日本落葉松胚狀體干化處理對萌發的影響
2024-04-14 12:51:34孫海濤齊力旺李萬峰
林業科學研究 2024年2期
關鍵詞:分類
孫海濤,楊 玲,齊力旺,李萬峰*
(1. 林木遺傳育種全國重點實驗室 東北林業大學林學院,黑龍江 哈爾濱 150040;2. 林木遺傳育種全國重點實驗室國家林業和草原局林木培育重點實驗室 中國林業科學研究院林業研究所,北京 100091)
日本落葉松(Larix kaempferi(Lamb.) Carr.)為松科(Pinaceae)落葉松屬(Larix)喬木,廣泛分布于溫帶和寒溫帶等地區。它是我國重要的用材樹種,具有早期速生、病蟲害少、輪伐期短、成林快、抗逆性強等特點。基于有性繁殖的落葉松傳統育種存在優良性狀難以保持、生長周期長等問題極大地限制著日本落葉松優異種質資源的大規模利用。和扦插、嫁接相比,日本落葉松體細胞胚胎發生技術具有遺傳穩定性強、繁殖系數高等優點,已成為當前最有潛力的無性繁殖技術,受到越來越多的關注[1-6]。
體細胞胚胎發生(Somatic embryogenesis)是指體細胞經歷脫分化并在體外形成類似胚胎的胚狀體,這一過程與合子胚的發育過程有許多相似之處[7-8]。在1987 年von Aderkas 以雜種落葉松(L.ecidua×L.leptolepis)雌配子體為外植體誘導出胚性愈傷,但未獲得再生植株[9]。Klimaszewska以歐日雜種落葉松(Larix×eurolepis)未成熟種子為外植體,成功培育出再生植株[10]。此后體細胞胚胎發生技術在多種雜種落葉松[11-12]、華北落葉松(L.gmeliniivar.principis-rupprechtii(Mayr) Pilger)、西伯利亞落葉松(L.sibiricaLedeb.)[13-14]、長白落葉松(L.olgensisHenry)[15-16]和日本落葉松[17-18]等落葉松中都得以應用并取得一定的進展,但依然存在一些問題亟待解決,比如胚狀體萌發率和成苗轉化率低等,這些問題限制了日本落葉松體胚發生技術的發展和應用。
