我國華南中麝鼩分子系統地理學分析
2024-06-29 00:00:00欒天琪郭秋穎高藝韓美鳳張春鳳陳歡蔡赫金志民張雋晟劉鑄
野生動物學報 2024年2期

摘 要 華南中麝鼩(Crocidura rapax)亞種分化存在異議。為了明確我國華南中麝鼩的分子系統地理格局,為其分類學提供進一步分子生物學證據,測定了我國貴州華南中麝鼩20個樣本的Cyt b 基因序列,并結合GenBank下載的19個華南中麝鼩Cyt b 基因序列進行一并分析。結果表明:系統發生樹首先分成3個大的進化支,即華南中麝鼩華南亞種(C. r. rapax)進化支、西南中麝鼩(C. vorax)進化支、華南中麝鼩臺中亞種(C. rapax kurodai)進化支。華南中麝鼩華南亞種分成貴州、云南、四川東南部譜系和四川中西部譜系。遺傳距離結果發現,最大遺傳距離出現在華南中麝鼩華南亞種四川中西部譜系與華南中麝鼩臺中亞種之間(0. 118 1),其次是華南中麝鼩華南亞種貴州、云南、四川東南部譜系與華南中麝鼩臺中亞種之間(0. 117 4)。華南中麝鼩華南亞種2個譜系間遺傳距離為0. 019 5。中介網絡分析也觀察到相似分化格局。中介網絡圖的結果與系統發育關系和遺傳距離的結果一致,支持來自臺灣的華南中麝鼩樣本為獨立物種的觀點,即臺灣長尾麝鼩(C. tadae)。我國華南中麝鼩無亞種分化,分為貴州、云南、四川東南部譜系和四川中西部譜系。在華南中麝鼩的32個樣本中發現23個單倍型,核苷酸多樣性0. 012 85。種群歷史分析暗示華南中麝鼩近期盡管經歷了種群擴張,但并非為單次急劇擴張。
關鍵詞:華南中麝鼩;麝鼩屬;分子系統地理學
中圖分類號:Q95 文獻標識碼:A 文章編號:2310 - 1490(2024)- 02 - 0291 - 07
DOI:10.12375/ysdwxb.20240207
華南中麝鼩(Crocidura rapax)隸屬勞亞食蟲目(Eulipotyphla)鼩鼱科(Soricidae)麝鼩屬(Crocidura)。華南中麝鼩自發現后,在相當長的一段時間內被認為是C. russula 的一個亞種(C. r. rapax)[1]。Jiang等[2]通過形態學分析認為華南中麝鼩和西南中麝鼩(C.vorax)是兩個獨立的物種,并且兩個物種具有較相似的形態,根據形態學相似度將C. kurodai 歸類于華南中麝鼩。
