腸道菌群代謝產物與血管內皮功能的研究進展
2024-04-29 16:00:05呼瑞劉富強王軍奎
心血管病學進展 2024年2期
呼瑞 劉富強 王軍奎
【摘要】血管內皮功能對維持心血管系統穩態至關重要,其功能障礙是心血管疾病的重要原因。近年來,許多研究表明腸道菌群參與了血管內皮功能的改變,可能通過直接影響腸道菌群或間接影響腸道菌群代謝產物,如短鏈脂肪酸、次級膽汁酸、吲哚-3-甲醛、三甲胺-N-氧化物、苯乙酰谷氨酰胺、脂多糖、尿毒癥毒素等對血管內皮功能產生影響。現綜述近年來發現的腸道菌群代謝產物,為探索腸道菌群對血管內皮功能的影響提供理論基礎和新思路。
【關鍵詞】血管內皮功能;腸道菌群;代謝產物
【DOI】10.16806/j.cnki.issn.1004-3934.2024.02.000
Metabolites of Intestinal Flora and Vascular Endothelial Function
HU Rui1,LIU Fuqiang2,WANG Junkui2
Xian Medical University,Xian 710021,Shaanxi,China;2.Department of Cardiology,Shaanxi Provincial Peoples Hospital,Xian 710068,Shaanxi,China)
【Abstract】Vascular endothelial function is critical for maintaining homeostasis of the cardiovascular system,and its dysfunction is an important cause of cardiovascular disease. In recent years,many studies have shown that intestinal flora are involved in the alteration of vascular endothelial function,which may affect vascular endothelial function by directly influencing intestinal flora or indirectly influencing intestinal flora metabolites,such as short-chain fatty acids,secondary bile acids,indole-3-carboxaldehyde,trimethylamine-N-oxides,phenylacetylglutamine,lipopolysaccharides,and uremic toxins. The metabolites of intestinal flora discovered in recent years are reviewed to provide a theoretical basis and new ideas for exploring the effects of intestinal flora on vascular endothelial function.
