.68)。恢復28 d 時,對照、6 h、12 h 處理的針葉相對含水量之間無顯著差異,顯著高于24 h 處理(0.24),18 h 處理居中(0.47,圖6)。

圖6 針葉相對含水量在45 ℃不同時長處理下多重比較Fig. 6 DUNCAN comparation of the relative needle water content under different duration with 45 ℃ treatment

表4 45 ℃不同耐熱時長處理藍云杉針葉相對含水量方差分析Table 4 Variance analysis of the relative water content of needles of Picea pungens under different duration with 45 ℃ treatment
2.2.3 葉綠素熒光參數 45 ℃處理后,針葉PSI 熒光參數(Y(I)、ETR(I)、Y(ND)、Y(NA))在耐熱時長處理間差異極顯著(P<0.01,表5)。經DUNCAN 多重比較,對照下Y(I)值為0.36,顯著高于6 h(0.14)、12 h(0.11)等全部處理,12、18 和24 h3 種時長處理間差異不顯著(0.08~0.11)。ETR(I) 和Y(I) 趨勢一致。Y(ND) 趨勢則相反,對照處理下值最低(0.27),6 h 處理下最高0.64,12、18 h 間無顯著差異(0.55、0.49)。Y(NA)在對照、12、18 h 處理間差異不顯著,低于24 h 處理,6 h 處理居中。恢復7 d 時,Y(I)、ETR(I) 在耐熱時長處理間差異極顯著(P<0.01);恢復28 d 時,耐熱時長處理顯著影響Y(I)、ETR(I)和Y(ND) 值(P<0.05)。經DUNCAN 多重比較,恢復7 和28 d,Y(I)、ETR(I)在時長6 h處理值最高,分別為0.37、33.56 和0.32、28.7,和對照無顯著差異,顯著高于其他3 種時長處理;恢復28 d 時,Y(ND)在時長24 h 處理值最高,其他時長處理間無顯著差異。
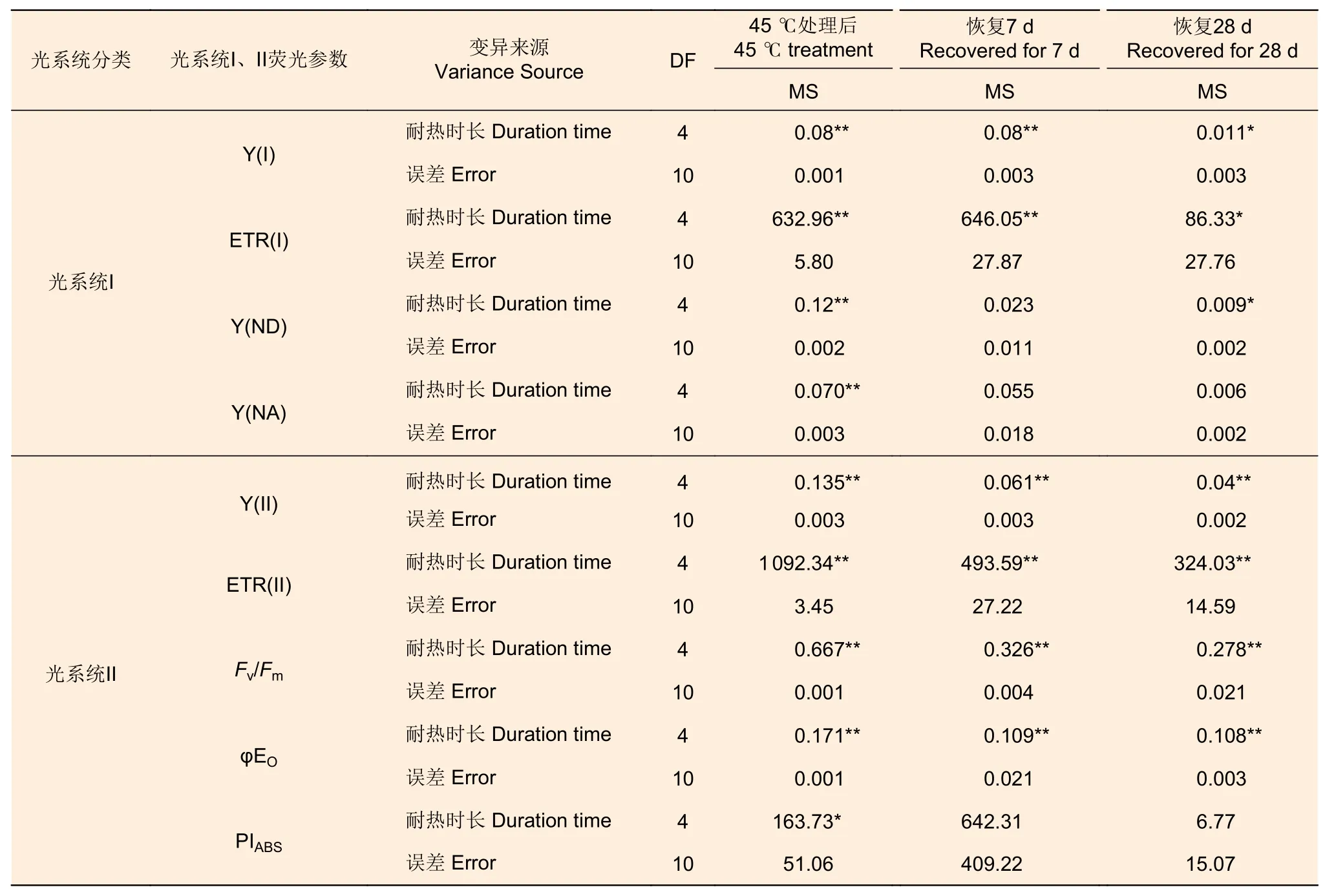
表5 45 ℃不同耐熱時長處理藍云杉熒光參數方差分析 Table 5 Variance analysis of chlorophyll fluorescence parameters of needles of Picea pungens under different duration with 45 ℃ treatment
