氮添加對森林土壤有機碳庫固存及CO2排放的影響研究進展
2024-05-06 03:19:40蘇立城陳曉珊羅志忠陳煜佳吳道銘曾曙才
生態學報 2024年7期
蘇立城,陳曉珊,羅志忠,胡 英,陳煜佳,吳道銘,曾曙才
華南農業大學林學與風景園林學院,廣州 510642
森林生態系統是陸地生態系統中最大的碳庫,森林土壤則是陸地生態系統中最大的有機碳庫,據權威研究估算,全球有約 50% 的地上碳和 40% 的地下碳儲存在森林生態系統中,目前全球森林生態系統的碳儲量達到了 (861±66) Gt,其中森林土壤的碳儲量約為 (383±30) Gt[1]。森林生態系統不僅是溫室氣體的“源”,也是溫室氣體的“匯”,森林土壤呼吸作用所釋放的二氧化碳(CO2)是溫室氣體的重要來源之一,且極易受人類活動的干擾[2],近年來,由于溫室效應加劇,圍繞如何提高森林土壤有機碳庫固存能力并減緩CO2排放的研究受到了廣泛關注。
氮是限制植物生長和土壤微生物生命活動的主要營養元素,外源氮的添加對提高森林凈初級生產力、促進森林可持續發展而言具有重要意義。但在森林經營過程中,氮沉降或施肥措施引起的大量氮素攜入可能會改變土壤養分供應和理化性質穩態,導致土壤酸化,從而增加土壤微生物生命活動的環境壓力并改變其群落結構特征和生態功能[3],具體表現為影響輸入有機質(如凋落物、細根周轉和根系分泌物等)的降解、固定和土壤有機碳(SOC)的礦化過程,最終影響森林土壤的碳循環過程并改變土壤有機碳庫的固存和CO2排放通量[4—5]。由于森林土壤中的碳、氮循環過程存在緊密的耦合作用,因此氮也是影響森林土壤碳循環過程的重要因素之一,即使森林土壤中的氮含量發生微小變化,也可能會破壞森林土壤的碳氮平衡,繼而影響有機碳庫的穩定性和改變CO2的排放通量,對全球氣候變化產生重大影響[6]。因此,明確森林土壤碳循環過程對氮添加的響應,對預測全球氣候變化條件下森林土壤有機碳庫穩定性及尋求降低CO2排放通量的方法而言具有重要的指導意義。
1 森林生態系統的土壤碳循環過程
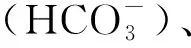
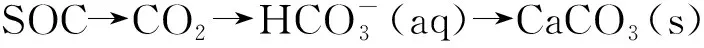
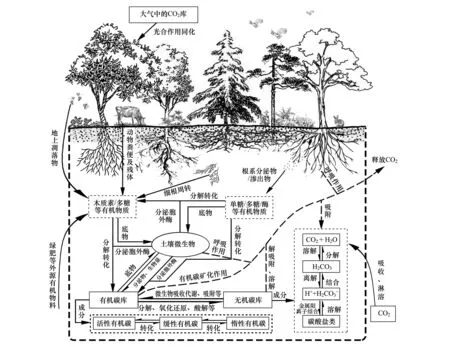
圖1 森林生態系統中的土壤碳循環過程Fig.1 Schematic overview of soil carbon cycling in forest ecosystem
相關Meta分析表明[15—18],森林土壤的碳循環過程極其復雜,不僅與其他元素如氮、磷循環過程相互耦合,同時還受到氣候、森林經營、人為活動、土壤理化環境以及土壤微生物、相關酶活性等眾多因素的共同影響。
2 氮添加對森林土壤有機碳庫固存的影響
2.1 氮添加對森林土壤有機碳輸入的影響
氮是植物生長的主要限制性營養元素,不僅是植物體內蛋白質和葉綠素的主要成分,也是核酸、各類酶、磷脂和某些生長激素的組成成分,有“生命元素”之稱,可促進或抑制植物的生長固碳[19]。促進作用表現為當氮適量時,可促進植物體合成蛋白質、提高相關酶活性來增加葉面積和葉綠素含量,進而提高光合作用速率和碳同化能力[20];抑制作用表現為當氮添加不足或過量時,造成植物體內營養失衡,抑制植物葉片形態結構和生理功能,導致植物光合作用過程受限,降低碳同化速率[21]。因此,氮添加可通過促進或抑制植物光合作用來影響植物的生長發育和有機物積累,繼而改變植物的地上凋落物產量、化學組成和地下碳分配格局來影響有機物質的回歸,并調節土壤微生物的相關特性來影響SOC輸入[22]。即氮添加對SOC輸入的影響主要可表現為兩個方面:一是通過影響植物生長固碳后地上凋落物的相關特性和微生物的分解速率來影響SOC輸入[23—24];二是通過影響植物的地下碳分配格局來調節細根周轉速率和改變根系分泌物、滲出物的產生來影響SOC輸入[25—27]。