在萌發培養前對胚狀體進行干化處理是解決上述問題的一種可行的方法。干化處理可以將“形態成熟”的胚狀體轉化成“生理成熟”的胚狀體,從而促進萌發[19-20]。這在云杉屬(Picea)體細胞胚胎發生技術體系中得到很好的應用,被認為是體細胞胚成功萌發的先決條件[21-22],但在日本落葉松體細胞胚胎發生技術體系中尚未見到過相關的研究和應用報道。
本研究以日本落葉松胚狀體為材料,觀察并統計不同類別胚狀體的萌發情況,探究干化處理對日本落葉松胚狀體萌發的影響,旨在提高日本落葉松胚狀體的萌發率,促進日本落葉松高效穩定再生體系的建立,為日本落葉松良種繁育和規模化應用提供技術支撐。
1 材料與方法
1.1 植物材料與培養條件
本實驗以日本落葉松C6 細胞系為材料,所用到的增殖培養基、成熟培養基和萌發培養基均參考齊力旺的方法[23]。培養基pH 為5.8,121 ℃高壓滅菌后,在無菌環境下分裝備用。增殖培養和成熟培養在暗環境(溫度22 ℃)下進行,萌發培養先在暗環境下進行3 d,之后在光環境(溫度24 ±2 ℃,光照14 h,光照強度1 000 lx)下進行。
1.2 方法
1.2.1 胚狀體形態分類 胚性愈傷組織成熟培養45 d 后,隨機選取100 個胚狀體,根據子葉數量和胚軸形態進行形態分類,顯微鏡下觀察、分類、拍照,統計不同類別胚狀體的發生頻率。成熟培養共進行4 次,共分析400 個胚狀體。
1.2.2 胚狀體萌發 重新選取300 個胚狀體進行分類與萌發培養。2 個月后統計其萌發情況。觀察到子葉伸長、胚軸生長和胚根伸長,分別用“√”標記,反之,用“ × ”標記,培養條件同上。
1.2.3 干化處理對胚狀體萌發和形態的影響 類型Ⅰ胚狀體采用“濾紙容器法”進行干化處理[24]。處理時,用鑷子將胚狀體平鋪在有3 層干濾紙(70 mm)的塑料培養皿(90 mm × 20 mm)中,隨后再將無蓋的塑料培養皿放在裝有15 mL 去離子水的玻璃培養皿(180 mm × 31 mm)中;玻璃培養皿用封口膜封口,隨后放置在光環境下(溫度24 ± 2 ℃,光照14 h,光照強度1 000 lx)。干濾紙、培養皿和去離子水在使用前進行高壓滅菌。干化處理分別進行0、7、14、21 和28 d。每次處理后取180 個胚狀體放入萌發培養基中,每皿60 個,重復3 次,30 d 后統計其萌發情況。
