海水中降解幾丁質細菌的篩選鑒定及培養條件優化
2024-09-03 00:00:00祁紅兵張敏怡
安徽農業科學 2024年16期


摘要 [目的]從海水中獲得高產幾丁質酶菌株并優化培養條件。[方法]采集湛江市金沙灣海水作為樣品,以膠體幾丁質為唯一碳源篩選分離產幾丁質酶菌株,DNS法測定酶活力并進行分子鑒定;單因素試驗和正交優化其產酶最佳條件。[結果]初篩得到6株能降解幾丁質的細菌,其中編號為Z-4的菌株幾丁質酶活力最強,形態鑒別及16S rDNA序列分析鑒定為嗜麥芽窄食單胞菌(Stenotrophomonas maltophilia)。菌株Z-4優化后的最佳產酶條件為可溶性淀粉添加量1.5%、蛋白胨添加量2.5%、KHPO添加量0.04%,在30 ℃、pH 7.0、接種量2%、160 r/min、培養72 h,酶活力達到0.452 U/mL,相比優化前提高了5倍。[結論]研究結果為海洋副產物有效利用幾丁質改善環境提供重要參考。
關鍵詞 海水;細菌;幾丁質酶;培養條件優化;篩選鑒定
中圖分類號 S 917.1 文獻標識碼 A
文章編號 0517-6611(2024)16-0001-05
doi:10.3969/j.issn.0517-6611.2024.16.001
開放科學(資源服務)標識碼(OSID):
Screening and Identification of Chitin Degrading Bacteria in Seawater and Optimizing Culture Conditions
QI Hong-bing,ZHANG Min-yi
(School of Life Science and Technology,Lingnan Normal University,Zhanjiang,Guangdong 524048)
Abstract [Objective] To obtain high-yield chitinase strains from seawater and optimize culture conditions.[Method]The seawater from Jinsha Bay of Zhanjiang City was collected as samples,chitinase producing strains were isolated using colloidal chitin as the only carbon source,the enzyme activity was measured by DNS method and molecular identification;single factor experiment and orthogonal optimization were used to optimize the optimal conditions for enzyme production.