白術多糖通過花生四烯酸代謝通路緩解環磷酰胺誘導的小鼠腎損傷
2024-05-12 09:10:08李冰心龔淑影許丹寧李婉雁付新亮江丹莉洪龍勝余梓榆白帥飛田允波
食品工業科技 2024年10期
李冰心,龔淑影,許丹寧,李婉雁,曹 楠,付新亮,江丹莉,洪龍勝,余梓榆,白帥飛,田允波
(仲愷農業工程學院動物科技學院,廣東廣州 510225)
白術多糖(Polysaccharide ofAtractylodes macrocephalaKoidz,PAMK)是中藥白術最主要的有效生物活性成分之一。研究表明,白術多糖具有抗炎、抗氧化、降血糖與免疫調節等多種生物活性[1-3]。在免疫方面,PAMK 通過改善肝脂代謝和增加抗氧化酶活性,有效地保護肝臟免受高能量低蛋白飲食引起的損傷,進一步減輕肝臟負擔,從而顯著提升免疫系統的功能[4],并通過介導信號通路減輕肝臟炎癥損傷和氧化應激[5]。此外,PAMK 還可以通過提高白細胞數量、修復骨髓、脾臟及胸腺的組織結構、提高白細胞功能等途徑減輕環磷酰胺對小鼠白細胞的抑制和破壞作用,并對正常小鼠的淋巴細胞功能具有明顯的提升作用[6]。
腎臟是哺乳動物重要的排泄器官,在維持體內平衡中起著至關重要的作用。環磷酰胺(Cyclophosphamide,CTX)是一種常用的抗癌藥物,但它也會產生許多副作用,包括尿毒性或腎毒性[7]。CTX 的代謝轉化導致兩種代謝物的形成:磷酰胺芥末和丙烯醛,磷酰胺芥被認為具有抗腫瘤活性,而丙烯醛作為一種生物半衰期短的高活性代謝物,可能是CTX 誘導腎損傷的原因[8-9]。丙烯醛可以產生高活性氧(ROS),干擾組織抗氧化防御系統,對哺乳動物細胞具有致突變性[10-12],為了避免這些氧化毒性副作用,當前常用抗氧化劑對丙烯醛進行解毒,比如芍藥苷已被用于緩解CTX 引起的腎損害[13]。Gunes 等[14]發現香芹酚能夠作為一種抗氧化劑,保護生物分子,如膜脂,免受自由基誘導的損傷,并且能夠減少CTX 注射后大鼠的氧化應激的反應。然而,PAMK對CTX 誘導的小鼠腎損傷的影響尚未見報道。
本研究擬通過注射CTX 建立小鼠腎損傷模型,以探究PAMK 對CTX 誘導的小鼠腎損傷的影響,并通過轉錄組測序掌握PAMK 緩解CTX 誘導的小鼠腎損傷的作用機制,為后續研究PAMK 緩解CTX誘導的機體損傷提供參考。
1 材料與方法
1.1 材料與儀器
PAMK 純度95%,中國西安楊凌慈源生物技術公司;注射用環磷酰胺國藥準字H14023686 山西普德藥業股份有限公司;SPF 級C57BL/6 雄性小鼠 42~43 日齡,100 只,購自廣東省醫學實驗動物中心;超氧化物歧化酶(SOD)、谷胱甘肽過氧化物酶(GSH-Px)、總抗氧化能力(T-AOC)、丙二醛(MDA)、過氧化氫酶(CAT)試劑盒 南京建成生物工程研究所;預混型定量用反轉錄試劑盒 日本TAKARA 公司;Trizol 試劑 美國Thermo Fisher 公司;SYBR Green Master Mix PCR 中國康潤生物技術公司。
Epoch 超微量微孔板分光光度計 美國伯騰儀器有限公司;7500 REAL TIME PCR SYSTEM 實時熒光定量PCR 儀、VERITI 梯度PCR 儀 美國ABI 公司;Micro 21R 高速冷凍離心機 美國Thermo Fisher 公司;Bioanalyzer 2100 2100 生物分析系統 美國Agilent 公司;NovaSeq 6000 基因DNA 測序儀 美國Illumina 公司。