1.2.4 數據處理 本實驗以子葉和胚軸伸長、胚根長出作為胚狀體萌發的標準。
式中:y 為胚狀體萌發率,n 為萌發的胚狀體數量,N 為參與萌發培養的胚狀體數量。采用軟件IBM SPSS Statistics19.0進行方差分析。
2 結果與分析
2.1 日本落葉松胚狀體的形態和分類
日本落葉松C6 胚性愈傷組織經過成熟培養后產生形態各異的胚狀體。進一步觀察和分析后,胚狀體可分為10 種類型(圖1、表1),其中類型I 發生頻率最高,達到56.75%;類型Ⅲ、類型Ⅵ發生頻率分別為12.75%、13.25%,僅次于類型I;其余類型的發生頻率較低,都不足5%。
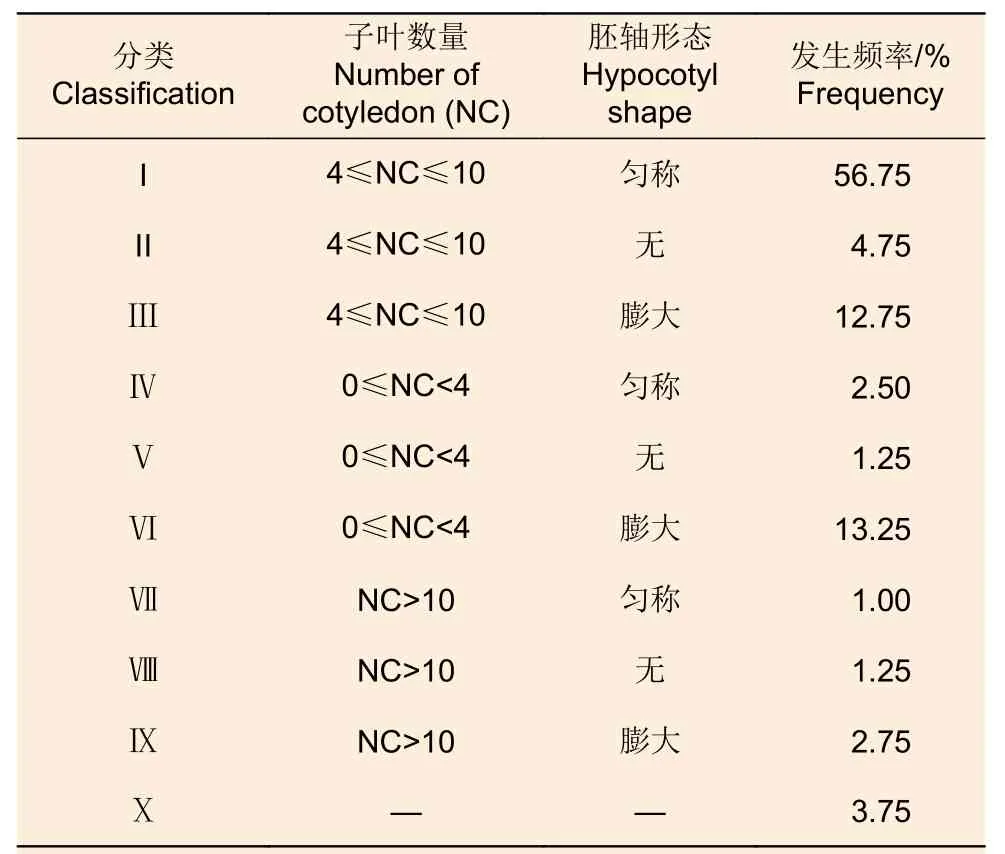
表1 日本落葉松胚狀體的形態和分類Table 1 Morphology and classification of Larix kaempferi embryoids

圖1 日本落葉松胚狀體的形態和分類Fig. 1 Morphology and classification of Larix kaempferi embryoids
2.2 未干化處理的胚狀體萌發情況統計分析
在萌發培養基上,隨機挑選的淡黃色胚狀體先在暗環境下培養3 d,之后在光環境下培養。7 d左右觀察到胚狀體發生膨大,子葉變綠。12 d 左右子葉變成翠綠色、胚軸伸長。隨后,部分胚狀體長出紅色根尖(圖2)。

圖2 不同類型日本落葉松胚狀體的萌發情況Fig. 2 Germination of different types of Larix kaempferi embryoids
類型Ⅰ胚狀體萌發培養5 d 后,子葉由黃變綠,胚狀體開始膨大;10 d 后子葉變成翠綠色,子葉和胚軸伸長,基部產生微小的紅色根尖(圖2A),萌發率為4.48%。類型Ⅹ的胚狀體在萌發培養過程中會慢慢褐化死亡。其它8 種類型的胚狀體都表現出子葉伸長,部分胚狀體有胚軸生長,但均未見胚根生長(表2,圖2B)。
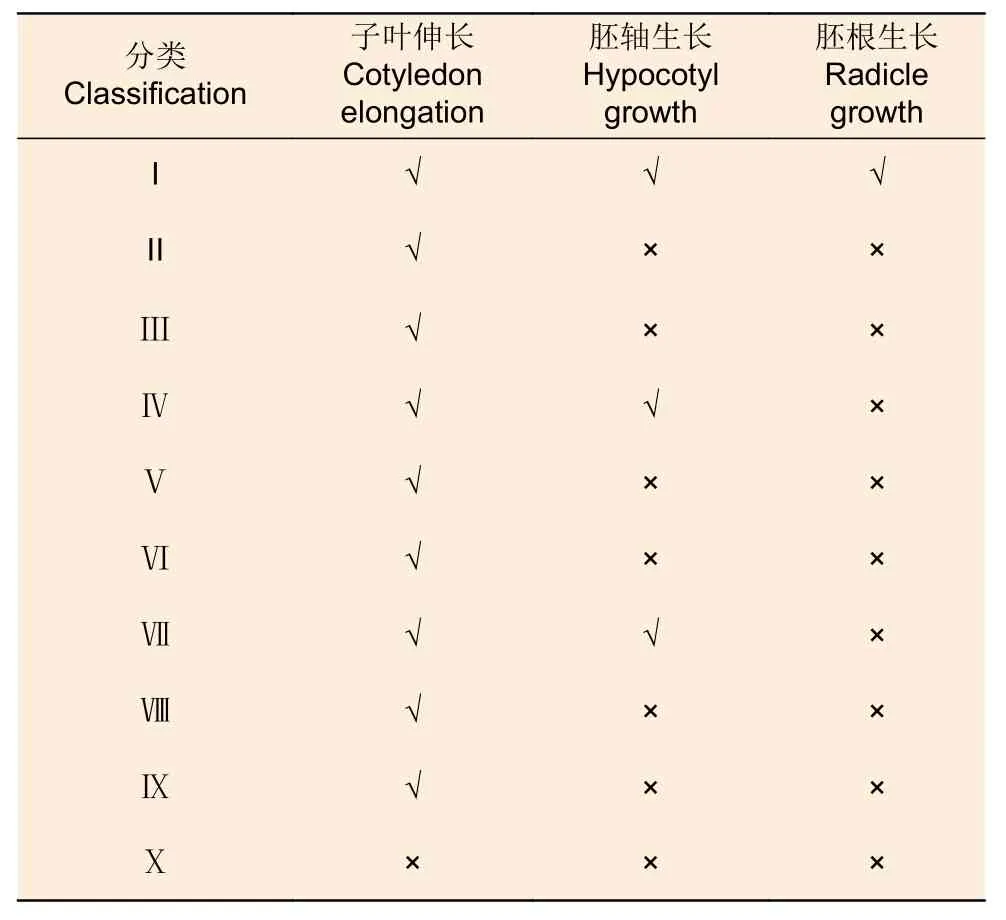
表2 日本落葉松胚狀體的萌發情況Table 2 Germination of Larix kaempferi embryoids