第一方面,氮添加可能會改變凋落物的產量和化學組成,同時通過影響土壤微生物群落特征來調節凋落物的分解速率,由此影響凋落物釋放SOC[28—29]。研究表明,氮添加會改變森林土壤的養分供應差異來影響植物的生長和碳同化,直接促進或抑制凋落物的產生并調節凋落物的數量和化學組成(C/N、木質素/N等),這會導致凋落物的可分解性(難易程度)和分解速率改變[30—31];其次,氮添加引起的森林土壤理化性質改變和養分供應差異也會影響土壤微生物的群落組成、活性和功能多樣性,進而促進或抑制凋落物分解腐化為SOC組分[32—33]。
氮的添加會促進森林生態系統的凈初級生產力[34]。其中,當氮添加量較低時,凋落物產量有所提高且基質的C/N和木質素/N呈下降趨勢,致使其所含的耐分解化合物成分減少,同時提高了相關微生物和酶的活性,促進凋落物分解腐化為SOM,由此增加SOC輸入[35—36]。而當氮添加量較高時,盡管凋落物的產量和基質成分與較低量氮添加時相比無明顯差異,但由于土壤微生物和酶活性的下降導致其分解腐化凋落物的速率明顯受限,對SOC輸入的貢獻則表現為無顯著影響或略有上升[37]。Hobbie等人[38]發現,適量氮添加提高了森林的凋落物數量,并使凋落物中木質素含量顯著下降,其分解腐化速率顯著提高。Carly等[39]的研究則證實凋落物數量的增加可以提高森林土壤中己糖和酚酸含量,刺激了土壤微生物和酶的活性而提高凋落物的分解速率。Thomas等[40]和Xia等[41]也發現,在低氮添加水平下(30 kg N hm-2a-1),森林土壤中的木質素分解酶類活性顯著降低,有效減緩了腐殖質的分解速率,增加了土壤中難分解有機碳的積累。而付琦等[42]通過梯度氮添加試驗發現,施氮增加了森林的凋落物數量和N含量,即降低了基質的C/N,促進SOC積累,但隨著施氮量增加,年SOC輸入量呈下降趨勢。Saiya-Cork等[43]的研究也表明,低氮添加量(30 kg N hm-2a-1)使森林土壤中的纖維素酶、脲酶、酸性磷酸酶和糖苷酶等活性顯著升高,顯著促進凋落物分解為SOM,但是隨著氮添加量的增加,相關酶活性顯著降低,SOM量下降。郭綺雯等[44]的梯度氮添加研究表明,中低濃度的氮添加顯著促進了凋落物中纖維素和半纖維素等易分解組分的分解,并釋放SOC,而高氮添加(300 kg N hm-2a-1)下則表現為抑制作用。周世興等[45]的研究則發現,氮添加會抑制凋落物中木質素和纖維素等難分解碳組分的分解,降低SOC的輸入,且抑制作用隨氮添加量的增加而顯著加強。
第二方面,氮添加通過改變根際土壤的理化性質或養分有效性來調節根系生命活動,此過程中,由于根系的生理更新作用,不斷產生的細根一部分會替代老根,其余的則會和被替代的老根一起凋亡分解,該過程稱為細根周轉,細根周轉產物即根系凋落物[46];同時,根系在生命活動過程中也會向土壤釋放出有機物質,包括碳水化合物、氨基酸、有機酸、脂肪酸、肽、核苷和酶類等,稱為根系分泌物和根系滲出物[47]。氮添加通過調節根際養分有效性可影響細根周轉速率以及根系的生物量和化學組成,或改變根際土壤的理化性質,使根系響應繼而主動或被動向土壤釋放有機物質(分泌物或滲出物)[48]。根系凋落物和根系分泌物、滲出物都屬于有機物質,是森林土壤有機碳庫的重要來源之一。
細根周轉途徑輸入SOC的過程與細根產量、周轉速率以及分解速率密切相關。研究表明,適量氮添加會提高森林植物的細根產量和周轉速率而增加根系凋落物的產量,同時會降低根系凋落物的C/N和木質素/N;且隨著氮添加量增多,傾向于提高根際胞外酶中纖維素酶活性并抑制木質素酶活性[49]。
