頂壇花椒雄花花粉形態掃描電鏡觀測及生活力測定
2024-10-25 00:00:00羅輝曾欽朦盧中科侯娜婁麗王港
中國瓜菜 2024年10期




摘 要:以頂壇花椒花為試驗材料,使用Eclipse E200生物顯微鏡和EM-30 Plus型掃描電鏡對其花樣品進行形態學觀察和花粉活力分析,旨在為頂壇花椒雜交育種提供科學依據。結果表明,頂壇花椒出現雌雄同株的現象,花粉成熟后花藥呈鮮黃色,花粉粒為長球形,表面具細網狀紋飾。不同單株花粉粒直徑存在顯著差異,其中1號單株的花粉粒直徑最大(522.32 μm),3號單株的花粉粒直徑最小(279.50 μm)。花粉的極軸長、赤道軸長、花粉大小及網孔直徑的平均長度分別為26.76、16.39、442.56和0.81 μm。花粉活力測試結果表明,1號和2號單株的花粉活力較高,亞歷山大染色率分別為44.37%和48.57%,但植株長勢較弱;5號單株的花粉活力較弱,亞歷山大染色率僅有6.67%,但整個植株長勢正常,無衰弱跡象。研究結果可為頂壇花椒繁殖策略的制定提供生物學依據。
關鍵詞:頂壇花椒;形態構造;花粉粒大小;花粉活力
中圖分類號:S573+.9 文獻標志碼:A 文章編號:1673-2871(2024)10-0118-06
收稿日期:2023-05-10;修回日期:2024-08-21
基金項目:貴州省科技廳中藥現代化項目(黔科合ZY字〔2012〕3002號);貴州省林業廳項目(黔林科合2018〔01〕);貴州省科技廳成果轉化項目(黔科合成果〔2019〕4233號);貴州省科技廳優秀青年科技人才項目(黔科合平臺人才[20195643]);貴州省科技廳科技支撐項目(黔科合支撐〔2021〕222號,黔科合支撐〔2022〕一般118號)
作者簡介:羅 輝,男,高級工程師,主要從事經濟林栽培研究。E-mail:785486162@qq.com
通信作者:王 港,男,副研究員,主要從事林木遺傳育種研究。E-mail:417328697@qq.com
Scanning electron microscopic observation and viability test of pollen grains of Zanthoxylum planispinum var. dintanensis
LUO Hui1, ZENG Qinmeng2, LU Zhongke2, HOU Na2, LOU Li2, WANG Gang2
(1. Guizhou Extension Station of Forestry Science and Technology, Guiyang 550001, Guizhou, China; 2. Guizhou Academy of Forestry Sciences, Guiyang 550005, Guizhou, China)
Abstract: The morphological observation and, pollen vitality analysis of the Zanthoxylum planispinum var. dintanensis flower were conducted using an Eclipse E200 biological microscope and an EM-30 Plus scanning electron microscope. This study aimed to provide a scientific basis for the hybrid breeding of Zanthoxylum bungeanum. The results showed that there was a phenomenon of monoecious growth in the Zanthoxylum planispinum var. dintanensis. Upon maturation, the anthers turned bright yellow, and the pollen grains were elongated spheres with fine mesh patterns on the surface. Significant variations were observed among different individual plants, specifically, the pollen grain diameter of No. 1 was the largest(522.32 μm), while that of No. 3 was the smallest(279.50 μm). The average length of the polar axis, equatorial axis, pollen size, and mesh diameter of the pollen were 26.76, 16.39, 442.56, and 0.81 μm, respectively. Pollen vitality tests revealed that No. 1 and No. 2 exhibited high vitality, with Alexander staining rate of 44.37% and 48.57%, respectively, although their associated plant growth was relatively weak. Conversely, the pollen vitality of Individual No. 5 was low, with an Alexander staining rate of only 6.67%, while the overall growth of the plant appeared normal with no signs of weakening. These research findings provide a biological basis for understanding the reproductive strategy of the Zanthoxylum planispinum var. dintanensis.