1.2 實驗方法
1.2.1 試驗設計與樣品采集 100 只42~43 日齡雄性C57BL/6 小鼠被隨機分為四組,分別為對照組(Control)、環磷酰胺組(CTX)、白術多糖組(PAMK)、白術多糖+環磷酰胺組(PAMK+CTX),25 只/組,每組5 個重復,每個重復5 只。實驗期間,PAMK 組和PAMK+CTX 組灌胃200 mg/kg PAMK 1 次/d;Control 組和CTX 組給予等量的生理鹽水。實驗第25~27 d,CTX 組和PAMK+CTX 組腹腔注射100 mg/kg CTX 1 次/d;Control 組和PAMK 組注射等量生理鹽水。實驗第35 d,每組隨機選取5 只小鼠采集腎臟組織,取部分腎臟組織立即凍入液氮,-80 ℃保存備用;剩余組織樣品于10%中性緩沖福爾馬林中固定保存。本試驗所有小鼠均接受人道處理,試驗經仲愷農業工程學院實驗動物倫理委員會批準,批準協議號為:2023030501。
1.2.2 腎臟組織學及超微形態學觀察 腎臟組織在10%中性緩沖福爾馬林溶液中固定48 h,包埋石蠟,切成5~6 μm 厚的切片。常規組織學檢查,石蠟切片蘇木精-伊紅染色法(HE 染色),HE 染色切片在光學顯微鏡下觀察并用Case Viewer 軟件進行圖像采集。
1.2.3 腎臟抗氧化指標檢測 取0.2 g 腎臟組織加入1.8 mL 磷酸鹽緩沖液(PBS),經組織破碎儀破碎制成10%組織勻漿,3000 r/min 離心10 min,取上清待測(n=5)。按照試劑盒的說明,檢測組織中超氧化物歧化酶(SOD)、谷胱甘肽過氧化物酶(GSH-Px)、總抗氧化能力(T-AOC)、丙二醛(MDA)、過氧化氫酶(CAT)含量。
1.2.4 RNA 提取及文庫構建 用Trizol 試劑提取腎臟組織總RNA,利用NanoDrop ND-1000 分析總RNA 質 量和 純度(濃度>50 ng/μL、RIN 值>7.0、OD260/280>1.8),并通過Bioanalyzer 2100 儀器檢測RNA 完整性。每個樣本分別取5 μg RNA 用于轉錄組文庫構建,運用Epicenter Ribo-Zero Gold 試劑盒去除樣本中的核糖體RNA(rRNA),剩余RNA 純化后進行反轉錄以構建cDNA 文庫。通過聯川生物技術有限公司的Illumina Novaseq? 6000 平臺對構建的小鼠腎組織cDNA 文庫進行雙端測序,測序模式為PE150。測序完成后,使用Cutadapt 軟件刪除包含連接物污染、低質量堿基和未定義堿基讀數的原始數據(raw reads)。利用FastQC 軟件(http://www.bioinformatics babraham.ac.uk/projects/fastqc/)獲得高質量的clean reads,并且將該數據與小鼠(Mus musculus)基因組序列進行序列比對。利用Tophat2檢測測序數據的飽和度,同時對比對上的reads 進行統計分析,以評估clean reads 在內含子(intron)、外顯子(exon)和基因間隔區(intergenic)的分布情況。