華南中麝鼩的地理分布和亞種分化存在著明顯的相關性。一部分學者認為在我國華南中麝鼩具有4個亞種:臺中亞種(C. rapax kurodai)分布于臺灣;綠島亞種(C. r. lutaoensis)分布于臺灣東南部的綠島;蘭嶼亞種(C. r. tadae)分布于臺灣東南部的蘭嶼島;華南亞種(C. r. rapax)分布于云南、貴州、四川、廣東、廣西和海南[2?6]。另一部分學者認為分布于臺灣及其周邊島嶼的3 個亞種(C. r. kurodai、C. r. lu?taoensis 和C. r. tadae)屬于另外一個物種,即臺灣長尾麝鼩(C. tadae)的3個亞種(指名亞種C. t. tadae、臺灣亞種C. t. kurodai 和綠島亞種C. t. lutaoen?sis)[7?8],在我國華南中麝鼩無亞種分化,分布于云南、貴州、四川、廣東、廣西和海南[7?9]。可見,華南中麝鼩的亞種分化存在異議。
系統地理學是研究物種間及物種內不同類群形成現有分布格局的歷史原因和演化過程的一門科學[10],分子系統地理學研究為動物分類地位的確定提供了從物種水平到系統發育水平的證據,進而為準確地評價動物物種分布格局奠定良好基礎[11]。鑒于目前缺乏對華南中麝鼩的分子系統地理學系統分析,本研究采集并測序了來自我國貴州的華南中麝鼩Cyt b 基因全序列,結合已有報道的我國臺灣、四川和云南的華南中麝鼩Cyt b 基因全序列(Gen?Bank),分析我國華南中麝鼩的分子系統地理格局,進一步為分類學提供分子生物學證據。
1 研究方法
1. 1 樣本采集
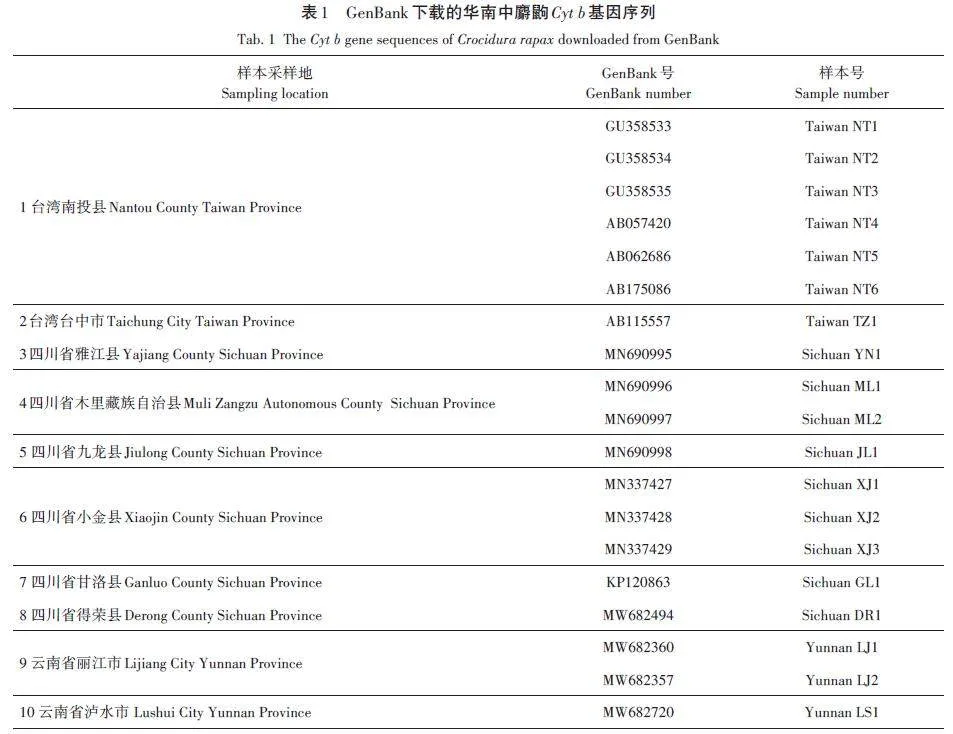
采用陷阱法,輔以鋏捕法采集華南中麝鼩樣本。樣本采集于我國貴州省,采集時間為2019年1月—2021年10月,共采集樣本20個,其中安順地區(26°21'48″ N,105°55'35″ E)樣本5個(Guizhou AS1~Gui?zhou AS5),畢節地區(26°24'22″ N,105°44'4″ E)樣本13 個(Guizhou BJ1~Guizhou BJ13),六盤水地區(26°33'1″ N,104°57'18″ E)樣本2個(Guizhou LPS1,Guizhou LPS2)。從GenBank下載了其他地區華南中麝鼩的Cyt b 基因全序列(19個),基本信息見表1。本研究通過牡丹江師范學院實驗動物倫理委員會審查(IACUC-MNU-0-401)。