一項針對全球不同森林生態系統細根周轉對氮添加響應的薈萃分析就表明,氮添加使森林植物細根產量平均增加了 27%,細根周轉速率及土壤碳儲量均有效提高[50]。Li等[51]研究發現,當氮添加量不超過 50 kg N hm-2a-1時,森林的細根產量和周轉速率隨氮添加量的增加而提高,且土壤碳儲量與細根產量之間存在顯著正相關。而Ferreira等[52]發現,在氮添加量高于 60 kg N hm-2a-1時會抑制植物細根的生物量和周轉速率,且抑制程度隨添加量上升而加重。Li等[53]則發現當氮添加量高于 100 kg N hm-2a-1時,細根生物量和周轉速率會受到顯著抑制。Sun等[54]發現,氮添加使 5 種不同森林類型的細根C/N和木質素/N都呈下降趨勢,且提高了細根周轉速率,但根系凋落物的分解速率均隨氮添加量的升高而顯著下降,分析根際胞外酶的活性后發現,氮添加促進了纖維素酶活性而抑制了木質素活性。關于土壤微生物如何響應氮添加來調節胞外酶分泌和活性,有研究就指出:氮限制條件下,氮添加會提高土壤養分有效性,導致更多的碳向微生物分配,促進微生物分泌胞外酶,從而提高微生物體外修飾作用,此舉有利于促進分解和降解外部有機物并轉化為SOC;而當氮添加過量時,土壤的酸化作用抑制會微生物分泌可降解復雜底物的胞外酶活性(如分解木質素的過氧化物酶、酚氧化酶等),從而降低微生物對根系凋落物的分解速率和釋放SOC[55]。Carrara等[56]發現在高氮添加量下植物會降低地下碳分配和土壤微生物活性,導致根系生物量下降了 24.7%、根際AM真菌生物量減少了 55.9%、胞外酶數量下降 18.7%,SOC轉化率大大降低。上述的相關研究表明,氮添加會降低植物細根的C/N和木質素/N,同時調節土壤微生物胞外酶的分泌及其活性來影響可分解性和分解速率來輸入SOC。
此外,根系分泌物和滲出物所形成的碳沉積也是SOC的重要來源,其中以根系分泌物為主。有研究表明,植物通過根系分泌物形式向根際土壤輸入的有機化合物約占光合產物的 5%—21%,分泌物的數量和成分受到植物種類、林齡以及根際土壤微環境等的影響[57]。根系分泌物的釋放主要受根際土壤養分有效性的調節,低養分有效性條件下會限制根系分泌物的分泌,而高養分有效性可促進根系分泌物的釋放,氮添加則傾向于增加根際土壤的養分有效性[58—60]。Qiao等[61]對亞高山針葉樹種進行了氮添加試驗,發現氮添加下植物根系分泌物顯著增多,以酚酸類化合物為主。龐麗等[62]研究了氮添加對馬尾松根系分泌物酸性磷酸酶和有機酸的影響,結果顯示,馬尾松不同家系根系分泌物對氮添加的響應均表現為促進作用。Ataka等[63]測定氮添加處理下寒溫帶森林根系分泌物的分泌速率,結果顯示氮添加顯著促進了根系分泌物的分泌速率和產量,使根際土壤中的DOC含量顯著提高。Iiu等[64]也發現,氮添加使溫帶森林植物白楊和白樺根系分泌物的分泌速率和根際土壤總有機碳(TOC)含量顯著提高。因此,氮添加傾向于促進根系分泌物和滲出物的釋放,同時調節基質的化學計量比及其可分解性,并最終被土壤微生物吸收轉化。Berhongaray等[65]的研究結論顯示,森林生態系統的SOC輸入主要來源于植被地下碳分配途徑的貢獻,地下碳分配(根系凋落物和根系分泌物、滲出物等)對SOC的轉化效率高達 76%;而地上凋落物對SOC轉化效率僅約 9%,絕大部分經由微生物礦化作用轉化為CO2釋放。這與Clemmensen等[66]、Cotrufo等[67]的研究結論一致。
由于氮添加使森林生態系統地上地下凋落物、根系分泌物和滲出物等有機質增多,土壤養分有效性提高,刺激了土壤微生物的體內修飾來促進外源有機質的分解,這提高了土壤的呼吸作用,使CO2排放通量升高。同時,受到外源有機質的刺激,土壤中原有的SOM會產生激發效應,加速SOC的礦化,該過程也會使土壤的CO2排放通量升高。因此,氮添加對凋落物分解腐化向土壤輸入SOC的實際影響還存在不確定性。但從全球尺度來看,絕大部分的氮添加研究,無論是基于野外的氮沉降觀測還是人為的施氮試驗,均提高了森林土壤的碳儲量,即促進森林生態系統的SOC輸入[68—70]。
2.2 氮添加對森林土壤有機碳庫及其組分的影響