1.2.5 差異表達基因GO 功能及KEGG 通路富集分析 采用DESeq2 軟件對兩組間基因差異表達進行分析,以P<0.05 且|log2(Fold change)|≥1 為閾值,篩選差異表達基因。利用Goseq R 軟件包和Kobas 在線工具(http://kobas.cbi.pku.edu.cn/genelist/)分別對差異表達基因進行GO 功能和KEGG 功能富集通路分析。GO 功能富集包括生物過程、細胞成分和分子功能。以P<0.05 作為篩選顯著富集的GO 條目或KEGG 信號通路的標準。利用在線網站Venny 2.1.0(https://bioinfogp.cnb.csic.es/tools/venny/index.html)對兩組KEGG 富集的通路進行交集分析。
1.2.6 轉錄組測序基因實時熒光定量PCR 檢測 根據RNA-Seq 結果挑選花生四烯酸通路相關基因CYB2B9、CYP2C65、NF-κB、PTGS1、ALOX5、ALOX12、BCL6、MFSD2A進行實時熒光定量PCR。使用Trizol 試劑,嚴格按照說明書提取相同批次的RNA,按照TaKaRa 逆轉錄試劑盒說明書將RNA 逆轉錄為cDNA。根據GenBank 中各基因序列,利用Oligo 7 軟件設計引物,引物信息見表1。引物均由上海生工生物工程有限公司合成。在實時定量PCR 采用SYBR Green 嵌合熒光法,反應體系包括SYBR Green Master Mix 10 μL,上游引物0.5 μL,下游引物0.5 μL,cDNA 1 μL,RNase free ddH2O 8 μL。反應程序為:95 ℃預變性2 min;95 °C 變性15 s,60 ℃退火30 s,72 ℃延伸30 s,40 個循環。采用2-ΔΔCt法計算相對表達量。

表1 引物信息Table 1 Information of the primers
1.3 數據處理
利用GraphPad Prism 8.0 軟件進行統計、作圖。根據one-way ANOVA 比較平均值,并通過Tukey 檢驗進行組間多重比較,數值以“平均值±標準誤”表示。P<0.05 表示差異顯著。
2 結果與分析
2.1 PAMK 對CTX 誘導的小鼠腎臟組織結構的影響
各組小鼠的腎組織切片如圖1 所示,Control 組和PAMK 組小鼠腎臟結構更為完整,腎小管包括近曲小管及遠曲小管各部位上皮細胞無變性,排列有序,腎小囊明顯,腎小體、腎小球等結構均正常,管腔形態大小正常,未見異狀。CTX 組出現了明顯的病理現象,如腎小球萎縮、腎小管上皮細胞脫落、周圍有少許出血點和炎性細胞浸潤;與CTX 組相比,PAMK+CTX 組腎組織病變明顯好轉。
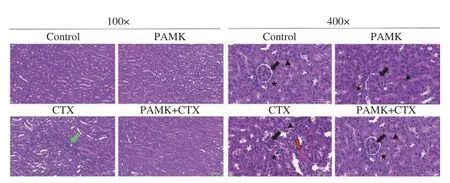
圖1 小鼠腎臟組織病理學變化(H&E 染色,100×,400×)Fig.1 Histopathological changes of mouse kidney (H&E staining,100×,400×)
2.2 PAMK 對CTX 誘導的小鼠腎臟抗氧化指標的影響
腎臟組織中MDA 含量、GSH-Px、SOD、CAT活性和總抗氧化能力檢測結果如圖2 所示,與Control 組相比,注射CTX 后的小鼠腎組織的SOD、GSH-Px 活性顯著降低(P<0.05);MDA 含量顯著升高(P<0.05);相較于CTX 組,CTX+PAMK 組小鼠腎組織的GSH-Px、SOD、CAT 活性和總抗氧化能力顯著升高(P<0.05);MDA 含量顯著下降(P<0.05)。與Control 組相比,PAMK 組小鼠腎組織中的總抗氧化能力顯著升高(P<0.05),GSH-Px、SOD、CAT 活性無顯著差異。