1. 2 試驗方法
用剪刀剪取樣本腿部肌肉組織,將腿部肌肉組織用酚-氯仿法進行基因組DNA提取。mtDNA的Cyt b基因全序列PCR 反應體系為50 μL,含200 μmol /L的dNTP,1. 5 mmol/L 的MgCl2,Primer(L14724/H15915)[12]各10 pmol/L,Taq DNA polymerase 2. 5 U(大連寶生物),模板為250 ng。PCR擴增的反應條件:94 ℃預變性3 min;94 ℃變性45 s,50 ℃退火50 s,72 ℃延伸45 s,共30個循環;72 ℃延伸7 min,4 ℃保存。PCR擴增產物經純化回收進行測序(哈爾濱博仕生物技術有限公司)。
1. 3 數據處理
使用EditSeq(DNAStar 軟件包)對試驗獲得mtDNA Cyt b 基因全序列和GenBank下載華南中麝鼩的序列(表1)進行序列編輯,再利用Clustal W軟件進行序列聚類比對[13]。利用DnaSP 5. 10分析單倍型數量(number of haplotypes,k)、多態位點數量(number of polymorphic sites,s)、單倍型多態性(hap?lotype diversity,h)、核苷酸差異平均數(average num?ber of nucleotide differences,i)、核苷酸多樣性(nucleotide diversity,π),計算Tajima’s D 和Fu’s 檢測中性選擇情況,進行顯著性檢測,并結合種群的增長模型(a population growth-decline model)分析錯配分布來檢測種群數量擴張情況[14]。利用MEGA5. 05 軟件,采用最大似然法(maximum likelihood,ML)構建系統發生樹[15]。利用Network 4. 6 構建中介網絡圖(median-joining network)分析單倍型間進化關系[16]。
2 結果
2. 1 系統發育關系
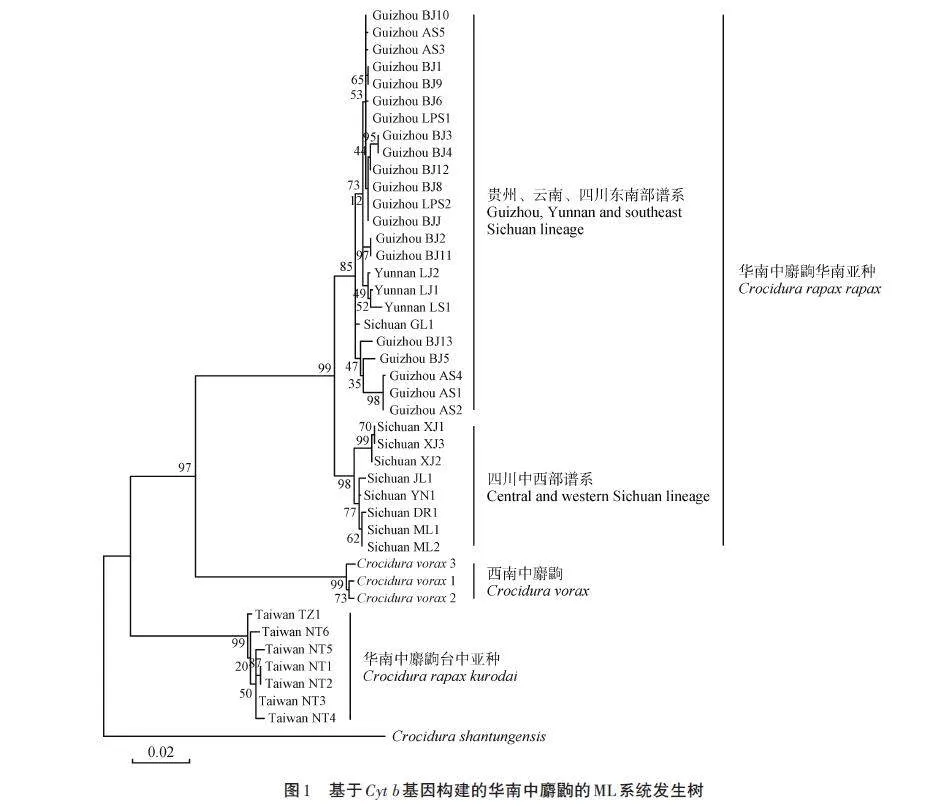