[Result]Six strains of chitinase-degrading bacteria were screened.The strain with the strongest chitinase activity was Z-4.The strain was identified as Stenotrophomonas maltophilia by morphological observation and 16S rDNA sequence analysis.The optimum conditions for producing enzyme of strain Z-4 were obtained as follows:soluble starch addition of 1.5%,peptone addition of 2.5%,KHPO addition of 0.04%;at 30 ℃,pH 7.0,inoculation amount of 2%,160 r/min and 72 hours of cultivation,the enzyme activity reached 0.452 U/mL,which was 5 times higher than before optimization.[Conclusion]The research results provide an important reference for the effective use of chitin in the ocean to improve the environment.
Key words Seawater;Bacteria;Chitinase;Optimization of culture conditions;Screening and identification
基金項目
湛江市非資助科技攻關計劃項目(2023B01023);嶺南師范學院校級教育教學改革項目 (嶺師教務〔2022〕154號)。
作者簡介 祁紅兵(1971— ),男,河南信陽人,副教授,博士,從事微生物生理研究。
收稿日期 2023-09-13
幾丁質是世界上含量巨大的生物多聚體之一,其廣泛存在許多物種的體內和外殼中,如線蟲的表皮、真菌的細胞壁、節肢動物的外骨架以及許多昆蟲的腸道[1]。幾丁質是一種多糖,在生物體內并不是以游離的狀態存在的,而是與其他物質結合在一起,因此幾丁質的開發利用需要對含幾丁質的原料進行處理,分離出幾丁質[1]。目前主要通過化學法對幾丁質原料進行降解,從而產生殼聚糖、幾丁質寡糖及其衍生物,但這種方法容易造成環境污染,與國家倡導的綠色發展理念相悖[2-3]。幾丁質的降解產物具有調節血糖、抗腫瘤、降血脂、抗菌等作用,目前已被應用于醫藥、化工、食品、環保等領域[4]。幾丁質酶是指能夠催化水解幾丁質的酶類總稱,在自然界幾乎所有生物類群中都能發現幾丁質酶的存在[5]。微生物幾丁質酶在農業、工業、食品、醫藥等領域展現出良好前景[6]。