森林土壤有機碳庫是由不同固存時間和分解階段的有機質所構成的混合體,包括植物源SOC和微生物源SOC,依據功能和性質的差異可分為活性有機碳、緩性(慢性)有機碳和惰性有機碳[71]。其中,活性有機碳易被吸收、分解和礦化,是植物營養、土壤微生物能量和CO2釋放的主要來源[72]。植物源碳(凋落物、根系凋落物、根系分泌物等)是森林土壤最主要的碳輸入,并在土壤微生物的介導下參與土壤碳循環過程。由于土壤中的碳、氮循環過程是緊密耦合的,因此氮添加所引起的土壤氮循環響應也會改變其中的許多碳循環過程,進而影響土壤有機碳庫及其組分的穩定性[73—74]。氮添加影響森林土壤有機碳庫組成和穩定性的途徑主要為以下 3 條:①影響植物碳同化和生理代謝過程,來改變凋落物、根系分泌物等植物源有機碳在數量及化學成分上的差異;②改變土壤養分有效性來影響土壤微生物的生命活動,進而調控SOM的降解、穩定和轉化,來改變不同SOC組分間的含量;③改變土壤的理化性質如pH、土壤團聚體結構和密度等,來破壞土壤環境對SOC的物理保護機制,進而影響土壤有機碳庫各組分的穩定性[75—76]。
作為陸地生態系統有機質的主要分解者,土壤微生物不僅控制了SOC的礦化過程,同時還是驅動各類有機碳組分降解轉化的“動力引擎”,主導著土壤的碳循環過程[77]。土壤微生物通過“體外修飾”及“體內周轉”途徑將植物源碳降解轉化,或以殘體的形式(腐殖質)儲存于土壤中;或被同化為微生物生物量和自身代謝產物,以微生物源碳形式儲存于土壤中。土壤微生物體外修飾即通過分泌胞外酶來分解或轉化大分子植物源碳底物,使其轉化為腐殖質;土壤微生物體內周轉即直接吸收小分子植物源碳底物后經同化作用合成為自身生物量。其中,經土壤微生物“體內周轉”途徑所同化的碳素,會隨著微生物的生命活動轉化為微生物生物量和自身代謝產物,并儲存在土壤中成為有機碳庫的一部分,這類有機碳組分被稱之為微生物源碳。關于以微生物“體內周轉”過程為驅動力的微生物源碳形成過程,Liang等[78]在2017年形象的提出了“土壤微生物碳泵(MCP)”的概念,在該研究中,Liang等人系統地梳理了土壤碳循環過程中土壤微生物調控SOC形成和穩定的相關途徑,并提出了“土壤MCP概念體系”,這是目前有關土壤微生物介導并參與SOC形成的最新理論[78],Liang等[79]還系統地闡述了土壤MCP概念體系的相關內涵和影響因素。
SOC的激發效應策略可以解釋土壤微生物在底物利用中“體內周轉”和“體外修飾”作用對氮添加的響應[80]。土壤微生物的群落組成主要受養分有效性驅動,依據養分吸收偏好的差異可將不同微生物分為r生活史策略(rate-strategistis)和K生活史策略(Kapazit?tsgrenze-strategistis)[81]。其中,細菌多為r策略型微生物,它們繁殖力強、生殖期短、迭代快,能快速分解吸收易降解的有機物質并大量投入到繁殖迭代中,而不是用于維持個體生長,主要利用外源底物[82];而真菌多為K策略型微生物,通常生長緩慢、發育周期和繁殖周期較長,所吸收的能量主要用于維持個體生存以提高存活率和競爭能力,傾向于分解吸收難降解的復雜有機物質,主要利用原有SOM來獲取能量[83]。在SOC礦化過程中,土壤中有機質的成分及結構差異會影響微生物的底物利用,進而影響土壤微生物的群落結構和功能,反之亦然[84]。在氮限制森林生態系統中,土壤養分有效性偏低,底物不足,此時K策略微生物的碳代謝過程占主導地位,可迅速分泌胞外酶來分解固存的SOM以獲取養分,該過程為SOC正激發效應,即微生物的氮挖掘機制,該過程會傾向于降低土壤有機碳庫固存并導致活性有機碳組分含量提高[85]。而隨著氮的添加,土壤養分有效性上升,底物逐漸增多,此時r策略微生物迅速生長并逐漸占據優勢,優先利用外源底物獲取能量,從而降低對SOM的分解,該過程為SOC負激發效應,即微生物底物偏好機制,該過程有利于SOM的穩定,并提高了MBC含量,有利于土壤有機碳庫固存[86—87]。因此在富氮森林生態系統中,不管氮添加與否,均傾向于r策略微生物主導SOC激發,促進土壤有機碳庫固存,并提高穩定性有機碳組分含量。