以西南中麝鼩和山東小麝鼩(C. shantungensis)為外群,利用ML法構建系統發生樹(圖1)。系統發生樹首先分成3個大的進化支:一是華南中麝鼩華南亞種進化支;二是西南中麝鼩進化支;三是華南中麝鼩臺中亞種進化支。西南中麝鼩進化支夾在華南中麝鼩華南亞種進化支和華南中麝鼩臺中亞種進化支中間,表現出與華南中麝鼩華南亞種具有較近的親緣關系。本研究的系統發育關系結果支持來自臺灣的華南中麝鼩樣本為獨立物種的觀點,即臺灣長尾麝鼩。將華南中麝鼩華南亞種進化支以高支持率(99%)分成兩個進化譜系,一個是由貴州所有樣本、云南所有樣本和來自四川東南部甘洛縣的樣本構成的貴州、云南、四川東南譜系,另一個是由來自四川中部和西部的樣本構成的四川中西部譜系,可見兩個譜系分化較為明顯。
2. 2 遺傳距離
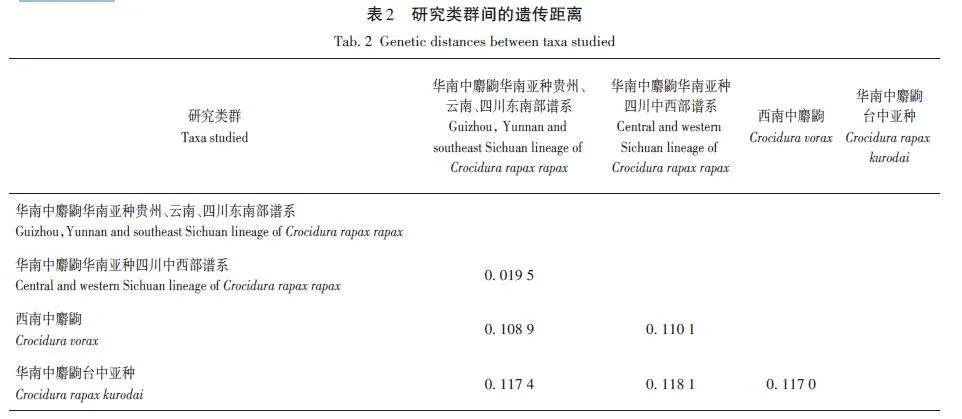
計算了華南中麝鼩華南亞種的2個譜系與西南中麝鼩和華南中麝鼩臺中亞種的遺傳距離(表2)。華南中麝鼩華南亞種的2個譜系間的遺傳距離最小(0. 019 5),最大的遺傳距離出現在華南中麝鼩華南亞種四川中西部譜系與華南中麝鼩臺中亞種之間(0. 118 1),其次是華南中麝鼩華南亞種貴州、云南、四川東南部譜系與華南中麝鼩臺中亞種之間(0. 117 4),可見華南中麝鼩華南亞種與華南中麝鼩臺中亞種之間的遺傳距離較大,超過了西南中麝鼩與兩個亞種之間的遺傳距離。遺傳距離結果與系統發育關系結果一致,支持來自臺灣的華南中麝鼩樣本為獨立物種的觀點,即臺灣長尾麝鼩。
2. 3 中介網絡分析
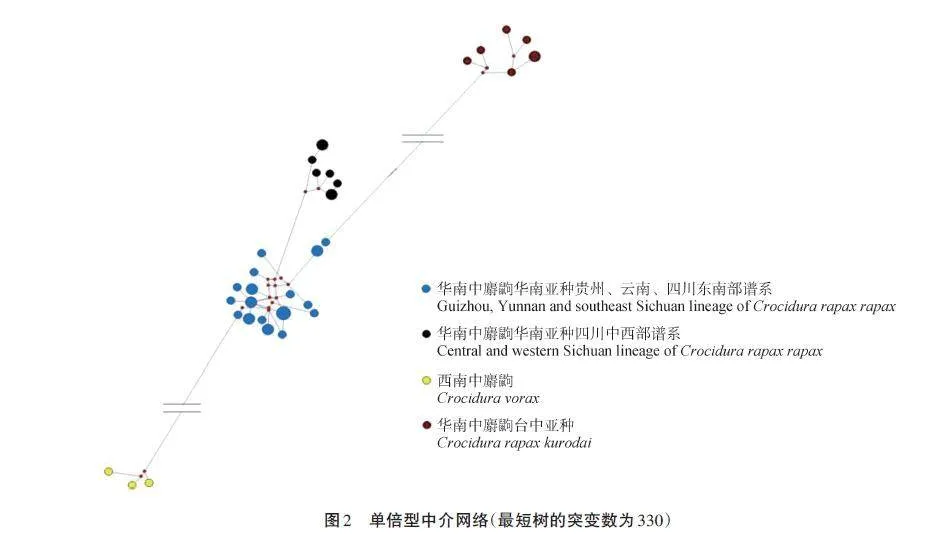
中介網絡圖(圖2)顯示出與系統發育關系類似的結果。華南中麝鼩華南亞種貴州、云南和四川東南部譜系的單倍型位于中介網絡圖的中部,各單倍型間表現出較近的網絡關系。華南中麝鼩華南亞種四川中西部譜系位于中介網絡圖的中部右上方,各單倍型間表現出較近的網絡關系,都來自于一個突變節點。西南中麝鼩的單倍型位于中介網絡的下方,表現出與華南中麝鼩華南亞種具有較遠的網絡關系。華南中麝鼩臺中亞種的單倍型位于中介網絡圖的最上方,表現出與華南中麝鼩華南亞種具有較遠的網絡關系。中介網絡圖的結果與系統發育關系和遺傳距離的結果一致,支持來自于臺灣的華南中麝鼩樣本為獨立物種的觀點,即臺灣長尾麝鼩。
2. 4 遺傳多樣性和歷史種群動態
鑒于系統發育關系、遺傳距離和中介網絡分析的結果都支持來自臺灣的華南中麝鼩樣本為獨立物種的觀點,即臺灣長尾麝鼩。本研究只對華南中麝鼩華南亞種包含的樣本(32個)進行了遺傳多樣性和歷史種群動態分析。華南中麝鼩的Cyt b 基因全序列為1 140 bp,4 種核苷酸堿基含量的平均值分別為A=29. 6%,T=31. 