地球表面積的3/4約為海洋,由于海洋生物資源數不勝數,各海洋大國紛紛把目光投向了海洋生物資源的研究開發上,其中海洋生物酶的研究便是各國的重點研究對象之一[7-9]。但自然界中產幾丁質酶的微生物普遍存在產酶量低、生產成本過高、酶活力低等問題,使得在實際中的應用并不多。因此,加大對海洋生物資源的研究,從海洋微生物中分離高產幾丁質酶的菌株具有重大的意義。
1 材料與方法
1.1 試驗材料
1.1.1 試驗菌株。
湛江金沙灣有不少貝類、蝦、蟹等,每年都會產生較多的殼類廢棄物,為幾丁質酶產生菌提供了適宜的生長和繁殖條件,因此以湛江金沙灣采集的海水作為樣品,從中分離篩選能產幾丁質酶的細菌。
1.1.2 培養基。
平板分離培養基:蛋白胨0.500%,七水硫酸鎂0.001%,七水硫酸鋅0.001%,膠體幾丁質0.500%,磷酸二氫鉀0.005%,瓊脂2.000%,pH 7.0。種子培養基:蛋白胨0.500%,七水硫酸鎂0.001%,七水硫酸鋅0.001%,膠體幾丁質0.500%,磷酸二氫鉀0.005%,pH 7.0。發酵培養基:蛋白胨0.500%,七水硫酸鎂0.001%,七水硫酸鋅0.001%,膠體幾丁質0.500%,磷酸二氫鉀0.005%,pH 7.0。
1.1.3 主要試劑。
N-乙酰-D-氨基葡萄糖(NAG),分析純,潤友化學(深圳)有限公司;甲殼素,分析純,上海麥克林生化f6c4da06c6ff31cecc610fb609e2a03c科技有限公司;3,5-二硝基水楊酸(DNS),武漢卡諾斯科技有限公司;細菌基因組DNA提取試劑盒,北京索萊寶科技有限公司;PCR引物,英濰捷基(上海)貿易有限公司。
1.1.4 主要儀器。
生化培養箱(SPL-150,廣州瑞彬科技有限公司) ;紫外可見分光光度計(752N,上海儀電分析儀器有限公司);凝膠成像儀(ChampChemi Professional,北京三江銳志科技有限公司);電泳儀(DYY-6C,成都一科儀器設備有限公司);紫外投射反射儀(WFH-201B,上海精科實業有限公司);基因擴增儀器(A300,杭州朗基科學儀器有限公司)。
1.2 試驗方法
1.2.1 膠體幾丁質的制備。
精確稱量細粉幾丁質5 g,加入100 mL濃鹽酸混勻,放置24 h,邊攪拌邊過濾到500 mL的蒸餾水中,靜置至沉淀析出,離心后收集膠體幾丁質,蒸餾水沖洗至中性,用0.2 mmol/L磷酸緩沖液定容,使膠體幾丁質溶液的最終濃度為1%[10-11]。
1.2.2 DNS溶液的制備。