針葉PSII 熒光參數(Y(II)、ETR(II)、Fv/Fm、φEo)在處理后、恢復7、28 d 在耐熱時長處理間差異均極顯著(P>0.01,表5)。經DUNCAN 多重比較,處理后,Y(II) 的值在對照下最高(0.33),其他各時長值均為0。恢復7 d 時,時長為6 h 的處理Y(II)值由0 恢復到0.25,和對照無顯著差異,顯著高于其他時長處理(幾乎為0)。ETR(II) 和Y(II) 趨勢基本一致。在6 h 時長處理下,Fv/Fm(0.11)顯著低于對照(0.86),而其他時長Fv/Fm幾乎為0。恢復7 d 時,6 h 時長處理Fv/Fm由0.11 恢復到0.47,其他時長依然幾乎是0。恢復28 d 時,6 h 時長處理恢復到0.75,和對照無顯著差異,12、18 和24h 時長處理分別恢復到0.33、0.21、0.19,顯著低于對照。φEo的變化趨勢和Fv/Fm基本一致,只是18 h 和24 h 處理后和恢復期φEo幾乎為0(圖7)。
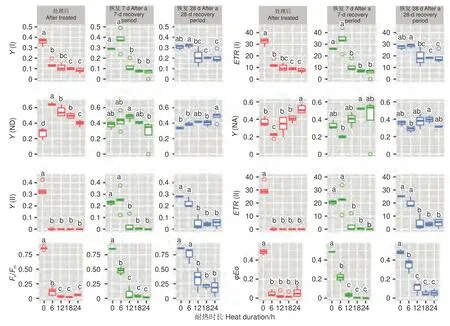
圖7 針葉熒光參數在45 ℃不同時長處理下多重比較Fig. 7 DUNCAN comparation of chlorophyll fluorescence parameters under different duration with 45 ℃ treatment
2.2.4 OJIP 曲線 藍云杉不同耐熱時長處理后、恢復7 d 和恢復28 d 的經標準化后的OJIP 曲線(圖8)顯示45 ℃處理6 h 后OJ 相上升,且出現K 峰,至JI 相開始下降,到IP 相趨于平緩。恢復7 d 時候除了JI 相外其他基本恢復正常,到恢復28 d 時,OJIP 曲線基本恢復正常。而處理12、18 h 后恢復7 d 和恢復28 d 時,OJIP 曲線KJ、JI相偏離對照較多,處理24 h 后的曲線一直未恢復。

圖8 5 種耐熱時長處理后(A)、恢復7 d(B)、恢復28 d(C)后的OJIP 曲線Fig. 8 OJIP curve of the three temperatures under five durationsfor after treatment (A) and the recovery period 7 d (B) and 28 d (C).