0%,G=13. 2%,C=26. 2%,其中T的含量最高,G 的含量最低,A+T的含量為60. 6%,明顯高于G+C 的含量(39. 4%)。序列包含68 個變異位點,為45 個簡約信息位點(parsimony informativesites)和23個單變異位點(singleton variable sites),變異位點數占總位點數的5. 97%,未發現插入和缺失。華南中麝鼩的遺傳多樣性參數見表3。在華南中麝鼩的32個樣本中發現23個單倍型,核苷酸多樣性0. 012 85。
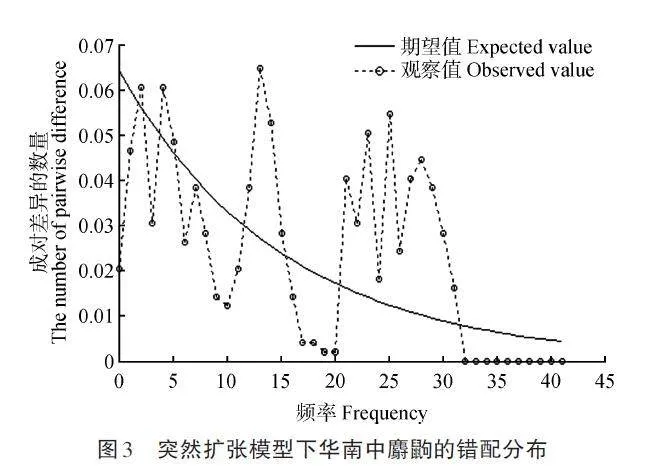
華南中麝鼩的Tajima’s D 結果(D=-0. 542 1,pgt;0. 100)盡管不顯著,但為負值,Fu’s FS 結果(FS=-3. 771 0,p=0. 014)為顯著性負值(表3),中性檢測結果顯示華南中麝鼩近期可能發生過種群擴張。采用錯配分布方法對種群歷史進行檢驗,構建的華南中麝鼩錯配曲線是多峰曲線(圖3),說明華南中麝鼩近期盡管經歷了種群擴張,但并非為單次急劇擴張。
3 討論
Tokuda等[17]在我國臺灣蘭嶼島發現了臺灣長尾麝鼩。Kuroda 將臺灣長尾麝鼩作為南小麝鼩(C.horsfieldii)的一個亞種,并且說明它在臺灣主島沒有分布[18]。Jameson等[19]在臺灣主島上發現了南小麝鼩的另外一個亞種(C. h. kurodai)。Fang 等[20]根據形態學和核型分析,發現C. kurodai 與南小麝鼩存在差異,認為C. kurodai 為獨立物種。Fang等[18]再次利用形態學和核型分析,認為臺灣長尾麝鼩為有效物種,包括3個亞種:C. tadae kurodai、C. t. tadae、C. t.lutaoensis。近些年,還有很多文獻將這些分布于臺灣及其周邊島嶼的麝鼩歸類于華南中麝鼩:臺中亞種(C. r. kurodai)、綠島亞種(C. r. lutaoensis)、蘭嶼亞種(C. r. tadae)[2?4,6]。本研究的系統發育關系、遺傳距離和中介網絡分析的結果都支持已有文獻中認為來自臺灣的華南中麝鼩樣本為獨立物種的觀點,即臺灣長尾麝鼩。本研究的結果支持已有文獻認為臺灣長尾麝鼩為獨立物種的觀點,華南中麝鼩在我國沒有亞種分化[7?9]。