【Keywords】Vascular endothelial function;Intestinal flora;Metabolite
健康的血管內皮是維系心血管系統穩態的基石,當血管內皮由生理狀態轉為功能障礙狀態被稱為血管內皮功能障礙。血管內皮功能障礙機制尚不完全明確,可能涉及到氧化應激、炎癥因子、低密度脂蛋白等多種因素[1]。近年來,大量研究發現,腸道菌群失調可能是導致血管內皮功能障礙的一個新機制,可通過腸道菌群和微生物代謝產物從腸道環境遷移至心血管系統代謝的相關組織中引起炎癥和氧化應激,導致血管內皮功能障礙[2]。因此,現綜述近年來新發現的腸道菌群代謝產物對血管內皮功能的影響,提出對腸道菌群代謝產物進行靶向干預可為血管內皮功能障礙的防治帶來希望。
1 ?血管內皮功能
血管內皮細胞是血漿和血管組織之間形成的一種特化上皮細胞,它不僅完成血漿和組織液的代謝交換,還能合成和分泌多種生物活性物質,保證血管的正常收縮和舒張功能,維持血管張力,調節血壓,平衡凝血和抗凝[3]。血管內皮功能障礙是一種以血管收縮和舒張機制失衡為特征的病理狀態,是動脈粥樣硬化、高血壓、肺動脈高壓、糖尿病、與神經退行性疾病相關的微血管病變、敗血癥等心血管疾病風險的標志[4-5]。已知的血管內皮細胞分泌的血管收縮因子包括內皮素-1、血栓素A2等,血管舒張因子包括一氧化氮(nitric oxide,NO)、前列環素、內皮衍生超級化因子等,其中作用最重要的是NO。NO是一種可溶性氣體,具有重要的血管舒張功能,內皮型一氧化氮合酶(endothelial nitric oxide synthase,eNOS)利用煙酰胺腺嘌呤二核苷酸提供的電子催化L-精氨酸和O2,生成NO和L-瓜氨酸,NO主要由內皮細胞受剪切應力作用而釋放,當NO釋放到血管平滑肌細胞,可激活鳥苷酸環化酶直接導致血管擴張。除此之外,NO在血管中還具有多種作用,如通過抑制血小板聚集和黏附、纖溶酶原激活物抑制物-1發揮抗血栓作用,抑制白細胞的聚集發揮抗炎作用[6-7]。血管內皮功能障礙與NO產生不足和將生物可利用的NO轉化為其他物質的消耗過多密切相關,最終導致血管穩態失衡,導致促炎狀態和血栓形成[7]。
2 ?腸道菌群的概述
腸道微生物是由細菌、古細菌、真菌、原生動物和病毒組成的生態群落[8],其中細菌占99%以上,故以腸道菌群代指腸道微生物[9]。腸道菌群大約是由1014個細菌組成的高度多樣化的群落[10],其中最主要的6大門是厚壁菌門、擬桿菌門、變形菌門、放線菌門、梭桿菌門和疣微球菌門,在健康菌群中,厚壁菌門和擬桿菌門是主要優勢菌群,占總數的80%~90%[11]。腸道菌群產生大量具有生物學效應的代謝產物,如三甲胺N-氧化物(trimethylamine-N-oxide,TMAO)、苯乙酰谷氨酰胺(phenylacetylglutamine,PAGln)、尿毒癥毒素、NO、脂多糖(lipopolysaccharide,LPS)、短鏈脂肪酸(short-chain fatty acid,SCFA)、膽汁酸、維生素K、維生素B復合物、腸道激素及神經遞質等[12],參與人體各種功能的調節,包括為宿主提供營養,參與生長代謝和免疫調節,清除致病微生物,維持腸道上皮的完整性和穩態。