參照付星等[12]的方法,稱取3,5-二硝基水楊酸6.3 g,加入500 mL蒸餾水,攪拌并加熱使其溶解,逐步加入300 mL 10% NaOH溶液,同時不斷攪拌,按順序依次加入182 g酒石酸鉀鈉、5 g結晶酚和5 g NaSO,攪拌并加熱使其溶解(溫度不宜超過50 ℃),待溶液冷卻至室溫后,定容至1 L,棕色瓶中儲存備用,室溫下放置7 d后使用。
1.2.3 N-乙酰-D-氨基葡萄糖(NAG)標準曲線的繪制。
1.2.3.1 NAG標準溶液的配制。用適量蒸餾水將100 mg的NAG溶解,定容至100 mL,配制成濃度為0.1%的NAG標準溶液,并置于4 ℃冰箱保存。
1.2.3.2 標準曲線的繪制。依次將NAG標準溶液、蒸餾水和DNS溶液加入試管中,混勻后,煮沸5 min。迅速冷卻后,定容至25 mL,以不加NAG標準液的0號管為對照調零,用紫外可見分光光度計在540 nm處測定吸光度。以吸光度為縱坐標、NAG含量為橫坐標繪制標準曲線(圖1),得出線性回歸方程y=0.328 8x-0.009 1(R2=0.993 4)。
1.2.4 幾丁質酶活力的測定。將發酵液在6 000 r/min離心15 min,取上清液0.5 mL加入1 mL的1%膠體幾丁質,于40 ℃中反應30 min,以滅活的上清液作為對照,加入1.5 mL蒸餾水和1.5 mL DNS,充分混勻,在沸水浴中加熱5 min,迅速用流水冷卻至室溫,定容至25 mL,在540 nm測吸光度[13-14],并根據NAG標準曲線線性回歸方程計算出NAG含量。酶活力計算公式如下:
酶活力(U/mL)=m×n×1 000/(M×T×V)
式中:m為NAG含量;n為稀釋倍數;M為NAG相對分子量(221.21);T為反應時間;V為酶液體積。
1.2.5 產幾丁質酶菌株的分離純化。
1.2.5.1 初篩。
取1 mL樣品加入裝有9 mL無菌海水的試管中,混勻,分別稀釋制成不同倍數的稀釋液。從10-4~10-7 的4管海水稀釋液中各吸取0.1 mL,均勻地涂布在平板分離培養基上,在30 ℃條件下培養96 h[15]。
1.2.5.2 復篩。
從初篩平板中挑取能夠產生透明圈的菌株,進行劃線分離純化。取劃線后長出的單菌落接入到種子培養基中,30 ℃、搖床160 r/min振蕩培養24 h,取種子液0.6 mL 接入30 mL發酵培養基中,同樣條件下振蕩培養5 d,取發酵液在6 000 r/min的條件下離心15 min,收集上清粗酶液,測定酶活力,斜面培養基中保存酶活力較高的菌株。
1.2.6 菌株的形態觀察。
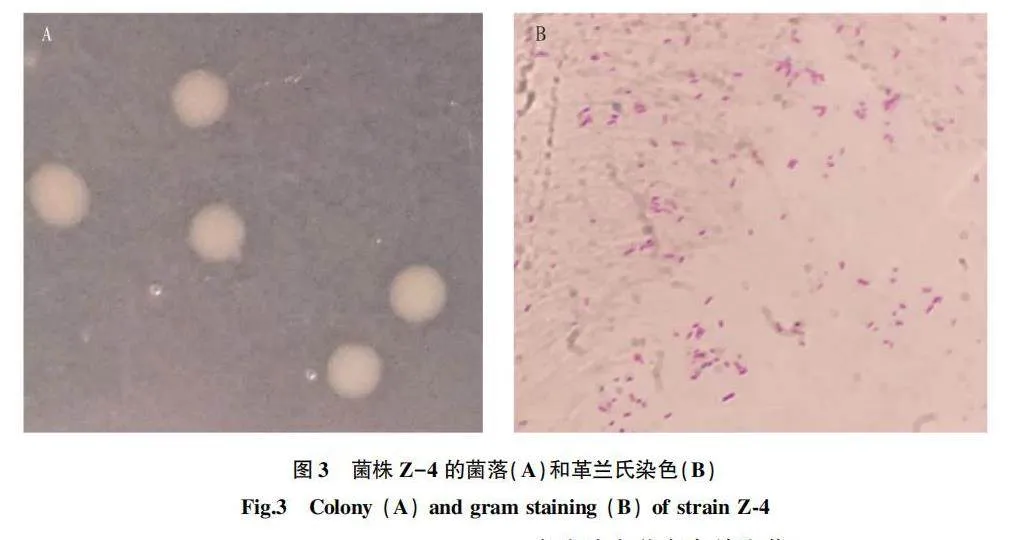
將菌株劃線于2216E瓊脂平板,30 ℃培養72 h,期間觀察菌落形態及革蘭氏染色情況。
1.2.7 菌株的分子鑒定。