2.3 干化處理對胚狀體萌發的影響
在萌發培養過程中,僅類型Ⅰ胚狀體表現出完整的萌發性狀,因此用類型Ⅰ胚狀體進行干化處理。與未干化處理的胚狀體相比,干化處理7、14、21 和28 d 的胚狀體萌發率均有增加,其中干化處理14 d 的胚狀體萌發率最高,達到69.43%(表3,圖3),說明干化處理可以顯著促進胚狀體萌發。
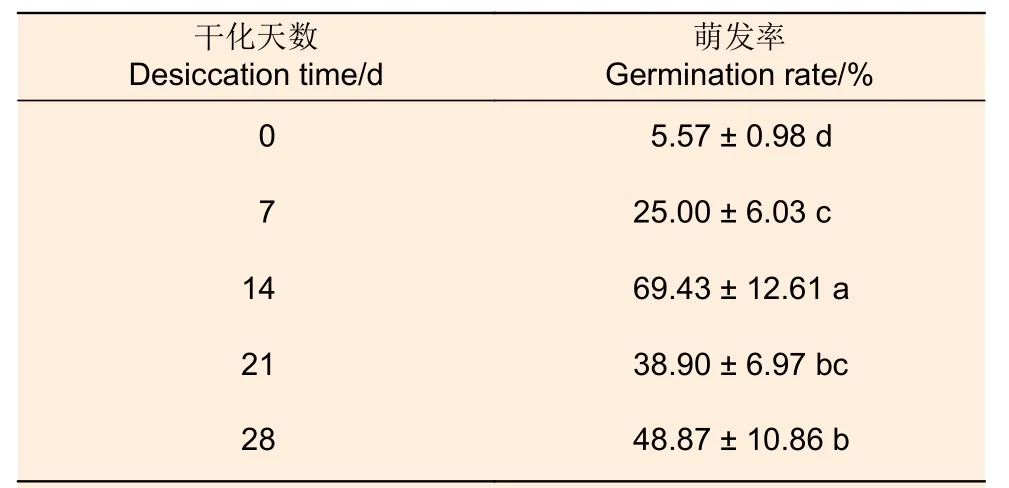
表3 干化對日本落葉松胚狀體萌發的影響Table 3 Effect of desiccation on the germination of Larix kaempferi embryoids
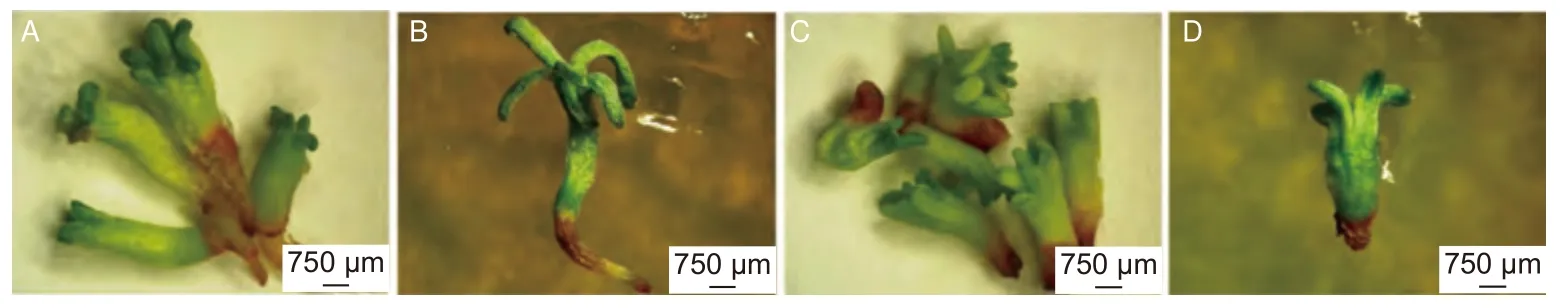
圖3 干化對日本落葉松胚狀體形態和萌發的影響Fig. 3 Effects of desiccation on morphology and germination of Larix kaempferi embryoids
2.4 胚狀體對干化處理的響應
在干化處理過程中,胚狀體的顏色有兩種變化,一種是子葉、胚軸變綠,胚根變紅(圖4A),另一種是子葉、胚軸顏色無明顯變化,胚根微微變紅(圖4B、C)。參考粗枝云杉胚狀體干化處理后的分類標準[24],將子葉和胚軸變綠、胚根變紅的日本落葉松胚狀體稱為有干化響應的胚狀體,將另外一種胚狀體稱為無干化響應的胚狀體。

圖4 日本落葉松胚狀體的干化響應Fig. 4 Desiccation response of Larix kaempferi embryoids
進一步分析發現,有干化響應的胚狀體更易萌發。所有經過干化處理的720 個胚狀體中,有干化響應的胚狀體共有649 個(90.1%),其中有317 個(48.8%)萌發;無干化響應的胚狀體共有71 個,但僅有11 個(15.5%)萌發。干化處理14 d,有干化響應的胚狀體162 個(90%),其中122 個(75.3%)萌發,而無干化響應的胚狀體18 個(10%),只有3 個(16.7%)萌發(表4)。因此,可以將干化響應作為挑選胚狀體進行萌發培養的依據。