Key words: Zanthoxylum planispinum var. dintanensis; Morphological structure; Pollen size; Pollen vitality
頂壇花椒(Zanthoxylum planispinum var. dintanensis)是竹葉椒(Zanthoxylum planispinum)的一個變種[1],為蕓香科花椒屬植物,單性花,花序上所有小花均為雌花,無雄花,大多為純雌株。頂壇花椒在貴州當地廣為栽培,已成為重要的經濟林木,對當地的生態環境修復發揮著重要作用[2]。近年來,許多學者對頂壇花椒開展了深入研究,主要集中在營養元素[3]、生理[4]、土壤養分[5]等領域,但對頂壇花椒花粉活力方面的研究較少。花粉是植物的特有器官,是植物種質保存、雜交育種及交換的重要材料[6]。不同植物的花粉經過演化和發展形成了獨特的形態特征,即便是在同一種內的花粉,其生活力和形態也具有多樣性[7]。花粉的形態特征主要受內環境的影響,通常由基因控制,具有較強的遺傳穩定性及保守性[8],可為系統發育、識別和植物種類的鑒定提供科學依據,并在分類學及遺傳學中廣泛應用[9]。
無融合生殖是一種特殊的無性繁殖方式,在不經過雌雄配子融合的情況下產生種子,并能培育為成熟個體。這種生殖方式能夠使植物的基因型保持不變,從而固定雜種優勢,對作物育種具有極其重要的意義。無融合生殖在植物中的表現形式多樣,包括營養的無融合生殖、無融合結子等類型,其遺傳機制復雜,受多種途徑調控[10]。花椒屬植物普遍具有一個重要的生物學特性——無融合生殖,即雌性植株不需要經過受精,直接由珠心細胞發育成胚,進而繁殖后代[11]。劉映紅等[12]研究表明,野花椒有正常的花粉,且花粉具有活力;葉萌等[13]對青花椒進行6年的觀察,發現四川金陽地區的青花椒均為純雌株,無雄花,而四川峨眉山市、洪雅縣、雅安市雨城區有極少的藤椒存在雄花;李立新等[14]對花椒進行遺傳多樣性分析,發現由于花椒是無融合生殖方式,因此種群內變異較小。Fei等[15]通過細胞學觀察、分子標記以及倍性鑒定等手段,進一步證明了花椒具有無融合生殖特性,并篩選出參與無融合胚胎發生的候選基因。此外,Wang等[11]對花椒無融合生殖的分子機制提出了新的見解,并基于基因組、轉錄組和代謝組技術對花椒屬植物的兩種生物堿的生物合成、無融合生殖及其抗旱特性進行了深入研究。