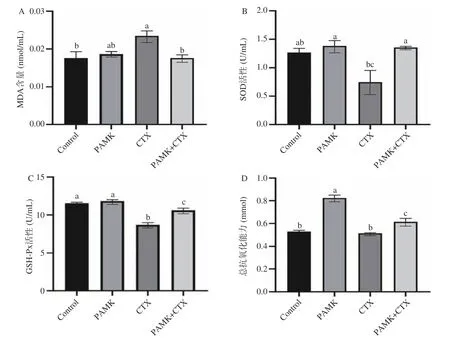
圖2 PAMK 對小鼠機體中氧化應激指標的影響Fig.2 Effects of PAMK on oxidative stress indexes in mice
2.3 轉錄組測序數據分析
本研究構建的9 個cDNA 文庫中獲得了37125316~43483558 條清潔讀數(clean reads),有效比對率在94.36%以上,所有樣本的Q20%值均>99.90%,Q30%值均>97.81%,GC 含量(RNA 序列中,含有鳥嘌呤(Guanine,簡稱G)和胞嘧啶(Cytosine,簡稱C)這兩種堿基的比例在47.50%~50.00%,說明測序質量的錯誤率低,數據可靠性高,可用于后續分析。通過對各樣本的mRNA 區域分布進行統計分析,結果顯示,發現各樣本中外顯子(exon)、內含子(intron)和基因間隔區域(intergenic)所占比例相似,其中由外顯子轉錄而來的mRNAs 片段比例最高,占95%以上(圖3)。
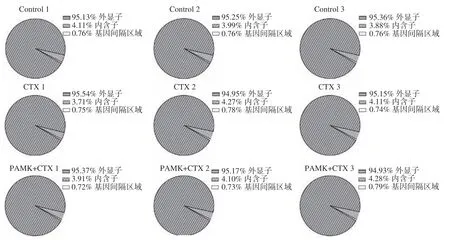
圖3 小鼠腎臟組織中mRNAs 分布統計Fig.3 Distribution statistics of mRNAs in bursae of Fabricius of mouse
2.4 差異表達基因分析
通過DESeq2 軟件對轉錄組測序數據進行分析,結果發現,Control 組vs CTX 組共獲得493 個差異表達基因,其中236 個基因上調,257 個基因下調(圖4A~圖4B);CTX 組vs PAMK+CTX 組共獲得333個差異表達基因,其中184 個基因上調,149 個基因下調(圖4C~圖4D)。
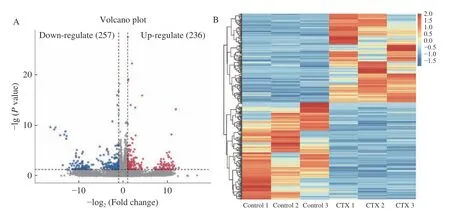
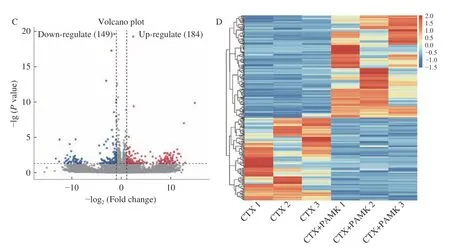
圖4 差異表達基因火山圖和熱圖Fig.4 Volcano map and heatmap of differentially expressed genes
2.5 差異表達基因GO 功能與KEGG 通路富集分析
GO 功能富集分析發現,差異表達基因多富集在與炎癥和免疫反應相關的GO 條目,其中Control組vs CTX 組主要富集在環氧合酶P450 途徑、花生四烯酸環氧合酶活性、轉錄調控區DNA 結合的負調控等途徑(圖5A);CTX 組vs PAMK+CTX 組主要富集在花生四烯酸環氧合酶活性、細胞色素P450 途徑、絲氨酸型肽酶活性等途徑(圖5C)。KEGG 通路富集分析結果顯示,Control 組vs CTX 組差異表達基因主要富集在花生四烯酸代謝(T 細胞受體、NFκB 等信號通路(圖5B);CTX 組vs PAMK+CTX 組主要富集在花生四烯酸代謝、亞油酸代謝、TRP 通道的炎癥介質調節等信號通路(圖5D)。表2 和圖6 結果顯示,Control 組vs CTX 組、CTX vs PAMK+CTX組的KEGG 富集通路中有交集的通路有7 個。其中花生四烯酸代謝通路在Control 組vs CTX 組和CTX 組 vs PAMK+CTX 組中的富集最顯著。
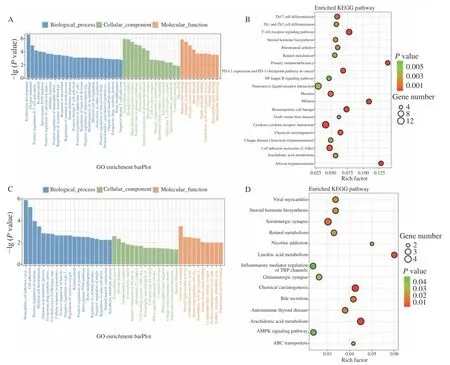