3 討論
3.1 藍云杉對高溫的熱適應
在較冷地區生長的樹木通常不耐高溫[23]。適中的溫度促進云杉生長[24]。當溫度超過植物生長的最佳溫度,光合作用和呼吸作用失衡可能會導致生長受到抑制[25-26]。夏季高熱對云杉生長有負面影響[27],比如麗江云杉(Picea likiangensis(Franch.)E. Pritz.)、林芝云杉(Picea likiangensisvars.linzhiensisW. C. Cheng & L. K. Fu)、紅皮云杉(Picea koraiensisNakai)和白杄(Picea meyeriRehder & E. H. Wilson)4 種云杉在高溫(白天溫度35 ℃)下生物量降低[5],生長受到抑制[26,28],表明35 ℃超過了適宜云杉生長的最佳溫度[25]。研究發現9 種云杉屬植物半致死溫度在45~50 ℃之間[29]。本研究中藍云杉可耐45 ℃處理6 h,處理后表型變化小,且在28 d 后基本能恢復,但不耐50 ℃和55 ℃處理。這可能與其針葉相對含水量是分不開的。恢復期,只有45 ℃處理6 h 的苗木針葉相對含水量和對照無明顯差異,50 ℃和55 ℃處理遠低于對照。而通常熱脅迫和干旱是連在一起的,失水傷害是影響苗木質量的首要因素。干旱似乎是云杉生長的最大限制因素。當水勢下降到-2.0~3.0 MP 之間時,黑云杉(PiceamarianaBritton, Sterns & Poggenb.)、西加云杉(Picea sitchensis(Bong.) Carrière)和白云杉氣孔迅速關閉,凈光合速率也會隨水勢的下降,降到光補償點[30]。50 ℃和55 ℃處理藍云杉在恢復期整株針葉顏色變黃并伴有針葉脫落。雖然落葉是一種應對脅迫的積極反應,但是藍云杉在這個過程中失水嚴重,恢復28 d 時相對含水量均顯著低于45 ℃處理和對照。Tabbush 和 Paul 發現西加云杉苗木晾曬3 h 18 min 后,2 a 后的優樹保存率下降到68%,同時高生長也受到影響[14]。所以在極端高溫脅迫出現時,溫室可以通過降低葉面溫度保護植物不被高溫損傷。比如適時通風、外遮陽、噴霧降溫、增濕等[31],以減少脅迫程度和脅迫時間。此外,氣溫上升可能會對樹木的碳儲存產生負面影響[32]。對高溫的高度敏感性間接導致植株碳饑餓[33]。在35~40 ℃的溫度下,紅云杉(PicearubensSarg.)表現出零碳平衡或負碳平衡[34]。因此,溫度升高可能導致儲備的碳水化合物過度消耗[35]。
3.2 藍云杉對不同時長的熱適應
熱損傷不僅與溫度有關,還與高溫持續時間有關[12]。較低的溫度和較短的暴露時間通常導致間接葉損傷,而較高的溫度和較長的暴露時間會發生直接葉損傷[12]。光合功能退化和細胞損傷程度依賴于累積的熱量,和溫度、熱脅迫的時間長短是相關聯的[12]。同樣時間下溫度越高或者同樣的溫度,脅迫時間越長累積的熱量越多,葉片受損更嚴重。白云杉在48~48.5 ℃下30 min 產生50% 的葉損傷,而52 ℃及以上的溫度30 min 則可以產生100%的葉損傷[16]。高溫對黑云杉的破壞作用隨暴露時間呈指數級變化,溫度越高,造成破壞所需的時間越短。50% 黑云杉幼苗受到直接損傷需要在暴露47 ℃下51 min,而 48 ℃下只需要24 min[36]。當幼苗持續在高溫下時長達180 min 時,黑云杉葉損傷閾值低至44 ℃[36]。當高溫脅迫時間達到一定閾值時,脅迫所造成的損傷無法恢復[19,36]。本研究中,藍云杉在45 ℃處理6 h 可以恢復正常生長,而45 ℃處理24 h 后整株針葉發生損傷,干枯掉落,植株無法恢復正常生長。楊樹在42 ℃下熱激脅迫短于6 h 時,PSII 經過自我修復可恢復正常功能,但超過6 h 就會造成不可逆轉的損傷[19]。