當不合理的飲食模式、環境因素、腸道感染或抗生素的使用導致腸道微生物的種類和數量發生改變時,就會發生腸道菌群失調,引起炎癥和代謝的紊亂[13]。研究[5]發現,腸道菌群失調不僅會導致炎癥性腸病和結腸癌等腸道相關性疾病,還可導致肥胖、糖尿病、過敏,尤其是心血管疾病。Cui等[14]納入29例冠心病患者和35例健康人作為對照組,從糞便中提取腸道菌群DNA,行高通量測序發現冠心病患者擬桿菌門和變形菌門減少,厚壁菌門和梭桿菌門相對豐度增加,厚壁菌門/擬桿菌門比例顯著上升,且兩組間的腸道菌群組成存在顯著差異,提示腸道菌群改變可能與冠心病發生有關。
3 ?腸道菌群與血管內皮功能
3.1 ?腸道菌群改變對血管內皮功能的直接影響
當腸道益生菌數量減少或致病菌數量增加時就會發生腸道菌群失調,使得腸道菌群與宿主之間的相互作用受到影響,腸道菌群失調可通過直接影響機體糖脂代謝或間接改變腸道通透性影響機體循環中炎癥反應和氧化應激水平,從而導致血管內皮功能障礙的發生發展[15]。血管內皮功能障礙是心血管疾病發病的關鍵環節,與冠狀動脈疾病、高血壓、慢性心力衰竭等心血管疾病密切相關。在一項臨床研究中,Tsutsumi等[16]招募60例健康受試者,隨機分配到長鏈單不飽和脂肪酸(刀魚油)組和對照(橄欖油)組,每日攝入4.2?g油(12粒膠囊),持續干預4周后腸道菌群組成發生改變,結果表明刀魚油組受試者血流介導的血管擴張(flow-mediated?dilation,FMD)顯著增加4.2%,血管內皮功能得到改善。在動物研究[16]中,將ApoE-/-小鼠隨機分為3組,分別是烯油酸(C20∶1)組、鯨油酸(C22∶1)組、對照(橄欖油)組,持續喂養12周,收集小鼠糞便樣本,行16S rRNA基因測序發現,經過C20∶1、C22∶1處理后的小鼠,阿克曼氏菌的豐度增加、擬桿菌門比例增加、厚壁菌門比例顯著降低、厚壁菌門/擬桿菌門的比例降低,小鼠血管內皮功能改善,減輕了動脈粥樣硬化。Malik等[17]研究發現冠心病患者每天服用植物乳桿菌299v,6周后可改善心臟血管內皮功能。以上研究表明,當腸道菌群組成及比例發生改變,益生菌比例增多時可使血管內皮功能得到改善。
3.2??腸道菌群代謝產物對血管內皮功能的影響
3.2.1 ?SCFA
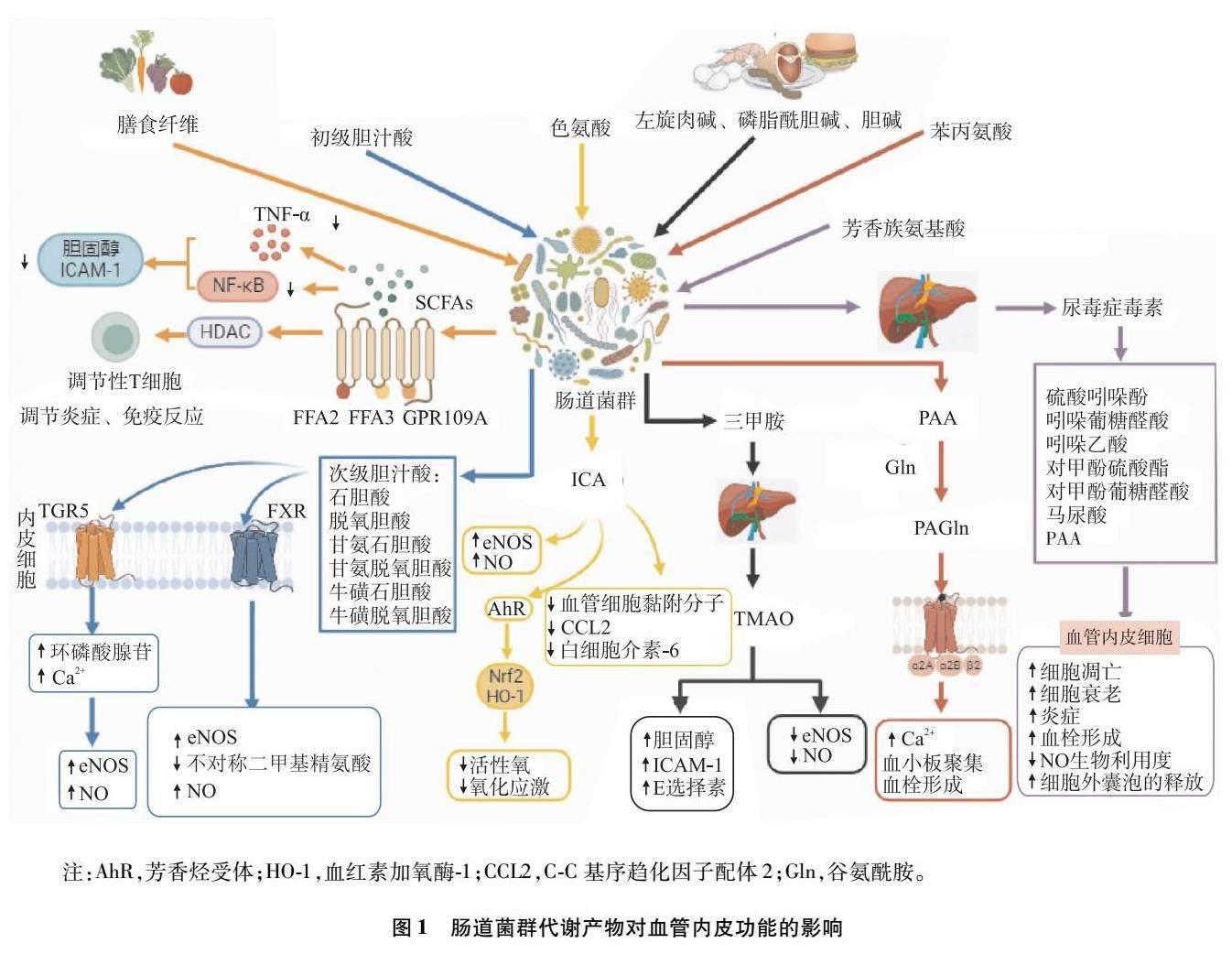
SCFA是由結腸內的腸道厭氧菌通過發酵難以消化的膳食纖維而產生的碳原子少于6個的脂肪酸,包括含量豐富的乙酸、丙酸和丁酸,以及含量較少的戊酸、己酸,擬桿菌門、厚壁菌門、毛螺菌科、乳酸桿菌、糞桿菌和瘤胃球菌主要參與SCFA的生成[5,18]。SCFA通過維持腸道穩態、減少炎癥、調節血管功能在人體健康中發揮重要作用。研究[18-20]表明,SCFA不僅可通過激活G蛋白偶聯受體(G-protein?coupled receptor,GPR)[如游離脂肪酸(free fatty acid,FFA)2、FFA3、GPR109A]和抑制組蛋白去乙酰化酶(histone deacetylase,HDAC)來誘導調節T細胞的分化,調節炎癥及免疫反應,通過與FFA2受體、FFA3受體及嗅覺受體結合參與血壓的調節,還可通過抑制腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)、核因子κB(nuclear factor-κB,NF-κB)信號通路來降低膽固醇水平和細胞間黏附分子(intercellular adhesion molecule,ICAM)表達,調節免疫應答,改善血管內皮功能,以此減緩動脈粥樣硬化的發展。