圖5 差異表達基因GO 功能(A、C)和KEGG 通路(B、D)富集分析Fig.5 GO function (A,B) and KEGG pathway (B,D) enrichment analysis of differentially expressed genes
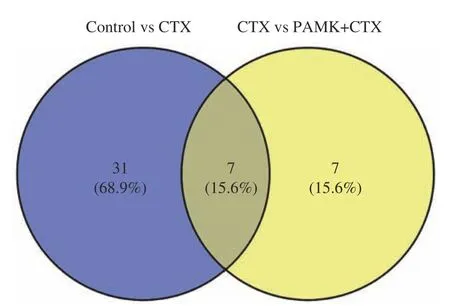
圖6 Control vs CTX 和CTX vs PAMK+CTX KEGG 富集通路交集維恩圖Fig.6 Gene intersection Venn diagram of Control vs CTX and CTX vs PAMK+CTX KEGG enrichment pathway

表2 Control vs CTX 和CTX vs PAMK+CTX KEGG交集通路Table 2 Intersection pathways of Control vs CTX,CTX vs PAMK+CTX KEGG
2.6 實時熒光定量PCR 檢測
用實時熒光定量檢測Control 組vs CTX 組和CTX 組vs PAMK+CTX 組關鍵基因CYP2C65、CYP2B9、PTGS1、ALOX5、ALOX12、NF-κB和抗氧化基因BCL6、MFSD2A。由圖7 可知,與Control組相比,CTX 組Cyp2b9、PTGS1、NF-κB和Mfsd2amRNA 表達量顯著升高(P<0.05),BCL6mRNA 表達量顯著降低(P<0.05),Cyp2c65、ALOX5和ALOX12mRNA 表達量無顯著差異(P>0.05);與CTX 組相比,PAMK+CTX 組Cyp2c65、BCL6mRNA 表達量顯著升高(P<0.05),Cyp2b9、PTGS1和Mfsd2amRNA表達量顯著降低(P<0.05);ALOX5、ALOX12和NFκBmRNA 表達量無明顯差異。
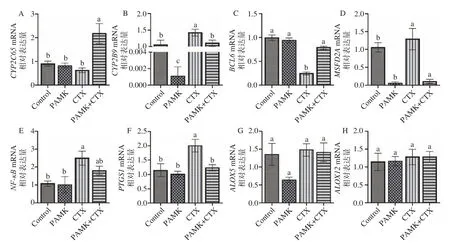
圖7 PAMK 對CTX 處理下小鼠腎臟組織相關基因的mRNA 相對表達量的影響Fig.7 Effect of PAMK on the relative expression of mRNA for genes in mouse kidney tissue treated with CTX
3 討論與結論
腎臟能清除有毒物質,如果腎臟功能不正常,會影響機體有害物質的排泄,使毒素的積累不能立即清除,從而可能導致各種疾病。目前腎臟疾病的防治已成為眾多學者關注的焦點[15]。通過觀察小鼠腎組織病理切片,HE 染色結果顯示CTX 組腎小球萎縮,腎小管腔變形,周圍有少許出血點并有炎性細胞浸潤,導致腎組織出現明顯的病理損傷。與CTX 組相比,經白術多糖治療后,腎臟損傷程度明顯改善,主要表現為腎小管結構清晰,周圍炎性細胞明顯減少,表明PAMK 治療逆轉了CTX 致腎臟組織炎癥損傷。