1.2.7.1 基因組DNA的提取。按細菌基因組DNA提取試劑盒說明書提取總DNA后,用1.0%瓊脂糖凝膠進行電泳。
1.2.7.2 16S rDNA基因擴增與測序。
用于PCR擴增的引物為27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-TACCTTGTTACGACTT-3′),由英濰捷基(上海)貿易有限公司合成。PCR反應體系為30 μL。PCR擴增條件:98 ℃預變性2 min;98 ℃變性10 s,58 ℃退火30 s,68 ℃延伸1 min,35個循環;68 ℃充分延伸10 min。
PCR擴增產物用1.0%瓊脂糖凝膠進行電泳,并將擴增產物送往上海立菲生物技術有限公司進行測序。將所測得的16S rDNA基因序列與GenBank數據庫中的已知序列進行BLAST比對分析,采用鄰接法(Neighbor-joining)在MEGA 7.0軟件構建系統發育樹。
1.2.8 菌株生長曲線的繪制。
種子培養基中接種菌株,30 ℃、搖床160 r/min培養,定時取樣,測定波長為600 nm時菌液的OD值,并繪制生長曲線,確定種子液的培養時間。
1.2.9 菌株產幾丁質酶條件的單因素試驗。
在發酵培養基的基礎上,分別考察碳源(細粉幾丁質、膠體幾丁質、葡萄糖、可溶性淀粉)、
碳源濃度(0.5%、1.0%、1.5%、2.0%、2.5%)、氮源(蛋白胨、酵母膏、牛肉膏、硫酸銨)、氮源濃度(1.0%、1.5%、2.0%、2.5%、3.0%)、KHPO濃度(0、0.01%、0.02%、0.03%、0.04%、0.05%)、發酵時間(1、2、3、4、5、6 d)、培養基初始pH(6.0、6.5、7.0、7.5、8.0、8.5)對菌株產酶的影響[16]。
1.2.10 正交試驗。
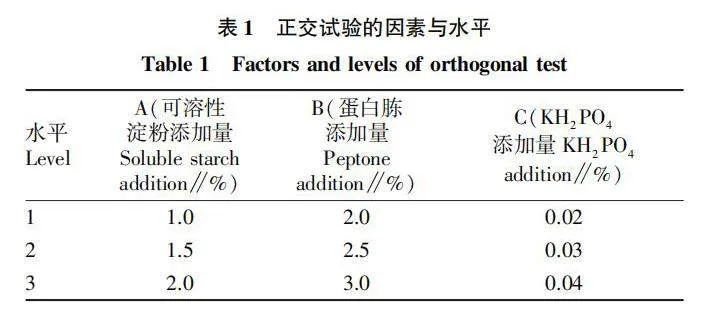
在單因素試驗基礎上,以可溶性淀粉添加量、蛋白胨添加量、KHPO添加量為考察因素,進行L(33)正交試驗,因素和水平的選擇如表1所示。
1.3 數據處理
通過SPSS、Excel進行數據統計并作圖,運用MEGA 7.0軟件構建系統發育樹。
2 結果與分析
2.1 產幾丁質酶菌株的分離純化
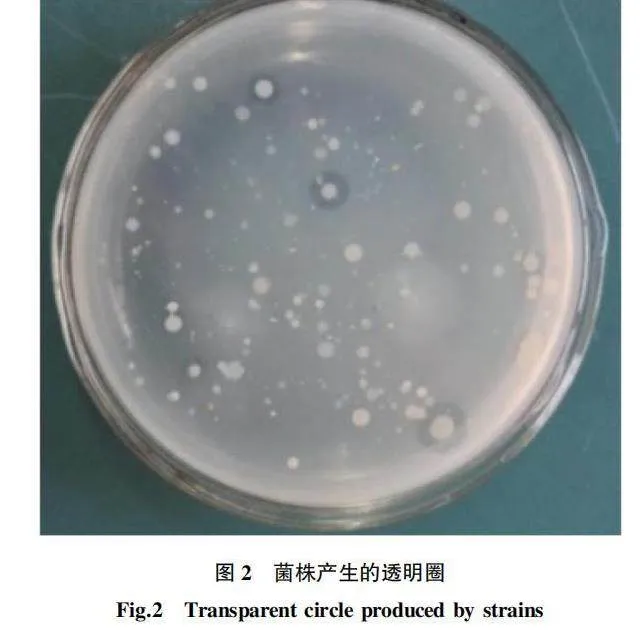
2.1.1 初篩。
培養1 d后平板長出少許菌落,2 d后菌落數明顯增多,開始出現透明圈,3 d后透明圈越來越明顯,挑取能產生透明圈的菌株,進行平板劃線。經初篩得到6株能降解幾丁質的菌株(Z-1、Z-2、Z-3、Z-4、Z-5、Z-6),部分菌株的生長狀況見圖2。
2.1.2 復篩。