3.3 藍云杉對高溫脅迫的光合響應
眾所周知,PSII 在對逆境的光合反應中起著重要作用[37-38]。光合作用對熱應激敏感,尤其PSII 高度敏感[11,39]。Fv/Fm比值是研究植物對逆境響應的重要生理指標,它用來度量PSII 原初光能轉換效率,常被作為植物是否發生光抑制的標準之一[40]。Fv/Fm比值是決定PSII 最大量子效率的重要參數,它反應線性電子傳遞速率和整體光合能力。Bj?rkman 發現,植物在正常情況下(包括不同生態類型),其Fv/Fm比值均穩定在0.832 ± 0.004左右[41]。在不同脅迫下該比值出現不同程度下降,表示一定比例的PSII 反應中心被破壞或失活,這種現象稱為光抑制[42]。45 ℃處理6 h 后Fv/Fm低至0.07,50、55 ℃處理后、恢復7 和28 d,藍云杉的Fv/Fm都為0,表明PSII 反應中心可能發生了不可逆失活,或光合機構受到破壞或損傷[43]。研究50、55 ℃處理6 h 和45 ℃處理大于6 h 后藍云杉的針葉發黃可能是Fv/Fm值下降的原因之一,因為針葉發黃常常意味著發生光氧化和葉綠素濃度降低[44]。對歐洲冷杉(Abies albaMill.)的研究中,初始脅迫溫度范圍是 42.7 ~43.2 ℃,Fv/Fm降至 50%的極限脅迫溫度是51 ℃[45]。黑云杉在 23 ℃時,凈光合速率就會顯著下降;而當氣溫達到或大于35~40 ℃時,光合速率將會很低甚至停止[26]。本研究中,45 ℃處理6 h 后恢復期,Fv/Fm恢復至0.75,和對照無顯著差異,表明PSII 恢復正常。φEo的變化趨勢和Fv/Fm基本一致,只是50、55 ℃,18 、24 h 處理后和恢復期φEo幾乎為0。表明QA后的電子傳遞受到了抑制,PSII 的受體測遭到了毀壞[46]。
盡管和PSII 相比,PSI 在高溫脅迫中比較穩定[11]。但有證據表明,在高溫下存在相當大的環式電子流,維持跨類囊體膜的能量以及三磷酸腺苷穩態,可能參與預防高溫條件下的不可逆損傷[47-48]。藍云杉經過45 ℃處理6 h 后PSI 的Y(I)、ETR(I)、Y(ND)和Y(NA) 在恢復期和對照差異不顯著,表明45 ℃溫度處理6 h 對藍云杉PSI 影響不大。PSI 通過維持強氧化態(P700+),將PSI 受體測的過剩光能耗散吊,減少PSI 損傷[49]。45 ℃處理12、18、24 h 后PSI 多少有些恢復,而50、55 ℃處理后的恢復過程中藍云杉PSI 的Y(I)和ETR(I)均為0,表明PSI 遭受到了嚴重損壞。PSI 一旦發生抑制,由于恢復時間較長,往往對光合作用造成更大的影響[50]。
4 結論
藍云杉的熱損傷不僅與溫度有關,還與高溫脅迫的時間有關。藍云杉可耐45 ℃處理6 h,但不耐50 和55 ℃處理6 h,藍云杉PSI 和PSII 遭受到了嚴重損壞。在45 ℃處理時長超過12 h,φEo幾乎為0,針葉發黃掉落,難恢復正常生長。因此,隨著全球氣候變暖,持續的夏季高溫將不利于藍云杉的生長。本研究為進一步深入探討藍云杉耐高溫機制研究提供了基礎。
致謝:感謝北京市公園管理中心科技課題“云杉屬樹種引種、抗高溫評價與新優種質高效繁育”(ZX2021012)和中央財政林業科技推廣示范項目“藍云杉栽培繁育技術推廣示范”(京[2023]TG05 號)資助!感謝林木遺傳育種國家重點實驗室科研儀器平臺。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