雌雄同株是指一個植物體上既有雌花又有雄花的現象,雌雄同株的植物可以自花授粉,也可以異花授粉,這種特性在植物繁殖策略中具有一定的優勢[16]。筆者在進行頂壇花椒種質資源調查和品種選育的過程中,連續6年進行觀察,發現頂壇花椒大多為純雌株,部分頂壇花椒園中存在5%~10%著生雄花的植株,且出現雌雄同株的現象,同時花粉具有活力,這與上述學者的研究結果有所不同。鑒于此,筆者連續2年在頂壇花椒花期采集樣品,采用生物學顯微鏡、掃描電鏡及TTC染色法對具有代表性的頂壇花椒單株雄花形態構造、花粉形態指標及花粉活力等進行研究,以期為頂壇花椒的發育生物學理論及雜交育種提供科學依據。
1 材料與方法
1.1 試驗地概況
試驗地位于貴州省貞豐縣北盤江鎮,地處珠江水系西江上源北盤江畔,屬喀斯特地貌地區,海拔550~1470 m,該地區屬北亞熱帶高原季風濕潤氣候,氣候溫和,年均溫16.4 ℃,全年無霜期335 d,雨量充沛,年均降水量1300 mm,雨熱基本同季,年均日照時數1549 h,年均相對濕度81%,土壤以石灰土為主。
1.2 樣品采集及保存
2021年3月至2022年3月,選擇具有代表性的5株雌雄同株(編號1~5號)頂壇花椒,從樹冠的不同方向采集正處于頂壇花椒盛花期的成熟花朵。將樣品放置于恒溫箱中干燥3 h,散粉后取下花粉,放置于鋪有濾紙的培養皿中,于4 ℃冰箱保存備用。
試驗地頂壇花椒不同單株見圖1,其中1~4號(圖1-A~D)單株的樹齡為12年生,5號(圖1-E)的樹齡為2年生,在連續兩年的觀察中發現都有少數的雄花出現,且均為雌雄同株。1號頂壇花椒的植株長勢較弱,表現出衰弱跡象,2號頂壇花椒一半有雄花,一半無雄花,同時發現開雄花的枝條長勢比無雄花的枝條長勢弱,3號和4號也出現同樣的情況,但5號頂壇花椒幼株,開完雄花后植株長勢正常。
1.3 研究方法