氮添加通過調控底物的化學組成差異來影響土壤微生物碳利用效率(CUE)的高低。CUE是指微生物通過“體內周轉”合成每單位生物量碳與所吸收的底物有機碳之比,該比值的熱力學限制為0.88[88]。即微生物在吸收底物轉化為自身生物量過程中,至少需要消耗12%的碳提供能量,但在實際中,由于環境條件的限制,維持土壤微生物生命活動所消耗的碳從未低于12%[89]。研究表明,底物的可分解性越高則CUE越高,也就是說,底物越容易被微生物吸收轉化的話,微生物耗能就越低,更能促進MBC組分積累而增加土壤有機碳庫[90]。根據推導,若凋落物和根系分泌物等底物所含的復雜化合物成分越低,則越有利于SOC轉化和固存。底物的可分解性取決于其養分有效性。例如,當底物的C:N:P化學計量比升高時,基質中的難分解化合物含量也會升高(即高分子量有機物質,如纖維素、半纖維和木質素等),土壤微生物在分解底物轉化為自身生物量過程中,會被迫分泌更多的胞外酶去捕獲短缺的營養元素來維持所合成的微生物生物量C∶N∶P化學計量比平衡,此過程會消耗更多的碳來提供能量[91—92]。由于氮添加提高了土壤養分有效性,促進森林植物生長和碳同化,提高了植物的凋落物產量和地下碳分配并降低了基質的C/N和木質素/N,從而使其分解、轉化過程中的CUE有效提高,因此相關的研究普遍得出氮添加促進了森林土壤有機碳庫固存和組分穩定性提高的結論[93—94]。
根據Meta分析,在全球尺度上,氮添加(沉降)使森林生態系統碳匯每年增加了 276—448Tg,其中約 40% 為森林土壤有機碳庫固存[95]。例如,以氮限制森林生態系統為例,Vandeneden等[96]在為期 9 年的研究中發現,氮添加使溫帶森林土壤中的纖維素酶和多酚氧化酶活性顯著下降,抑制了SOM的分解速率,引起土壤有機碳庫增加的同時也提高了難分解有機碳的含量。當土壤形成富氮環境時亦是如此,Hyvonen等[68]、Du等[97]的研究均表明,隨著氮添加量增加會導致土壤中有機碳礦化速率降低而促進SOC積累,并促進DOC組分、LOC組分和ROC組分轉化為穩定的有機碳組分固定。Liu等[98]發現,長期氮添加會降低森林土壤中微生物對惰性有機碳的分解速率,進而使森林土壤有機碳庫中惰性有機碳比例不斷上升,有利于土壤碳庫的長期固存。郭潔云等[99]采用整合分析法,對 172 篇已發表的森林土壤氮添加論文進行了綜合分析,結果表明氮添加顯著增加了森林凋落物產量,顯著降低了凋落物的C/N;絕大多數的研究都顯示氮添加提高了土壤有機碳庫固存并改變了各組分含量,使有機質層土壤中活性有機碳的DOC和MBC組分含量提高,而礦質層土壤中活性有機碳的DOC和MBC組分含量下降,惰性有機碳含量提高。
除了上述土壤微生物介導所引起的土壤有機碳庫及其組分含量對氮添加的響應變化過程,土壤的物理保護機制也對氮添加下的土壤有機碳庫固存和穩定過程起到了一定作用[100—101]。土壤微生物源源不斷的迭代過程會導致微生物死亡殘體和部分代謝產物持續增加,這些微生物殘留物類有機質隨著時間的推移趨向于在土壤中積累并穩定固存,該過程稱之為“續埋效應”[78]。研究認為,在“續埋效應”過程中,土壤團聚體可吸收SOC形成團聚體結合態有機碳,使其避免被分解,從而促進有機碳庫穩定[102—103];此外,土壤的礦物質也可以吸附SOC或直接與其結合形成礦物結合態有機碳[104],該現象被Xiao等[105]稱為礦物碳泵效應。目前認為,氮添加會有利于土壤團聚體形成并吸收SOC[106]。此外,相關研究表明,氮添加會改變SOC的垂直分布格局,使有機質層SOC向礦質層轉移,并在礦物質的物理保護作用下成為礦物結合態有機碳組分(惰性SOC),增加有機碳庫的穩定性[107—109]。
如表1列舉的研究結果所示,受氮添加量、施氮方式、施氮持續時間以及氣候、土壤基質和森林植被類型等因素的共同影響,森林土壤有機碳庫中不同組分對氮添加的響應并非呈現線性關系。盡管未能揭示不同森林生態系統土壤有機碳庫對氮添加的具體響應規律,但梳理氮添加對不同森林生態系統SOC輸入、轉化和分解過程的相關研究結論,有助于預測未來氮沉降(施氮)對不同森林生態系統碳匯功能的影響,以便及時調整施肥策略以提高森林碳匯能力緩解全球溫室效應。
3 氮添加對森林土壤CO2排放的影響及其作用機制
3.1 森林土壤排放CO2的途徑及其對氮添加的響應