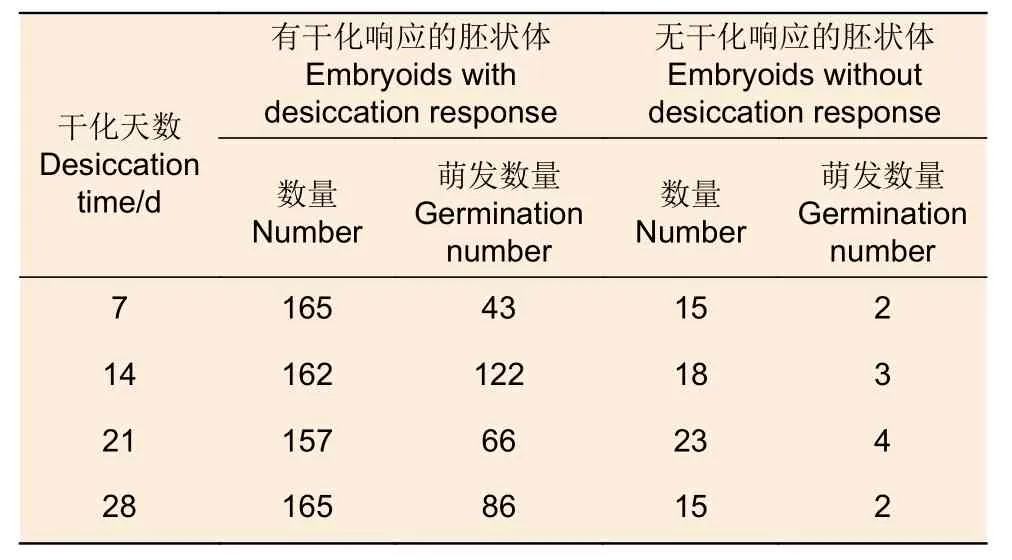
表4 干化響應與胚狀體萌發Table 4 Desiccation response and germination of somatic embryos
3 討論
在日本落葉松體細胞胚胎發生過程中,胚性愈傷組織經歷增殖培養和成熟培養后會產生形態各異的胚狀體,這些胚狀體的萌發能力存在差異。齊力旺[23]在研究華北落葉松過程中發現,子葉數量少、胚軸短的胚狀體都難以萌發。李哲馨等對日本落葉松胚狀體按照子葉數量進行了分類,發現子葉數量多于或等于4 個的胚狀體的萌發率高于其它類別的胚狀體[25]。上述工作表明,先對胚狀體進行分類而后進行萌發培養有助于提高萌發率和工作效率。本研究首先按照子葉數量和胚軸形態對日本落葉松胚狀體進行了更為細致的分類,發現它們可以分為10 種類型;隨后進行了萌發培養,發現僅有類型I 胚狀體可以萌發,萌發率為4.48%,其余類型胚狀體都難以萌發。以往對胚狀體進行分類多以子葉數量為依據[25-27],本研究以子葉數量和胚軸形態作為分類的依據,使分類標準更加細化與完善。
在萌發培養過程中,僅部分類型Ⅰ胚狀體表現出完整的萌發性狀,子葉伸長、變綠,胚軸伸長,胚狀體下端長出胚根,而其余類型胚狀體未長出胚根。推測部分類型Ⅰ胚狀體的根端發育相對完善,而其它類型胚狀體的根端發育異常。這些結果表明,胚狀體形態結構的完整性決定著它們是否可以萌發。研究結果也進一步說明在萌發培養前對胚狀體進行形態分類,有助于挑選出適宜進行萌發培養的胚狀體。
胚狀體“生理成熟”是萌發的關鍵前提,而干化處理可以使胚狀體達到生理成熟、促進其萌發[19-20]。Jing 等在云杉研究中發現,萌發率隨著干化時間呈現上升趨勢,干化處理14 d萌發率最高[28]。Maruyama等在日本黑松(Pinus thunberghiiParl.)和日本赤松(P.densifloraSiebold & Zucc.)研究中同樣發現,干化處理可以顯著提高萌發率[29]。本研究發現,未經干化處理的類型I 胚狀體萌發率為5.57%,而干化處理14 d 后萌發率提高了63.86%,說明干化處理可以顯著提高日本落葉松類型I 胚狀體的萌發率。由于未經干化處理的其余類型胚狀體萌發培養后未表現出完整的萌發性狀,本研究未對其進行干化處理。因此,干化處理是否可以使這些胚狀體達到生理成熟,進而促進其萌發,還需進一步確定。