1.3.1 顯微鏡及掃描電鏡觀察 采用Eclipse E200生物學顯微鏡對花朵進行形態觀察。將保存備用的樣品用軟毛刷輕微蘸取微量花粉,輕掃于電鏡的固定液中,搖勻,在室溫下固定2 h,再放置于4 ℃冰箱保存。固定好之后,使用0.1 mol·L-1的PB(磷酸)緩沖液清洗3次,每次15 min;依次經過不同濃度(φ,后同)的酒精(30%、50%、70%、80%、90%、95%、100%)進行脫水,再放置于干燥器皿內干燥,將干燥后的樣品在ETD-900M磁控離子濺射儀中噴金30 s左右,置于EM-30 Plus超高分辨率臺式掃描電鏡下,選擇有代表性的視野觀察花粉的形態特征,隨機選取10粒花粉進行測量。
1.3.2 花粉活力檢測 采用亞歷山大染色法進行花粉活力檢測[17]。使用毛筆將花粉粒輕輕刷到載玻片上,并用滴管滴加亞歷山大染料溶液染色5~10 h,然后在顯微鏡下觀察、統計,原生質呈現紅色且花粉壁呈現綠色的花粉具有活力,均呈現綠色的則為無活力或發育不良花粉。觀察2~3張片子,每片取5個視野,統計花粉活力。花粉活力/%=紅色花粉粒數/花粉粒總數×100。
1.4 數據分析
試驗數據采用Excel 2007進行統計分析,用AutoCAD 2022測量花粉的形態指標,包括極軸長(P)、赤道軸長(E)、極赤比(P/E)和網孔直徑,P×E為花粉的大小;參照《孢粉學概論》[6],0.88<P/E<1.14為圓球形,1.14≤P/E<1.33為近長球形,1.33≤P/E<2.00為長球形,P/E≥2.00為超長球形。利用Origin 2020b軟件進行形態指標的差異顯著性分析。
2 結果與分析
2.1 頂壇花椒花的形態構造及花粉粒大小
利用Eclipse E200生物顯微鏡對采集的樣品進行觀察,結果如圖2所示。雌花的花小,花被片一輪,平均直徑為3 mm,淺綠色(圖2-A);子房上位有明顯的花盤,花盤上著生2個離生心皮組成的雌蕊;子房為淺綠色,外側向外凸起,且下部內側相貼,呈花瓶狀;花柱為閉合型,平均長620 μm,淺黃綠色,外傾且柱頭二裂(圖2-C)。雄花的大小與雌花相似,花被片一輪,8~10枚,綠色,有退化的雌蕊著生于花被片的內側,6枚,花藥背側著生,為橢圓形,花粉成熟后的花藥呈鮮黃色,花粉為黃色(圖2-B、2-D~E)。
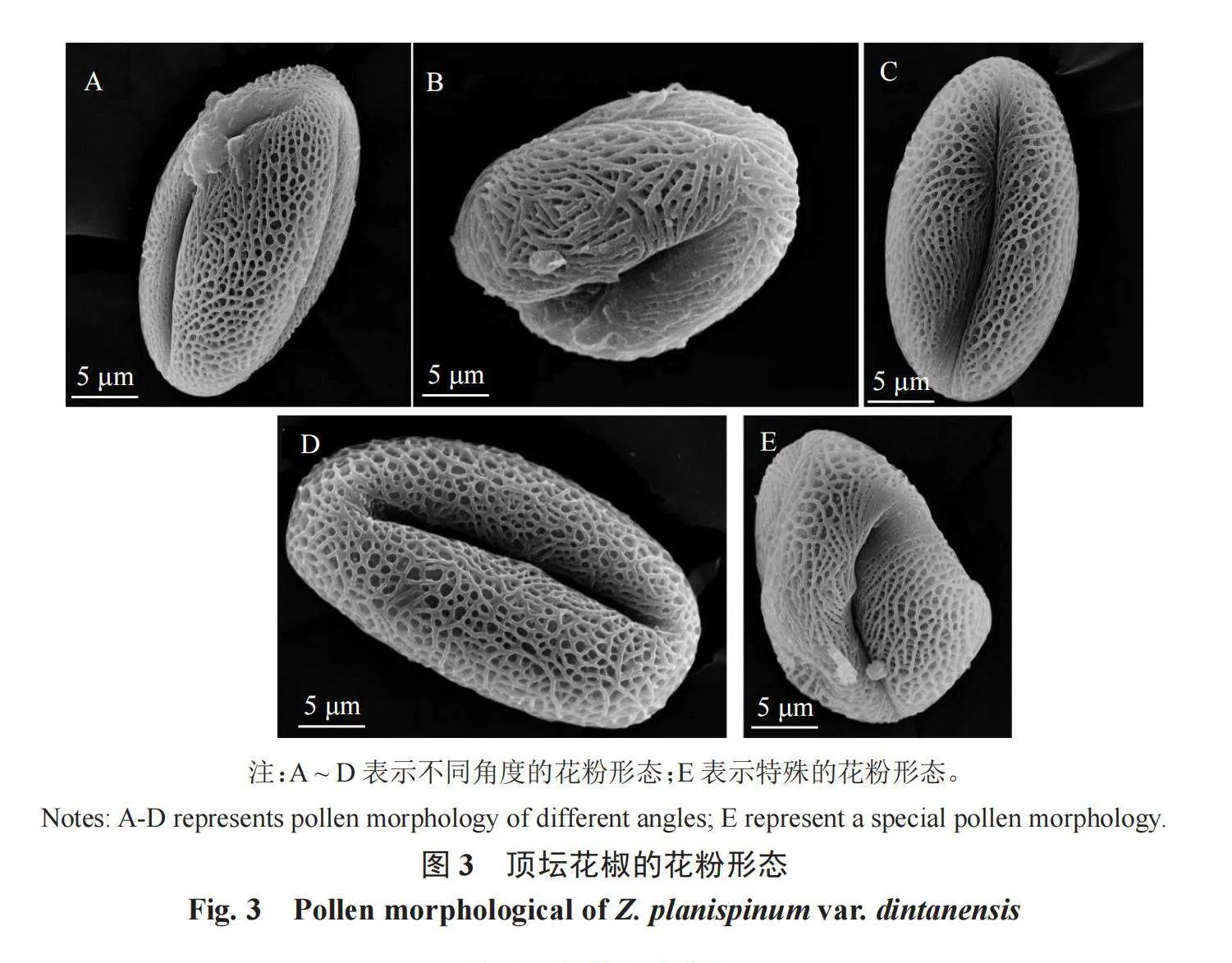