釋放CO2是土壤碳循環中的碳輸出過程,由土壤呼吸作用和無機碳解吸附、溶解過程產生,其中主要由土壤呼吸作用所釋放。土壤呼吸作用包括植物根系自養呼吸、土壤微生物和動物的異氧呼吸以及含碳化合物的化學氧化過程[119]。研究表明,森林土壤釋放CO2的途徑主要來源于植物根系自養呼吸和土壤微生物異氧呼吸,其中又以土壤微生物的異氧呼吸為主,占森林土壤CO2總排放量的 60% 以上[120—121]。土壤微生物通過呼吸作用氧化有機物質來獲取能量并釋放CO2,用以維持其生命活動和生態功能,即SOC礦化,該過程的速率和程度會直接影響CO2排放,而通過SOC礦化途徑所釋放的CO2可占到森林土壤CO2總排放量的 70% 以上[122—123]。因此,微生物呼吸作用的大小可表征土壤的CO2排放強度。
在氮添加條件下,土壤有機碳庫與SOC礦化、土壤微生物之間會發生復雜的交互聯系。如圖2 所示,氮添加會提高土壤的養分有效性,促進植物根系養分吸收,提高了植物的碳同化能力,繼而增加了植物源碳輸入,在土壤微生物介導下,植物源碳輸入會產生激發效應引起SOC礦化,而土壤微生物的活性和生物量又受氮素有效性的調控。同時,氮添加會促進植物根系生命活動,由此提高根系自養呼吸釋放CO2;氮添加引起的土壤酸化也會誘導土壤無機碳溶解釋放CO2[97]。因此,目前研究結論可以明確的是:①氮添加提高了植物的根系呼吸作用,因此促進了根系途徑釋放CO2[124];②氮添加降低了土壤pH,促進了土壤無機碳的溶解并釋放CO2[125]。除此以外,由于土壤有機碳的激發效應存在正激發和負激發,正激發會促進SOC積累而減少CO2排放,負激發會促進SOC分解而增加了CO2排放,而激發效應的方向受到外源有機質底物數量和化學組成、土壤微生物群落組成和功能、土壤環境條件等眾多因素的影響,導致激發效應途徑的CO2排放與否難以確定,因此難以準確判斷不同森林土壤CO2排放通量對氮添加的響應規律。總的來說,不同森林土壤CO2排放對氮添加的響應分為促進、抑制和無顯著影響[126—128]。
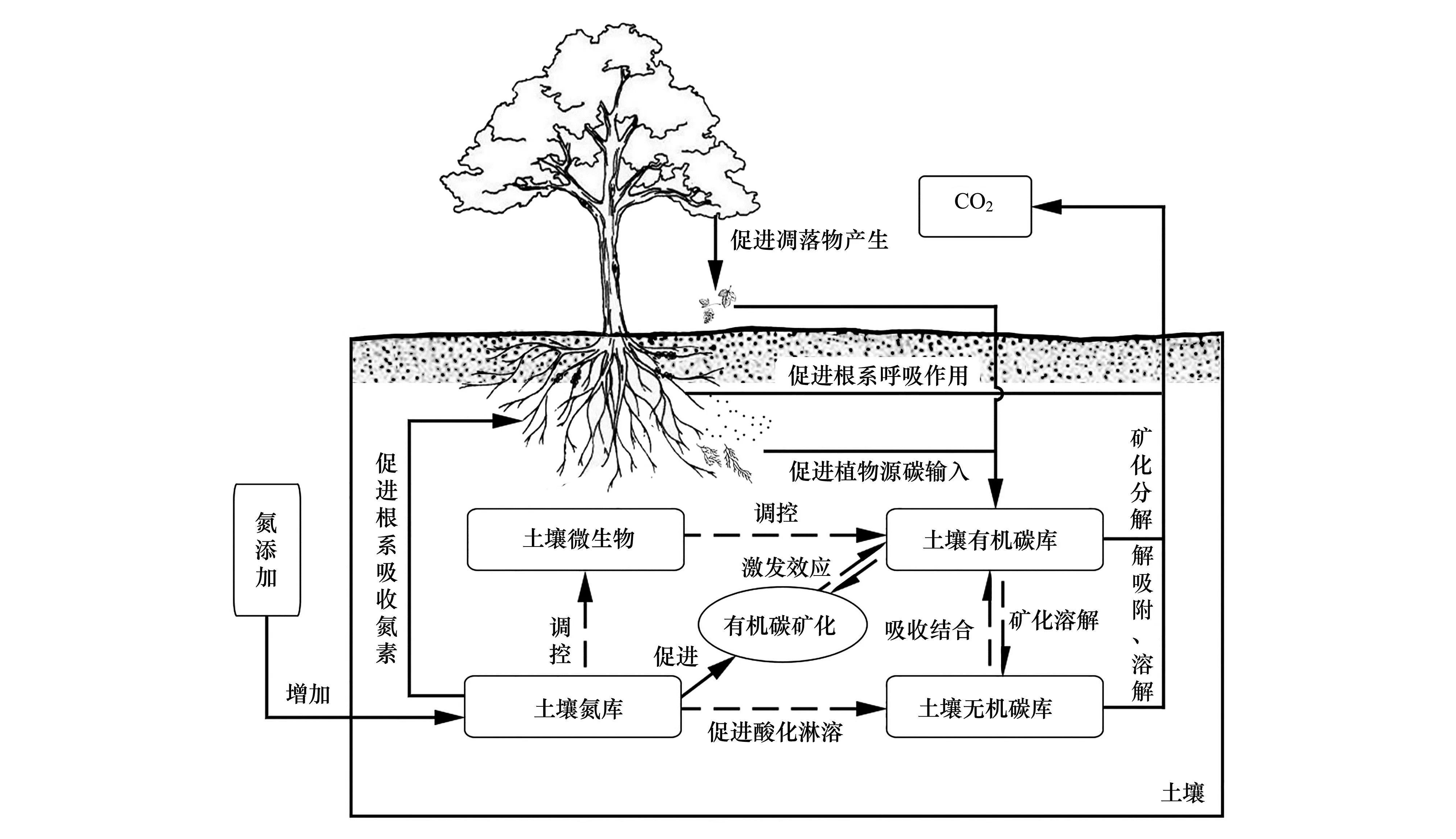
圖2 氮添加對森林土壤CO2排放的影響機制Fig.2 Mechanism of the effect of nitrogen addition on CO2 emissions from forest soil
3.2 氮添加對森林土壤CO2排放的促進作用
在氮限制森林中,氮添加提高了土壤養分有效性,傾向于促進土壤微生物活性和植物生長,由此提高森林土壤的呼吸作用而促進CO2排放。促進機制包括:①氮添加促進植物生長發育使根系生物量增多,提高了根系自養呼吸速率;②氮添加促進了森林植物的光合碳同化,其凋落物回歸和地下碳分配為土壤提供了更多底物,促進了SOC礦化的正激發效應;③氮添加為土壤微生物提供了氮源而促進其生長繁殖,提高了異氧呼吸速率,同時土壤微生物活性的提高進一步促進了SOC礦化的正激發效應。
因此,在氮限制的森林生態系統中,氮添加往往可促進森林土壤釋放CO2。王澤西等[129]研究發現,氮添加顯著促進了氮限制高山森林土壤呼吸速率,使森林土壤呼吸通量提高了 11%—15%,CO2排放通量隨施氮量增加先升高后降低,在中氮添加水平(50 kg N hm-2a-1)達到最高。Str?mgren等[130]在溫帶氮限制云杉林中連續施氮 22 年,發現森林土壤有機質層和礦質層SOC礦化速率分別提高了 118%和 19%,土壤CO2排放通量提高了 31%。Sun等[131]發現氮添加不僅顯著促進了青藏高原地區高山針葉林土壤活性有機碳的礦化,還促進了惰性有機碳的分解,使土壤CO2排放通量明顯提高。一些研究發現,氮添加量和氮素形態會影響其促進森林土壤CO2排放的效果。Sun等[131]發現,在養分貧瘠地區,森林土壤的CO2排放量隨氮添加量升高而顯著增多。李睿達等[132]在熱帶桉樹人工林施肥研究中發現氮添加顯著促進了土壤CO2排放,且隨著施氮量的增加而顯著增多。Geng等[133]在溫帶養分貧瘠的針闊混交林中開展氮添加試驗,發現當添加量低于 100 kg N hm-2a-1時土壤CO2排放通量顯著提高,而當氮添加量超過100 kg N hm-2a-1后,會顯著抑制CO2排放。Li等[134]對青藏高原針闊混交林開展的梯度氮添加結論也與該研究一致。