Li等[19]研究發現,SCFA,特別是丁酸作為HDAC的抑制劑具有調節血管內皮功能、抑制炎癥因子和氧化應激的作用,可保護血管內皮和改善動脈粥樣硬化。結合上述證據,外源性補充SCFA物質可能成為改善血管內皮功能和動脈粥硬化的新靶點。
3.2.2 ?次級膽汁酸
肝細胞合成膽汁酸,通過膽道系統釋放入腸道中,在腸道菌群酶的催化下轉化為次級膽汁酸,游離的次級膽汁酸具有激素活性和多效性,它們通過激活武田G蛋白偶聯受體(Takeda G-protein?coupled?receptor 5,TGR5)、法尼醇X受體(farnesoid X receptor,FXR)發揮多種作用,如調節糖脂代謝、維持腸道屏障的完整性、抑制炎癥因子的釋放[21]。研究[22]發現,次級膽汁酸與TGR5結合觸發肝竇內皮細胞中環磷酸腺苷表達,細胞內Ca2+水平升高,導致eNOS磷酸化的激活和NO的釋放,NO不僅可舒張血管和增加血流,還具有抗炎、減少血管內皮細胞黏附的作用,從而改善血管內皮功能。有研究[15]報道,牛磺酸熊去氧膽酸(taurine deoxycholic acid,TUDCA)通過激活FXR可增加eNOS表達,并可能減少不對稱二甲基精氨酸形成;通過激活TGR5可增加內皮細胞內Ca2+水平、eNOS和絲氨酸/蘇氨酸激酶磷酸化;通過促進硫化氫的生成來調控NO的合成和活性,從而促進血管舒張,改善血管內皮功能。Walsh等[23]納入12例健康的年輕受試者,隨機分為TUDCA組和安慰劑組,分別給予TUDCA(1?500 mg)和安慰劑(1?500 mg),8?h后測量肱動脈FMD作為基線,均給予75?g葡萄糖刺激后60?min和120?min重復測量肱動脈FMD,發現安慰劑組FMD較基線水平明顯降低,而TUDCA組變化不明顯,由此可得,短期內口服TUDCA(15?00?mg)可改善高血糖誘導的血管內皮功能障礙。
3.2.3??吲哚-3-甲醛
吲哚-3-甲醛(indole-3-carboxaldehyde,ICA)是一種腸道微生物來源的色氨酸代謝產物[24],Lu等[25]通過宏基因組學研究發現,高脂肪飲食組擬桿菌科和乳酸桿菌科的豐度低于對照組,而這些細菌已被證實可產生色氨酸酶,從而將色氨酸代謝成ICA。既往研究[26]證實ICA具有增強腸道上皮屏障和抗炎活性的作用。但最新研究發現了ICA對內皮細胞的作用。Lu等[25]研究發現,ICA雖不影響ICAM和內皮素的RNA水平,但卻增加了eNOS的轉錄水平,從而增加了NO的生成。ICA還降低了血管細胞黏附分子、C-C基序趨化因子配體2以及白細胞介素-6的基因表達。此外,ICA含有芳香烴結構,通過芳香烴受體激活后,芳香烴受體進入細胞核與核轉錄因子紅系2相關因子2(nuclear factor-erythroid 2-related factor 2,Nrf2)啟動子區域結合從而上調Nrf2-血紅素加氧酶-1表達,降低細胞內氧化應激和低密度脂蛋白誘導的活性氧水平[25,27]。總之,最新研究表明ICA通過增加eNOS轉錄,降低炎癥因子和活性氧水平,保護血管內皮細胞,緩解動脈粥樣硬化的發展。