即使是類型I 胚狀體,在干化處理后也表現出不同的性狀。其中子葉、胚軸變綠,胚根變紅的胚狀體,被稱為有干化響應的胚狀體;子葉、胚軸顏色無明顯變化,胚根微微變紅的胚狀體,被稱為未有干化響應的胚狀體。進一步研究發現,具有干化響應的類型I 胚狀體比無干化響應的類型I 胚狀體更易萌發,這與粗枝云杉胚狀體(Picea asperataMast)干化處理的研究結果一致[24]。據此,推測無干化響應的類型I 胚狀體根端發育不完善,而具有干化響應的類型I 胚狀體根端發育相對完善。由此可知,類型I 胚狀體內部結構也存在差異,且這些差異可以通過干化響應表現出來。因此對類型I 胚狀體進行干化處理,有助于進一步挑選更適宜進行萌發培養的胚狀體。
4 結論
基于上述研究結果,建議挑選子葉數量多于4、胚軸勻稱的日本落葉松胚狀體進行干化處理。干化處理14 d 的胚狀體萌發率提高了63.86%,且具有干化響應的胚狀體更易萌發。研究結果有助于促進體細胞胚胎發生技術在日本落葉松遺傳育種中的應用,也為其它針葉樹繁育技術的研發和完善提供參考。
猜你喜歡
西北民族大學學報(自然科學版)(2021年4期)2021-12-29 02:54:24
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
小聰仔(科普版)(2020年12期)2021-01-18 09:16:52
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
學生天地(2019年32期)2019-08-25 08:55:22
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
小天使·一年級語數英綜合(2017年11期)2017-12-05 18:49:56
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