2.2 頂壇花椒花粉形態掃描電鏡特征及分析
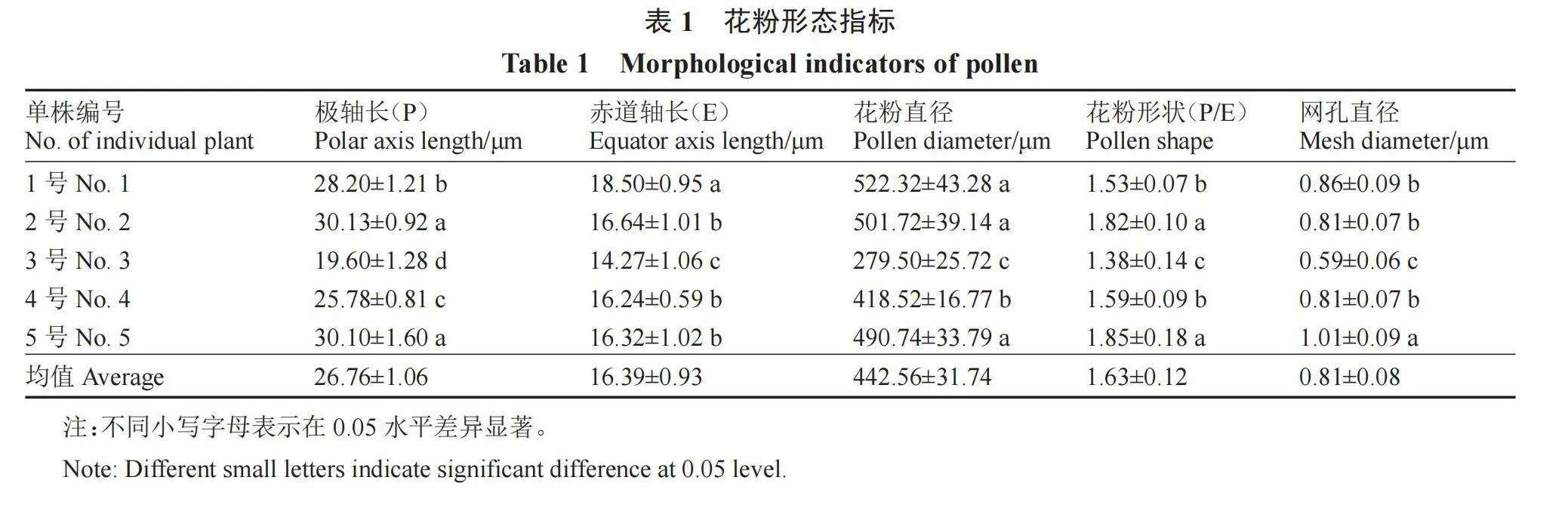
利用EM-30 Plus型掃描電鏡對頂壇花椒不同單株的花粉形態進行觀察,結果如圖3和表1所示。花粉為長球形,偶有特殊的花粉形態,如圖3-E,花粉的極面觀為3裂的橢圓形,具孔溝,細長,勾線形,表面具有細網狀紋飾,網孔呈不規則狀。2號和5號單株的極軸長差異較小,顯著高于其他單株,3號單株的極軸長最小,為19.60 μm;1號單株的赤道軸長為18.50 μm,顯著高于其他單株。3號單株的花粉直徑為279.50 μm,顯著小于其他單株;1號單株的花粉直徑最大,為522.32 μm。頂壇花椒5個單株花粉形狀的P/E 值存在顯著差異,但1.33<P/E<2.00,均為長球形。5號單株的網孔直徑最大,為1.01 μm,顯著大于其他單株;3號單株的網孔直徑最小,為0.59 μm,顯著小于其他單株。
2.3 頂壇花椒花粉活力檢測
花粉活力是影響經濟樹種初始坐果率及產量的重要因素,頂壇花椒5個單株的花粉活力差異顯著。從圖4和圖5可以看出,1號和2號頂壇花椒花粉活力經亞歷山大溶液染色后呈紅色的較多,分別為44.37%和48.57%,顯著高于其他單株;5號頂壇花椒花粉亞歷山大溶液染色后呈綠色的較多,花粉活力僅為6.67%。以上結果說明1號和2號頂壇花椒花粉活力較強,發育良好,而5號頂壇花椒多為發育不良或活力較弱的花粉。
3 討論與結論
在《四川植物志》[18]、《貴州植物志》[19]和《浙江植物志》[20]等各地植物志中均寫到竹葉花椒為單性花,在《福建植物志》[21]、《云南植物志》[22]和《中國植物志》[23]中描述了竹葉花椒的雌花和雄花。頂壇花椒是竹葉花椒的變種,均為無融合生殖;劉映紅等[12]發現,北京栽培的花椒未找到雄株,也未見雄花,而野花椒為雌雄異株;而葉萌等[13]發現,栽培的竹葉花椒兩個品種(藤椒和金陽青花椒)為雌花,具有無融合生殖特點,峨眉山市的藤椒園有雄花出現,但未做深入研究。筆者連續6年觀察到試驗地的頂壇花椒開出大量的“黃花”,且有雌雄同株的現象,這與劉映紅等[12]的研究結果不一致。