3.3 氮添加對森林土壤CO2排放的抑制作用
持續的氮添加會造成土壤酸化,而土壤pH過低被認為是毒害效應,會使土壤微生物活性降低,呼吸作用受限,繼而抑制CO2的排放量[135]。由此可得,氮添加抑制森林土壤CO2排放的機制可能是:①氮添加降低了土壤的C/N和pH,造成土壤微生物活性和生物量下降,因此降低了微生物的異氧呼吸;②氮添加使根際土壤養分失衡,使植物根系生物量和細根周轉速率受到影響,從而降低了根系的自養呼吸;③氮添加造成的氮富集效應抑制了土壤微生物和胞外酶活性,降低微生物群落的碳源獲取能力,提高了CUE,降低了SOC礦化的正激發效應,從而減少了CO2釋放[136]。
大量研究表明,在非氮限制的森林生態系統中,氮添加通常會抑制土壤微生物的活性而降低SOC的礦化速率,從而抑制了森林土壤的CO2排放[95]。無論是室內培養試驗還是野外試驗,若土壤屬于富氮狀態,氮添加均有可能會抑制土壤CO2排放[137]。Wang等[138]采集了富氮的亞熱帶針葉林的土壤進行室內氮添加培養試驗,發現氮添加使土壤呼吸速率下降了 6.6%,SOC礦化速率明顯抑制。Lu等[139]在熱帶富氮森林中開展野外氮添加研究,發現不同梯度的氮添加均顯著降低了SOC礦化速率,CO2排放通量與對照相比顯著減少。Zhang等[140]在亞熱帶富氮森林中開展的梯度氮添加試驗也與Lu等人的研究結論一致。Micks等[141]則發現,在溫帶富氮森林中,氮添加使土壤呼吸速率下降,同時CO2排放通量受到抑制。
氮添加量和施氮時間是影響森林土壤CO2排放通量的關鍵因素[142]。短期的低氮添加量通常促進土壤CO2排放或無影響、而長期的低氮添加量往往可以促進土壤CO2排放;短期的高氮添加量通常抑制土壤CO2排放或無影響、而長期的高氮添加量往往可以抑制土壤CO2排放[143—144]。Forsmark等[145]在氮貧瘠的溫帶針葉中開展了為期 12 年的梯度氮添加試驗,發現土壤呼吸作用對低氮添加量的響應并不敏感,土壤CO2排放通量與對照相比無顯著差異,但高氮添加量下微生物呼吸作用受到抑制,土壤CO2排放通量顯著降低。張雪等[146]通過盆栽試驗,證實短期的低氮添加對馬尾松盆栽土壤CO2的釋放量有促進作用,CO2釋放速率在氮添加后的短期內顯著提高,但在施氮后期迅速下降。而Chen等[147]則發現,高氮添加(100 kg N hm-2a-1)初期對土壤呼吸速率并無顯著影響,但隨著時間的推移,土壤呼吸速率顯著降低,CO2排放通量明顯下降。
3.4 氮添加對森林土壤CO2排放量無顯著影響
由于森林土壤所釋放的CO2主要來源于土壤的呼吸作用,若氮添加并未對森林土壤的呼吸作用產生顯著影響,那么森林土壤CO2排放量對氮添加的響應特征可能也表現為無顯著影響。關于氮添加對森林土壤CO2排放并未產生顯著影響的原因,可能受氮添加量不足和施氮時間過短的影響。有研究表明,短期的氮添加可能不會改變森林土壤的CO2排放通量。Sun等[148]以亞熱帶濕地松林土壤開展為期 1 年的氮添加試驗,發現該森林土壤的CO2排放通量并未受到影響。Maestrini等[149]在溫帶闊葉林中以 60 kg N hm-2a-1的氮添加量每月等量添加 1 年后,發現土壤CO2排放通量與對照相比無顯著差異。
此外,在土壤理化性質和微生物的共同作用下,森林土壤的CO2排放量可能也不會產生顯著波動。Allison等[150]在寒帶針葉林中開展氮添加試驗,發現氮添加降低了土壤微生物生物量的C/N,但并未改變微生物的生物量,對土壤呼吸速率及CO2排放通量無影響。Fernández等[151]在地中海地區的針闊混交林中進行氮添加試驗,發現氮添加主要與土壤溫度協同影響森林土壤的呼吸作用,但與不施肥的森林土壤相比,總CO2排放量并無顯著差異。Mori等[152]發現,在馬來西亞的原始熱帶雨林地區,由于受磷限制影響,氮添加未顯著改變土壤自養呼吸和異氧呼吸速率,從而并未對CO2排放通量造成顯著影響。Koehler等[153]在熱帶森林中進行的氮添加試驗(125 kg N hm-2a-1)同樣也顯示對森林土壤CO2排放通量無顯著影響。在富磷的非熱帶地區,也得出了類似Mori等和Koehler等的研究結論,Gao等[154]研究了氮添加對富磷環境的亞熱帶混交林土壤溫室氣體排放通量的影響,發現氮添加對該森林土壤的CO2排放通量無顯著影響。