為了更精確地判斷菌株的產酶能力,需要對這6株菌株進行復篩,培養5 d后分別測定酶活力,結果發現,Z-1、Z-2、Z-3、Z-4、Z-5、Z-6菌株的酶活力分別為0.017、0.053、0.032、0.075、0.038、0.024 U/mL,可見,這6株細菌都具有產幾丁質酶的能力,但酶活力有差異,其中編號為Z-4的菌株酶活力最高,所以選定菌株Z-4作為試驗菌株。
2.2 菌株的形態特征
菌株Z-4在2216E瓊脂平板培養后,其形態特征為圓形隆起,呈淡黃色,表面光滑,不透明,邊緣平整(圖3 A);革蘭氏染色陰性,多數呈桿狀(圖3 B)。
2.3 菌株的分子鑒定
2.3.1 16S rDNA的PCR擴增。
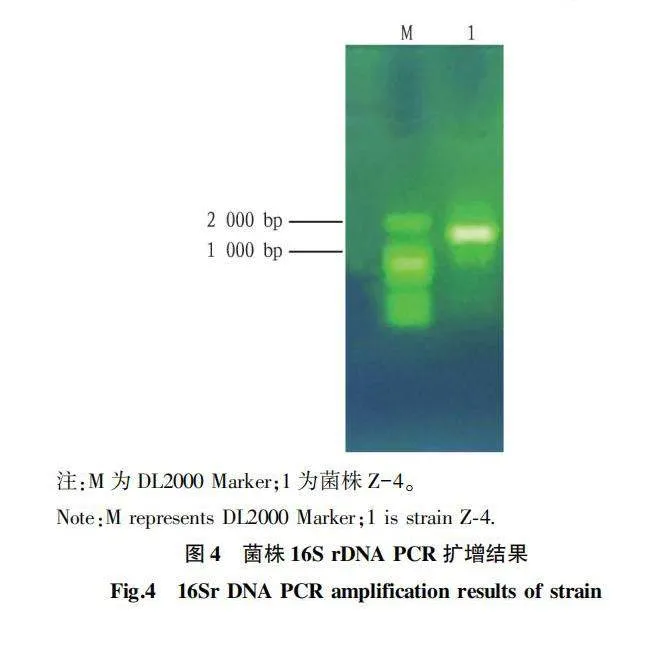
從圖4可以看出,經瓊脂糖凝膠電泳檢測菌株Z-4的PCR擴增產物,有一條大約為1 400 bp 的條帶出現,條帶清晰且明亮。
2.3.2 16S rDNA序列比對與系統發育分析。
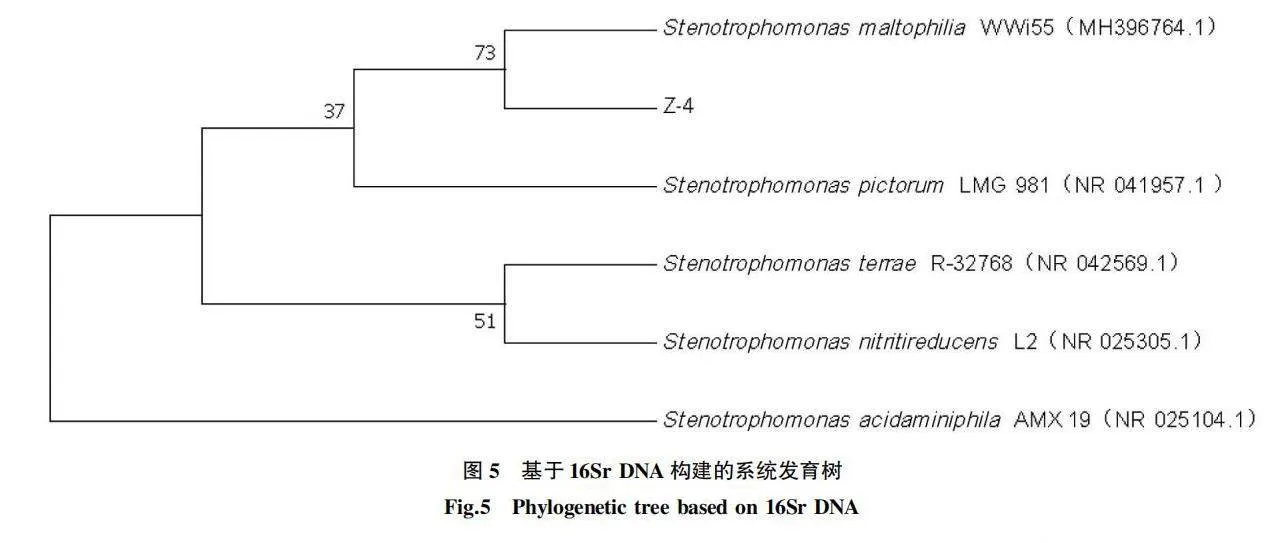
菌株Z-4的16S rDNA序列長度為1 426 bp,在NCBI的GenBank數據庫中進行序列比對,發現該菌株與Stenotrophomonas maltophilia(登錄號MH396764.1)有較高相似性。用MEGA 7.0軟件按Neighbor-joining法構建系統發育樹,如圖5所示,菌株Z-4與Stenotrophomonas maltophilia WWi55不能分開。根據菌株Z-4的形態鑒定和16S rDNA序列分析,將菌株Z-4初步鑒定為嗜麥芽窄食單胞菌。
2.4 菌株生長曲線的繪制
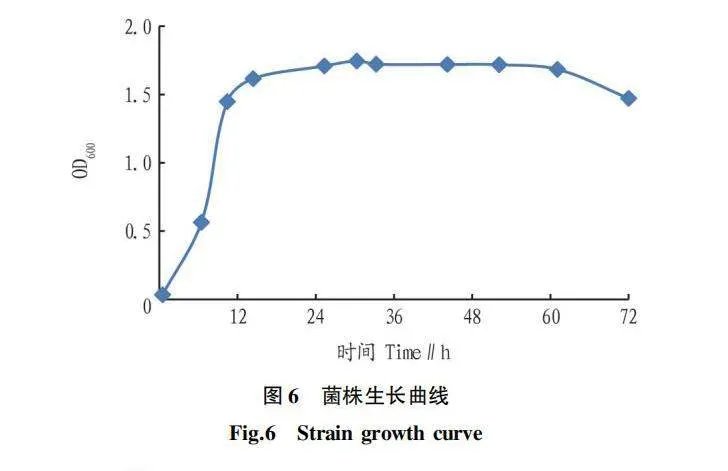
該試驗以所測的OD值為縱坐標、培養時間為橫坐標繪制菌株生長曲線,如圖6所示。菌株剛開始生長較緩慢,約有6 h的延遲期;6 h后,菌株生長快速,培養液的OD值呈對數上升,菌體處于對數生長期;到24 h 左右,培養液的OD值變化不大,菌株進入穩定生長的階段;在60 h以后,培養液中菌體的死亡量大于生長量,菌體生長處于衰亡期。由于在對數生長期的菌株生長活力大,生理和生化特征穩定,適應新環境的能力較強,進而提高了菌株的遺傳穩定性。所以,選取10 h為種子液的培養時間。
2.5 菌株產幾丁質酶條件的單因素試驗
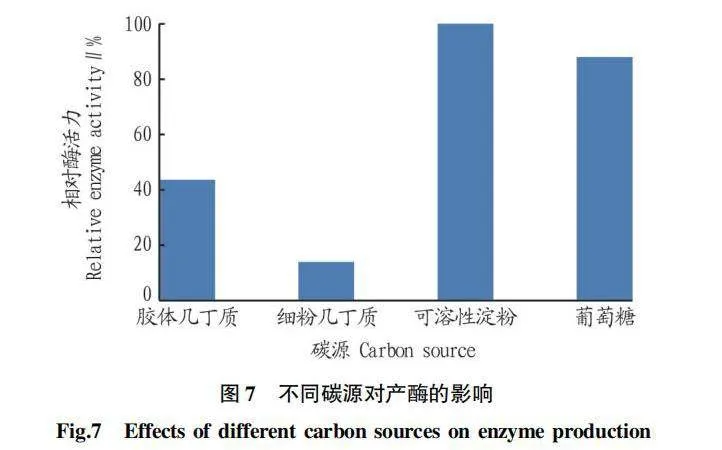
2.5.1 碳源對產酶的影響。
由圖7可知,菌株Z-4經膠體幾丁質誘導后,接種于不同碳源的發酵培養基中,所產酶量并不相同。其中,以可溶性淀粉作為碳源時,相對酶活力最高;以細粉幾丁質作為碳源時,相對酶活力最低。因此,選擇可溶性淀粉作為發酵培養基的碳源。
2.5.2 碳源濃度對產酶的影響。
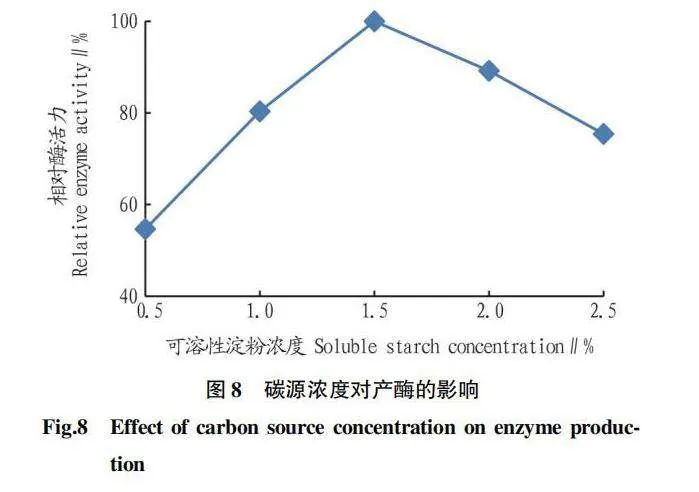