3.2.4 ?TMAO
富含左旋肉堿、磷脂酰膽堿(卵磷脂)、甜菜堿和膽堿等動物性食品通過腸道菌群代謝產生三甲胺,三甲胺被肝臟中的黃素單氧化酶迅速氧化為TMAO[13,28],TMAO是一種具有生物活性的分子。研究[4]發現,無論是在人臍靜脈內皮細胞(human umbilical vein endothelial cell,HUVEC)還是牛主動脈內皮細胞,TMAO均可減少eNOS的合成,減少NO生成,從而導致血管內皮功能改變,但其機制可能不盡相同。在HUVEC模型中,TMAO通過氧化應激和硫氧還蛋白相互作用蛋白、NOD樣受體熱蛋白結構域相關蛋白3炎性小體活化來抑制eNOS的合成和NO釋放,而在牛主動脈內皮細胞模型中,TMAO影響嘌呤誘導的細胞內Ca2+增加、eNOS磷酸化和NO釋放。活化的內皮細胞上表達了不同的黏附分子,這些黏附分子可直接誘導血管內皮功能障礙和動脈粥樣硬化。研究[4]發現,用TMAO處理的人主動脈內皮細胞表達更高水平的ICAM-1和E選擇素,誘導血管內皮功能障礙。TMAO亦可使膽固醇7α-羥化酶的表達降低,抑制膽固醇轉運,從而使細胞中膽固醇水平升高、泡沫細胞增多[29]。Brunt等[29]研究發現,在健康人群中,中老年人的TMAO水平高于年輕人,且與肱動脈FMD呈負相關(P<0.000?01),TMAO水平的升高是血管內皮功能障礙的上游驅動因素,且TMAO與心血管疾病風險預測成正相關。
3.2.5 ?PAGln
PAGln是腸道菌群分解必需氨基酸苯丙氨酸的代謝產物,苯丙氨酸存在于多種植物和動物來源的蛋白中,未被吸收的苯丙氨酸可被腸道微生物代謝形成苯丙酮酸和苯乙酸(phenylacetic acid,PAA),PAA可與谷氨酰胺結合生成PAGln,PAA與甘氨酸結合生成苯乙酰甘氨酸。在人血、分離的血小板及頸動脈損傷小鼠模型中發現,PAGln可通過GPR(包括α2A、α2B和β2腎上腺素能受體)導致血小板活性和聚集增強,促進Ca2+水平升高,加速了血小板的聚集和血栓形成,損傷血管內皮功能,最終促進動脈粥樣硬化發生發展[30]。Nemet等[30]對臨床隊列人群(n=1?162)的血漿進行靶向和非靶向代謝組學分析后發現,PAGln在機制和臨床方面與主要不良心血管事件(major adverse cardiovascular event,MACE)相關,利用液相質譜靶向檢測方法評估了4?000例擇期進行心臟評估的穩定受試者血漿,發現患有MACE的患者血漿PAGln水平較非MACE患者水平高(P<0.000?1),且高PAGln水平的患者具有更大患MACE的風險(P<0.000?1)。目前體內外試驗證實PAGln可增強血小板活化和血栓形成,損傷血管內皮功能,導致MACE風險增加。
3.2.6??LPS