4 結論與展望
總體而言,氮添加通過提高森林土壤的養分有效性而提高了森林的凈初級生產力,促進了凋落物和根際沉積碳(細根周轉產物、根系分泌物及滲出物等)產量,同時降低了上述基質中的大分子化合物含量,使其可分解性提高;盡管不同森林生態系統的土壤微生物群落特征及生態功能對氮添加的響應規律存在錯綜復雜的耦合和交互影響,但均傾向于促進植物源SOC的積累并趨向于穩定,有效提高了土壤有機碳庫固存和穩定性。氮添加對森林土壤的CO2釋放規律并不呈現簡單的線性關系,由于受到底物數量和質量、微生物群落組成和活性、土壤理化性質、氣候、植被類型等眾多因素的共同影響,不同森林生態系統的CO2排放量可能表現為促進、抑制或無影響。
森林土壤有機碳庫在全球碳循環過程中扮演著重要的角色,對維系生態系統碳循環穩定和緩解溫室效應而言舉足輕重。近年來,林業化肥濫用現象日漸嚴重,加劇了溫室效應,在全球氣候變暖的背景下,氮素輸入對森林土壤碳循環過程的影響越來越受到關注。由于受到植被類型、土壤理化性質、微生物種類和氣候等因素的多重影響,氮添加對森林土壤有機碳庫固存機制及CO2排放的研究尚不全面,人們對氮添加條件下森林植被的地下碳分配格局規律也缺乏系統認識,此外森林土壤有機碳庫中活性SOC向惰性SOC轉化的穩定機制及其與土壤微生物的耦聯關系并不清晰。因此,在今后的研究中可加強以下方面的突破:
(1)加強長期氮添加對森林土壤有機碳庫固存及穩定性影響的研究。研究發現,土壤有機碳庫及其組分通常對短期的氮添加無響應,而對長期的氮添加有顯著響應。建立長期的氮添加對森林土壤有機碳庫影響的研究可更好地掌握土壤有機碳庫的穩定固存機制,以便有效預測氮沉降對森林土壤有機碳庫的長期動態影響,這有利于尋找提高森林土壤有機碳庫固存能力和降低溫室氣體的森林碳匯途徑。
(2)加強全球氣候變暖背景下土壤微生物碳代謝過程對氮添加(氮沉降)的響應研究。氮添加主要引起土壤氮素有效性增加,這一變化會直接引起土壤微生物群落響應,進而影響CUE、改變土壤呼吸作用強度,而氣候變暖也會影響微生物的活性和生態功能,這進一步造成了森林生態系統CO2排放規律的不確定性。加強上述研究有望緩解目前的溫室效應困局。
(3)進一步探索氮沉降背景下森林植被差異對SOC分解、轉化和固定過程的影響機制。明確不同森林生態系統植物多樣性對土壤微生物種群結構和生態功能的影響,如優勢植物是否參與(或主導)了構建土壤微生物群落生態功能,繼而影響森林土壤的碳循環過程。
(4)借助微生物組學技術,從土壤—植物—微生物互作角度全面研究氮添加下土壤微生物群落及功能多樣性改變對森林土壤有機碳輸入及輸出的影響。借助微生物組學技術可以更全面、系統地解析氮添加對森林土壤微生物群落和功能多樣性的影響,有助于進一步闡明土壤微生物對SOC輸入和輸出的影響。
(5)加強氮添加對森林根際碳沉積途徑輸入SOC的相關機制研究。目前的研究主要集中在氮添加影響下凋落物回歸后的土壤碳循環過程響應及相關介導機制,但森林土壤碳庫的碳輸入過程還包括植物地下碳分配調節下的根際沉積碳途徑,而該過程對氮添加的響應十分復雜,關于氮添加下的植物—菌根互作機制、根際分泌物和滲出物響應規律、根際沉積物化學計量學特征等規律尚無明確的科學結論。
(6)結合其他關鍵影響因子研究氮添加對森林土壤碳循環過程的耦合效應。森林土壤的碳循環是一個復雜而綜合的過程,受到眾多因素的共同影響。因此有必要結合其他關鍵因子如氣候變暖、CO2濃度升高、降雨等來研究氮添加對森林土壤碳循環的耦合作用,此舉有助于分析在全球氣候變化下氮添加對森林土壤有機碳庫固存的影響。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