由圖8可知,當可溶性淀粉濃度過高或過低時,均不利于菌株產幾丁質酶;可溶性淀粉濃度為1.5%時,相對酶活力最高。
2.5.3 氮源對產酶的影響。
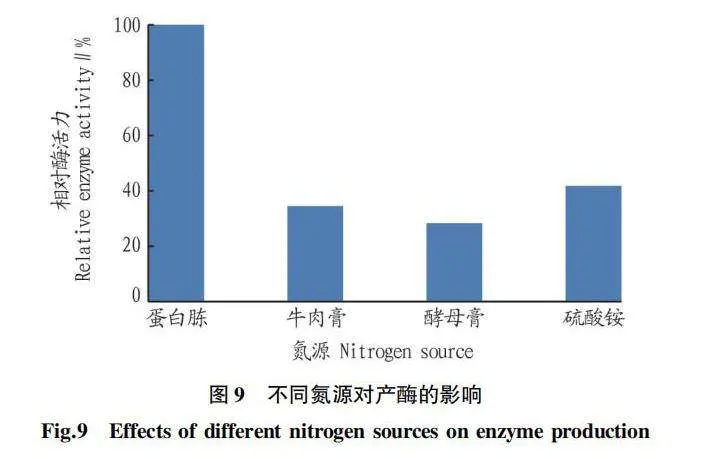
由圖9可知,采用不同氮源時,菌株Z-4的產酶能力各不相同。蛋白胨作為氮源時相對酶活力最高;而酵母膏作為氮源時相對酶活力最低。所以,選擇蛋白胨作為最佳氮源。
2.5.4 氮源濃度對產酶的影響。
由圖10可知,當蛋白胨濃度過高或過低時,均不利于菌株產幾丁質酶;蛋白胨濃度為2.5%時,相對酶活力最高。
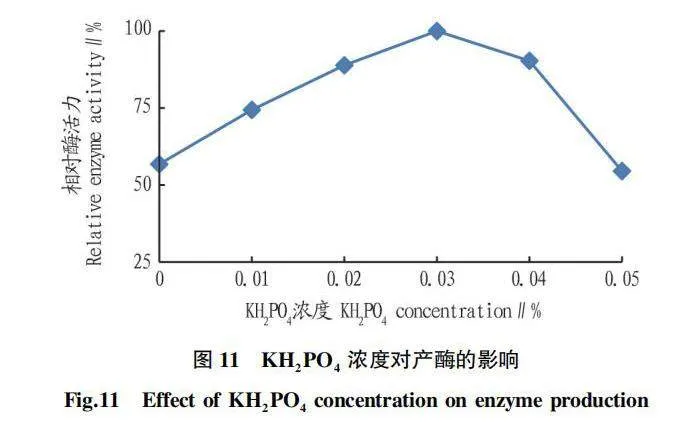
2.5.5 KHPO濃度對產酶的影響。
由圖11可知,KHPO對菌株產幾丁質酶起到促進作用,當KHPO濃度為0.03%時,相對酶活力最高,但當濃度達到0.05%時又起到抑制作用。
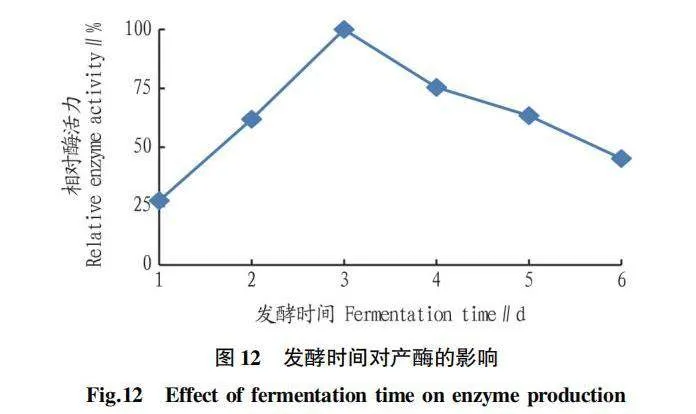
2.5.6 發酵時間對產酶的影響。由圖12可知,菌株Z-4剛開始培養時,相對酶活力增長緩慢;2 d后,相對酶活力增長迅速,到3 d時,達到最高值,之后隨著培養時間的延長,相對酶活力又緩慢下降。
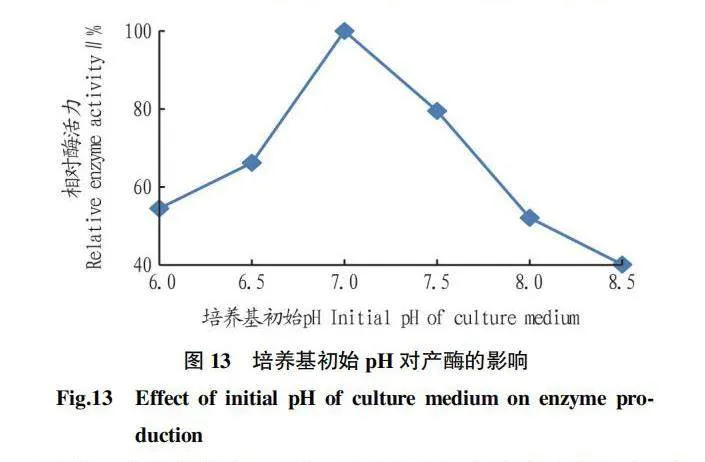
2.5.7 培養基初始pH對產酶的影響。由圖13可知,該菌株在中性或偏酸性條件下產幾丁質酶的能力比較強。當pH為7.0時,菌株的相對酶活力最高,當pH為8.5時,菌株的相對酶活力最低,幾丁質相對酶活力受培養基初始pH影響較大。
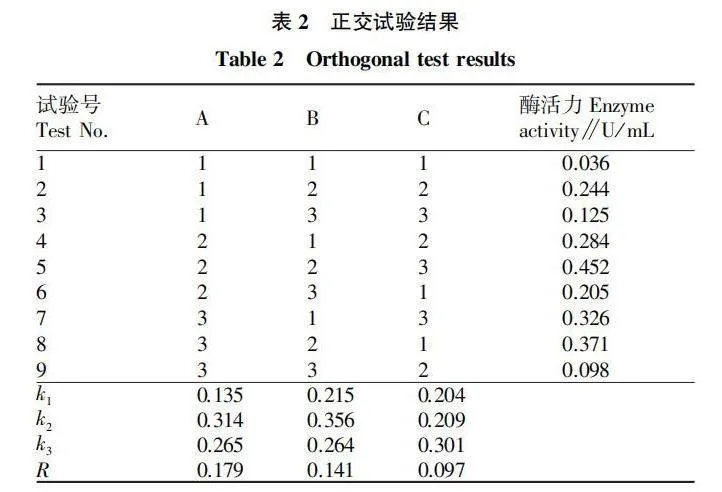
2.6 正交試驗
通過極差(R)分析(表2)可知,各因素對菌株Z-4產幾丁質酶影響的重要程度依次為A(可溶性淀粉添加量)>B(蛋白胨添加量)>C( KHPO添加量),即可溶性淀粉添加量對菌株產酶影響最大,蛋白胨添加量對其影響次之,影響最小的因素是KHPO添加量。
由正交試驗的結果還可以看出,細菌Z-4最適的產酶組合為ABC,即可溶性淀粉添加量1.5%、蛋白胨添加量2.5%、KHPO添加量0.04%,此時酶活力達到0.452 U/mL,比菌株Z-4 優化前的產酶活力(0.075 U/mL)提高了約5倍。
3 結論
從廣東湛江金沙灣的海水中篩選出6株產幾丁質酶的菌株,通過DNS法分別測定這6株菌的幾丁質酶活力,選取酶活力最高的菌株Z-4作為試驗菌株,經形態特征觀察及16S rDNA序列分析,初步確定該菌株為嗜麥芽窄食單胞菌。產酶條件優化結果顯示:蛋白胨添加量為2.5%,可溶性淀粉添加量為1.5%,KHPO添加量為0.04%,發酵培養基的初始pH為7.0,2%接種量,在30 ℃、160 r/min、培養72 h,菌株Z-4所產幾丁質酶活力最高,為0.452 U/mL,相比優化前提高了5倍。