LPS也稱內毒素,是由脂質和多糖合成的復合物,是革蘭氏陰性菌細胞壁的一種特有成分,腸道菌群失調可導致腸道通透性增加,促進LPS從腸腔易位進入循環,可誘導全身炎癥和細胞凋亡[15]。研究[31]表明,LPS可通過Toll樣受體4(Toll-like receptor 4,TLR4)/髓樣分化因子88/NF-κB通路誘導白細胞介素-1、白細胞介素-6和TNF-α的釋放,引發慢性炎癥,損害血管內皮功能,促進心血管疾病的發生發展。Zhao等[32]用不同濃度的LPS(0、0.5、1、2和4 μg/mL)處理HUVEC?24?h后發現,乳酸脫氫酶、白細胞介素-1β、TLR4、NOD樣受體熱蛋白結構域相關蛋白3的表達呈濃度依賴性增加,誘導HUVEC焦亡,并發現LPS可通過激活特異性蛋白1/網鈣結合蛋白-2 /eNOS信號通路誘導血管內皮焦亡,影響其功能。研究[15]發現,當腸道屏障受損時,LPS進入血液,與TLR4結合后,LPS促進血小板中P選擇素、白細胞介素-1β的表達,這會引起肺血管周圍炎癥反應的發生及原位血栓的形成,從而導致肺動脈高壓的形成。上述證據表明,腸道菌群失調使LPS進入循環,從各個機制導致全身炎癥和細胞凋亡,從而使血管內皮功能受損,導致心血管疾病的發生發展。
3.2.7??尿毒癥毒素
尿毒癥毒素是由腸道菌群代謝氨基酸衍生的代謝產物,蛋白質中的芳香族氨基酸(酪氨酸、苯丙氨酸和色氨酸)可被腸道菌群及肝臟代謝為毒素,如硫酸吲哚酚、吲哚葡萄糖醛酸、吲哚乙酸、對甲酚硫酸酯、對甲酚葡萄糖醛酸、馬尿酸和PAA。研究[33]發現,硫酸吲哚酚可通過誘導血管內皮細胞凋亡、衰老、炎癥、血栓形成以及降低NO生物利用度、減少細胞外囊泡的釋放來損傷血管內皮。在內皮細胞中,硫酸吲哚酚、對甲酚葡糖醛酸可激活NF-κB信號通路,上調ICAM-1和單核細胞趨化蛋白-1表達,從而促進血管內皮功能障礙和動脈粥樣硬化[5,34]。研究[33]表明,將硫酸吲哚酚作用于正常小鼠和大鼠的胸主動脈,發現硫酸吲哚酚可降低乙酰膽堿誘導的內皮依賴性血管舒張,這種作用可能歸因于NO生物利用度降低。Omori等[35]納入176例從未發生過心血管疾病的2型糖尿病患者和40例冠狀動脈疾病患者,通過對患者的血漿進行非靶向代謝組學分析發現硫酸吲哚酚與頸動脈最大內膜中層厚度呈正相關,與FMD成負相關(P<0.05)。上述證據表明,尿毒癥毒素可通過多種途徑促進血管內皮功能障礙,調控尿毒癥毒素可能是一種潛在的防治血管內皮功能障礙及動脈粥樣硬化的方法。見圖1。

注:AhR,芳香烴受體;HO-1,血紅素加氧酶-1;CCL2,C-C基序趨化因子配體2;Gln,谷氨酰胺。
圖1 ?腸道菌群代謝產物對血管內皮功能的影響
4 ?結論與展望
近年來,腸道菌群代謝產物被發現參與多種人類疾病的發生發展,根據目前所有研究,可認為腸道菌群代謝產物是血管內皮功能障礙發生發展的重要環節。值得關注的是,由于腸道菌群數量大、種類多、長期動態變化且易受各種飲食習慣及環境因素的影響,導致建模困難,臨床前研究遠遠不夠,腸道菌群在分子水平上影響血管內皮功能的具體作用機制還無法明確闡述。因此,接下來要對腸道菌群及其代謝產物與血管內皮功能障礙的關系深入探索,并對腸道菌群及其代謝產物進行精準的靶向干預,這將為血管內皮功能障礙的預防及治療帶來新希望。
參考文獻
[1] Xu S,Ilyas I,Little PJ,et al. Endothelial dysfunction in atherosclerotic cardiovascular diseases and beyond:from mechanism to pharmacotherapies[J]. Pharmacol Rev,2021,73(3):924-967.