本試驗發現CTX 處理后,MDA 含量增加、SOD、GSH-Px、CAT 活性降低。經過PAMK 治療后,MDA 的積累減少,小鼠機體內SOD、GSH-Px、CAT 活性和總抗氧化能力顯著提高。CTX 的使用與機體發生氧化應激密切相關,有研究表明CTX 會引起動物體內MDA 升高,抑制GSH-Px、CAT 和SOD 的活性,削弱機體的抗氧化能力,破壞氧化系統和抗氧化系統的平衡,導致機體發生氧化應激現象[16]。氧化應激反應是一種高響應性的防御機制,如果長期處于氧化應激狀態,可能會導致多種疾病的發生[17]。SAKR 等[18]發現了注射環磷酰胺可引起大鼠腎血管的擴張和充血,腎小管上皮細胞的空泡化以及腎小球的萎縮,并推測引起這些病理變化的原因可能是環磷酰胺誘導的氧化應激造成的。說明了CTX 處理的小鼠產生了氧化應激反應。近些年來,中藥白術中的主要活性物質白術多糖被證明有抗氧化、抗炎及免疫調節等作用,如陳浩祥等[19]的研究發現,對嶺南黃雞飼喂白術多糖可顯著提高血清中SOD 與GSH-Px 的活性,降低機體MDA 含量,從而緩解CTX 引起的機體氧化應激與肝細胞凋亡情況。上述結果表明了PAMK 能增強CTX 處理下的抗氧化能力,減少脂質過氧化程度,緩解由CTX 引起的氧化應激。
為了探究PMAK 對CTX 誘導的腎臟損傷的基因調控機制,本研究進行了轉錄組測序分析。研究結果顯示,存在大量與抗氧化相關的DEGs,其中BCL6、Mfsd2a在本實驗的轉錄組測序數據中有顯著性差異。BCL6的表達水平相對于CTX 組顯著上調,這種上調的程度與CTX 誘導的小鼠氧化應激的減輕呈正相關,同時PAMK 具有抗氧化的功能,BCL6的上調可能與PAMK 抗氧化應激反應的激活相關,從而減輕了CTX 誘導的氧化應激損傷。并且,有研究發現BCL6通過負調控NLRP3轉錄來減輕血管緊張素 II 或脂多糖誘導的人腎小管上皮細胞炎癥,BCL6在自發性高血壓大鼠中的過表達降低了SHR 的血壓、NLRP3的表達和腎皮質炎癥[20]。另外,有研究表明,主要Mfsd2a的下調可能通過抑制Caveola 介導的細胞內轉運,維持血腦屏障的完整性和抗氧化功能,從而保護中樞神經系統的正常功從而維持了血腦屏障的完整性和抗氧化功能[21]。本實驗的轉錄組測序結果與該研究一致,進一步支持了Mfsd2a下調有助于PAMK 緩解CTX 誘導的小鼠氧化應激反應。綜上,表明PAMK 通過調節抗氧化途徑來發揮其保護作用,從而減輕小鼠腎臟氧化損傷。
為了進一步探究PAMK 緩解CTX 誘導的腎損傷的機制,本研究進一步將兩組DEGs 進行富集分析發現,兩組交集通路中最顯著的是花生四烯酸代謝通路,其中花生四烯酸代謝通路已經有研究發現與炎癥發生與緩解密切相關。花生四烯酸(AA)作為細胞膜脂質的主要成分,是細胞膜脂含量的主要組成部分,主要由三種酶代謝:環氧合酶、脂氧合酶和細胞色素P450(CYP450)酶[22]。基于這三種代謝途徑,AA 可以轉化為各種代謝產物,引發不同的炎癥反應。在腎臟中,前列腺素、血栓素、白三烯和羥基二碳四烯酸(HETEs)是AA 產生的主要代謝物。前列腺素和白三烯水平升高導致腎臟炎癥損傷[23-25]。花生四烯酸可以通過花生四烯酸5-氧化酶的催化轉化為5-羥基花生四烯酸,然后5-羥基花生四烯酸可以進一步轉化為白三烯,白三烯是一類強炎癥介質,能夠引起血管擴張、增加血管通透性和促進炎癥細胞的吸附和遷移[26]。PTGS1(也稱為COX-1)是一種環氧合酶酶類,它參與花生四烯酸的代謝,催化花生四烯酸轉化為前列腺素H2 的反應,PTGS1可能導致前列腺素和血栓素的過量產生,引發炎癥反應和相關病理變化[27]。實時熒光定量結果顯示,PTGSI、ALOX5和ALOX12CTX 組中mRNA 的表達量均呈現增高的趨勢。表明了CTX 可能通過氧合酶和脂氧合酶途徑誘導小鼠腎臟炎癥損傷。此外,NF-κB是調控機體炎癥反應的關鍵性轉錄因子,有研究發現缺血再灌注所致急性腎損傷大鼠的腎組織NF-κB 信號通路相關基因NF-κB表達量顯著上升[28]。提示CTX 可能激活腎臟內的NF-κB通路導致小鼠腎臟損傷,上述結果證實,在PAMK+CTX 組中PTGS1、AOLX5、ALOX12、NF-κB的mRNA 表達量均有下降的趨勢,PAMK 在CTX 誘導的腎損傷中發揮了關鍵的腎臟保護作用。