華南中麝鼩被分成兩個譜系,即華南中麝鼩貴州、云南、四川東南部譜系和華南中麝鼩四川中西部譜系。由于四川省由北向南的水系較多,推測是水系起到了一定的天然屏障阻隔作用使得華南中麝鼩被分成兩個譜系。本研究未能獲得我國廣東、廣西和海南的樣本,這些地域華南中麝鼩的分子系統地理學問題有待進一步研究。
參考文獻:
[1] ELLERMAN J R, MORRISON-SCOTT T C S. Checklist of Pa?laearctic and Indian Mammals 1758 to 1946[M]. London: theTrustees of the British Museum, 1951:81.
[2] JIANG X L, HOFFMANN R S. A revision of the white-toothedshrews (Crocidura) of southern China[J]. Journal of Mammal?ogy, 2001, 82(4): 1059-1079.
[3] HUTTERER R. Order Soricomorpha[M]//WILSON D E, REEDERD A. Mammal species of the world: a taxonomic and geo?graphic reference. 3rd ed. Baltimore: Johns Hopkins UniversityPress, 2005: 249.
[4] SMITH A T, 解焱. 中國獸類野外手冊[M]. GEMMA F, 繪.陳延熹,楊戎生,張勁碩,等譯. 長沙: 湖南教育出版社, 2009:227-228.
SMITH A T, XIE Y. A guide to the mammals of China[M].GEMMA F, illus. CHEN Y X,YANG R S, ZHANG J S, et al.trans. Changsha: Hunan Education Publishing House, 2009:227-228.
[5] 蔣志剛, 馬勇, 吳毅, 等. 中國哺乳動物多樣性及地理分布[M]. 北京: 科學出版社, 2015:67; 69.
JIANG Z G, MA Y, WU Y, et al. China’s mammal diversity andgeographic distribution[M]. Beijing: Science Press, 2015:67; 69.