[2] Battson ML,Lee DM,Jarrell DK,et al. Suppression of gut dysbiosis reverses Western diet-induced vascular dysfunction[J]. Am J Physiol Endocrinol Metab,2018,314(5):E468-E477.
[3] Zhang Q,Liu J,Duan H,et al. Activation of Nrf2/HO-1 signaling:an important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress[J]. J Adv Res,2021,34:43-63.
[4] Querio G,Antoniotti S,Geddo F,et al. Modulation of endothelial function by TMAO,a gut microbiota-derived metabolite[J]. Int J Mol Sci,2023,24(6):5806.
[5] Amedei A,Morbidelli L.?Circulating metabolites originating from gut microbiota control endothelial cell function[J]. Molecules,2019,24(21):3992.
[6] Vanhoutte PM,Shimokawa H,Feletou M,et al. Endothelial dysfunction and vascular disease—A?30th anniversary update[J]. Acta Physiol (Oxf),2017,219(1):22-96.
[7] Tenopoulou M,Doulias P-T.?Endothelial nitric oxide synthase-derived nitric oxide in the regulation of metabolism[J]. F1000Res,2020,9:F1000 Faculty Rev-1190.
[8] Barko PC,Mcmichael MA,Swanson KS,et al. The gastrointestinal microbiome:a review[J]. J Vet Intern Med,2018,32(1):9-25.
[9] Eckburg PB,Bik EM,Bernstein CN,et al. Diversity of the human intestinal microbial flora[J]. Science,2005,308(5728):1635-1638.
[10] Maiuolo J,Carresi C,Gliozzi M,et al. The contribution of gut microbiota and endothelial dysfunction in the development of arterial hypertension in animal models and in humans[J]. Int J Mol Sci,2022,23(7):3698.
[11] Cortés-Martín A,Iglesias-Aguirre CE,Meoro A,et al. There is no distinctive gut microbiota signature in the metabolic syndrome:contribution of cardiovascular disease risk factors and associated medication[J]. Microorganisms,2020,8(3):416.
[12] Kazemian N,Mahmoudi M,Halperin F,et al. Gut microbiota and cardiovascular disease:opportunities and challenges[J]. Microbiome,2020,8(1):36.
[13] Jin M,Qian Z,Yin J,et al. The role of intestinal microbiota in cardiovascular disease[J]. J Cell Mol Med,2019,23(4):2343-2350.
[14] Cui L,Zhao T,Hu H,et al. Association study of gut flora in coronary heart disease through high-throughput sequencing[J]. Biomed Res Int,2017,2017:3796359.
[15] Chen YH,Yuan W,Meng LK,et al. The role and mechanism of gut microbiota in pulmonary arterial hypertension[J]. Nutrients,2022,14(20):4278.
[16] Tsutsumi R,Yamasaki Y,Takeo J,et al. Long-chain monounsaturated fatty acids improve endothelial function with altering microbial flora[J]. Transl Res,2021,237:16-30.