參考文獻
[1] 王琳,陳雅如,程湄婕,等.微生物幾丁質酶研究進展及應用[J].中國生物工程雜志,2022,42(12):101-110.
[2] THAKUR D,CHAUHAN A,JHILTA P,et al.Microbial chitinases and their relevance in various industries[J].Folia microbiologica,2023,68(1):29-53.
[3] 周玉玲,蔣思婧,賀妮莎,等.微生物幾丁質酶研究進展及其在N-乙酰氨基葡萄糖制備中的應用[J].微生物學報,2021,61(8):2192-2204.
[4] 尹璐.幾丁質降解菌篩選及發酵產酶條件研究[D].大連:大連工業大學,2011:34-35.
[5] 劉力睿,潘杰,李猛.微生物幾丁質酶的研究進展、應用及展望[J].生物資源,2020,42(5):494-504.
[6] 張新月,張月琪,王鳳彪,等.海洋細菌來源幾丁質酶的研究進展[J].食品工業科技,2021,42(22):383-389.
[7] 馬欽元,申雁冰,丁盼盼.常壓室溫等離子誘變與微生物微滴培養選育幾丁質脫乙酰 基酶高產菌株[J].中國釀造,2020,39(8):170-174.
[8] 蔡亞萍,蘇建強,謝忠,等.南海海域幾丁質降解菌的篩選及其特性研究[J].廈門大學學報(自然科學版),2008,47(S2):259-263.
[9] 王曉輝,張慶芳,遲乃玉.低溫幾丁質酶生產菌的篩選鑒定與產酶條件研究[J].大連大學學報,2014,35(3):82-87.
[10] 孫軍德,喬雪,李海群,等.富產幾丁質酶的鏈霉菌菌株篩選及其產酶條件優化[J].沈陽農業大學學報,2015,46(3):309-315.
[11] 舒玉鳳,李嬋娟,張麗.產幾丁質酶細菌的篩選、鑒定及酶學性質研究[J].湖北農業科學,2016,55(15):3887-3890.
[12] 付星,閆巧娟,江正強,等.高產幾丁質酶巴倫葛茲類芽孢桿菌的篩選和發酵條件優化[J].微生物學通報,2015,42(4):625-633.
[13] 施騰鑫,劉嘉,賀淹才.黏質沙雷氏菌產幾丁質酶的發酵工藝優化[J].華僑大學學報(自然科學版),2010,31(6):667-670.
[14] 袁淑博,陳曉通,余夢林,等.MEW06產幾丁質酶響應面發酵條件優化研究[J].綠色科技,2018(12):156-160.
[15] 鄧紅梅,畢方鋮,葉炬斌,等.幾丁質酶高產菌的篩選及其產酶條件的優化研究[J].化學與生物工程,2010,27(5):62-65.
[16] 謝啟發,薛小旭,黃志發,等.海洋產幾丁質酶廈門加西利亞單胞菌Chi34的篩選、鑒定及酶學性質分析[J].食品工業科技,2023,44(12):116-123.