[6] BURGIN C J, HE K. Family Soricidae (shrews)[M]//WILSON DE, MITTERMEIER R A. Handbook of the mammals of the world:Vol. 8: insectivores, sloths and colugos[M]. Barcelona: LynxEditions, 2018: 485.
[7] 魏輔文, 楊奇森, 吳毅, 等. 中國獸類分類與分布[M]. 北京:科學出版社, 2022: 268.
WEI F W,YANG Q S,WU Y, et al. Taxonomy and distribution ofmammals in China[M]. Beijing: Science Press, 2022: 268.
[8] 王應祥. 中國哺乳動物種和亞種分類名錄與分布大全[M]. 北京: 中國林業出版社, 2003: 23-24.
WANG Y X. A complete checklist of mammal species and subspe?cies in China a taxonomic and geographic reference[M]. Beijing:China Forestry Publishing House, 2003:23-24.
[9] 譚邦杰. 哺乳動物分類名錄[M]. 北京: 中國醫藥科技出版社, 1992: 43.
TAN B J. A systematic list of the mammals[M]. Beijing: ChinaMedical Science Press,1992: 43.
[10] AVISE J C, ARNOLD J, BALL R M, et al. Intraspecific phylo?geography: the mitochondrial DNA bridge between population ge?netics and systematics[J]. Annual Review of Ecology and Sys?tematics, 1987, 18: 489-522.
[11] 劉鑄, 徐艷春, 戎可, 等. 嚙齒動物分子系統地理學研究進展[J]. 生態學報, 2014, 34(2): 307-315.
LIU Z, XU Y C, RONG K, et al. The current progress in ro?dents molecular phylogeography[J]. Acta Ecologica Sinica,2014, 34(2): 307-315.
[12] IRWIN D M, KOCHER T D, WILSON A C. Evolution of the cy?tochrome b gene of mammals[J]. Journal of Molecular Evolu?tion, 1991, 32(2): 128-144.
[13] THOMPSON J D, GIBSON T J, PLEWNIAK F, et al. TheCLUSTAL_X windows interface: flexible strategies for multiplesequence alignment aided by quality analysis tools[J]. NucleicAcids Research, 1997, 25(24): 4876-4882.
[14] LIBRADO P, ROZAS J. DnaSP v5: a software for comprehen?sive analysis of DNA polymorphism data[J]. Bioinformatics,2009, 25(11): 1451-1452.
[15] TAMURA K, PETERSON D, PETERSON N, et al. MEGA5:molecular evolutionary genetics analysis using maximum likeli?hood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology and Evolution, 2011, 28(10): 2731-2739.
[16] BANDELT H J, FORSTER P, R?HL A. Median-joining net?works for inferring intraspecific phylogenies[J]. Molecular Biol?ogy and Evolution, 1999, 16(1): 37-48.
[17] TOKUDA M, KANO T. A bat and a new shrew from K?t?-sh?(Botel-Tobago)[J]. Annotationes Zoologicae Japonenses, 1936,15(4): 427-432.
[18] FANG Y P, LEE L L. Re-evaluation of the Taiwanese whitetoothedshrew, Crocidura tadae Tokuda and Kano, 1936 (Insec?tivora: Soricidae) from Taiwan and two offshore islands[J]. Jour?nal of Zoology, 2002, 257(2): 145-154.
[19] JAMESON E W, JONES G S. The Soricidae of Taiwan[J]. Pro?ceedings of the Biological Society of Washington, 1977, 90:459-482.
[20] FANG Y P, LEE L L, YEW F H, et al. Systematics of whitetoothedshrews (Crocidura)(Mammalia: Insectivora: Soricidae)of Taiwan: karyological and morphological studies[J]. Journal ofZoology, 1997, 242(1): 151-166.
基金項目:牡丹江師范學院科研項目(GP2022009,HX2023004);黑龍江省省屬高等學校基本科研業務費科研項目(1452TD008,1451PT008,1451TD002);黑龍江省自然基金項目(LH2021C095)