[17] Malik M,Suboc TM,Tyagi S,et al. Lactobacillus plantarum 299v supplementation improves vascular endothelial function and reduces inflammatory biomarkers in men with stable coronary artery disease[J]. Circ Res,2018,123(9):1091-1102.
[18] Hemmati M,Kashanipoor S,Mazaheri P,et al. Importance of gut microbiota metabolites in the development of cardiovascular diseases (CVD)[J]. Life Sci,2023,329:121947.
[19] Li M,van Esch BCAM,Wagenaar GTM,et al. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells[J]. Eur J Pharmacol,2018,831:52-59.
[20] Brown JM,Hazen SL. Microbial modulation of cardiovascular disease[J]. Nat Rev Microbiol,2018,16(3):171-181.
[21] Gou X,Qin L,Wu D,et al. Research progress of Takeda G protein-coupled receptor 5 in metabolic syndrome[J]. Molecules,2023,28(15):5870.
[22] Guizoni DM,Vettorazzi JF,Carneiro EM,et al. Modulation of endothelium-derived nitric oxide production and activity by taurine and taurine-conjugated bile acids[J]. Nitric Oxide,2020,94:48-53.
[23] Walsh LK,Restaino RM,Neuringer M,et al. Administration of tauroursodeoxycholic acid prevents endothelial dysfunction caused by an oral glucose load[J]. Clin Sci (Lond),2016,130(21):1881-1888.
[24] Paeslack N,Mimmler M,Becker S,et al. Microbiota-derived tryptophan metabolites in vascular inflammation and cardiovascular disease[J]. Amino Acids,2022,54(10):1339-1356.
[25] Lu Y,Yang W,Qi Z,et al. Gut microbe-derived metabolite indole-3-carboxaldehyde alleviates atherosclerosis[J]. Signal Transduct Target Ther,2023,8(1):378.
[26] Scott SA,Fu J,Chang PV,et al. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor[J]. Proc Natl Acad Sci U S A,2020,117(32):19376-19387.
[27] Nguyen C,Edgley AJ,Kelly DJ,et al. Aryl hydrocarbon receptor inhibition restores indoxyl sulfate-mediated endothelial dysfunction in rat aortic rings[J]. Toxins (Basel),2022,14(2):100.
[28] Liu Y,Dai M.?Trimethylamine N-oxide generated by the gut microbiota is associated with vascular inflammation:new insights into atherosclerosis[J]. Mediators Inflamm,2020,2020:4634172.
[29] Brunt VE,Gioscia-Ryan RA,Casso AG,et al. Trimethylamine-N-oxide?promotes age-related vascular oxidative stress and endothelial dysfunction in mice and healthy humans[J]. Hypertension,2020,76(1):101-112.
[30] Nemet I,Saha PP,Gupta N,et al. A cardiovascular disease-linked gut microbial metabolite acts via adrenergic receptors[J]. Cell,2020,180(5):862-877.
[31] Chistiakov DA,Bobryshev YV,Kozarov E,et al. Role of gut microbiota in the modulation of atherosclerosis-associated immune response[J]. Front Microbiol,2015,6:671.
[32]? Zhao J,Liu Z,Chang Z.?Lipopolysaccharide induces vascular endothelial cell pyroptosis via the SP1/RCN2/ROS signaling pathway[J]. Eur J Cell Biol,2021,100(4):151164.
[33] Matsumoto T,Kojima M,Takayanagi K,et al. Role of S-equol,indoxyl sulfate,and trimethylamine N-oxide?on vascular function[J]. Am J Hypertens,2020,33(9):793-803.
[34] Kumar T,Dutta RR,Velagala VR,et al. Analyzing the complicated connection between intestinal microbiota and cardiovascular diseases[J]. Cureus,2022,14(8):e28165.
[35] ???Omori K,Katakami N,Arakawa S,et al. Identification of plasma inositol and indoxyl sulfate as novel biomarker candidates for atherosclerosis in patients with type 2 diabetes—Findings from metabolome analysis using GC/MS[J]. J Atheroscler Thromb,?2020,?27(10)?:1053-1067.
收稿日期:2023-09-24