此外,AA 還可以通過細胞色素P450 單加氧酶產生的19-HETE 和20-HETE 調節腎離子運輸;由CYP450 酶產生的環氧二十碳三烯酸(EETs)也在炎癥過程中腎臟損傷中起著重要作用,CYP450 途徑產生的EETs 能夠促進腎小管細胞的增殖和分化,促進腎損傷后的修復。研究發現,EETs 還具有抗炎和抗氧化作用,可以減輕腎損傷引起的炎癥和氧化應激反應,有助于保護腎臟免受進一步的損傷用[29-31]。并且,顯著富集通路中的DEGs,大多參與CYP450 通路,通過對腎臟的轉錄組分析,共同調控基因可能是PAMK 的調控靶點,兩組相關的關鍵基因Cyp2c65、Cyp2b9mRNA表達量呈相反的趨勢與測序結果一致。Cyp2b9是細胞色素P450 家族的一員,眾所周知,它通過產生超氧化物自由基而產生氧化應激[32]。Lu 等[33]研究結果表明,肝臟在HFD(高脂飲食)的作用與Cyp2b家族相關。此外,研究發現LPS 誘導肝臟和十二指腸中Cyp2c、Cyp2j總含量的強烈抑制;同樣處理條件下,在十二指腸,Cyp2c65和Cyp2j6的表達顯著減少[34]。這與上述實驗結果一致,CTX 組Cyp2c65表達量顯著減少,PAMK+CTX組表達量顯著上升,說明了PAMK 可以通過Cyp2c65、Cyp2b9調控花生四烯酸代謝通路影響代謝性產物,緩解CTX 誘導的小鼠腎損傷。P450 酶具有許多生理相關功能,包括調節心臟血管張力[35-36],腎臟離子轉運介導[37],血管抗炎特性[38],以及控制胰腺肽激素的分泌[39]。CYP2J 成員已被證明在AA 和亞油酸以及各種外源性藥物的代謝中具有活性[40-41]。有研究發現CYP2J 酶可能參與脂肪酸代謝,并可能影響許多心血管和腎臟代謝過程[42-43]。同時PAMK 具有調節脂代謝酶的活性,提高抗氧化能力,PAMK 對CTX 所致的炎癥和氧化應激有正向的調節作用[44]。另外,細胞色素P450 途徑產生的EETs 加快對腎臟損傷細胞的增殖分化,促進血管新生,恢復腎臟基本功能,及時將代謝廢物和有害物質排出體外[45]。以上結果表明,在CTX 誘導的小鼠腎損傷過程中,PAMK可能通過影響花生四烯酸環氧合酶的活性緩解小鼠腎臟損傷。
綜上所述,本研究結果表明PAMK 可能通過調節花生四烯酸代謝通路,緩解小鼠腎臟的氧化應激反應,從而降低CTX 引起的腎臟損傷。這一發現揭示了PAMK 在維持腎臟形態和結構完整性方面的重要作用,這些結果為進一步探索PAMK 的調控機制以及開發相關治療方法提供參考依據。
? The Author(s) 2024.This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學苑創造·A版(2020年9期)2020-10-13 09:41:02
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
食品科學(2013年15期)2013-03-11 18:25:48