通過對頂壇花椒的實地及顯微鏡觀察,并對花粉活力檢測,發現1號和2號頂壇花椒單株的花粉活力最強,但其植株長勢較弱,特別是1號頂壇花椒單株整株長勢弱,有枯死跡象,而2號頂壇花椒單株左側的枝條開雄花,右側的枝條無雄花,發現無雄花的枝條長勢比開雄花的枝條強,有可能是因為開雄花的枝條養分消耗較多,而無法正常供應整個枝條正常生長,導致植株長勢衰弱,3號和4號頂壇花椒也表現出同樣的現象。5號頂壇花椒整株開雄花后,整個植株長勢正常,無衰弱跡象,出現這種現象可能是因為5號頂壇花椒單株是幼株,其生長能力旺盛,養分充足,且開出的雄花花粉活力差,因此開雄花對5號幼株的長勢影響較小。
筆者對不同頂壇花椒單株的花粉形態指標進行測定,極軸長為19.60~30.13 μm,赤道軸長為14.27~18.50 μm,花粉直徑為279.50~522.32 μm,這與劉炳倫[24]研究的花椒屬的形態特征相似。開雄花的植株會出現植株生長衰弱的現象,但幼株并沒出現這一現象。目前尚不清楚其發生原因,后續將進一步觀察頂壇花椒產生雄花的現象是否與環境有關,對頂壇花椒雄花現象的深入研究對完善花椒的生殖發育理論和生產應用具有重要價值。
綜上所述,筆者發現頂壇花椒出現雌雄同株現象,通過掃描電鏡觀察得出頂壇花椒花粉為長球形,偶有特殊形態,花粉的極面觀為3裂的橢圓形,表面具有細網狀紋飾,網孔呈不規則狀。不同頂壇花椒單株的花粉活力存在差異,1號和2號單株的花粉活力較高,但植株長勢較弱;5號單株的花粉活力較弱,但植株正常生長。
參考文獻
[1] APPELHANS M S,REICHELT N,GROPPO M,et al.Phylogeny and biogeography of the pantropical genus Zanthoxylum and its closest relatives in the proto-Rutaceae group (Rutaceae)[J].Molecular Phylogenetics and Evolution,2018,126:31-44.
[2] 李紅,喻陽華,龍健,等.頂壇花椒葉片功能性狀對早衰的響應[J].生態學雜志,2021,40(6):1695-1704.
[3] 敖厚豫,李欣,余天華,等.基于麻味成分的頂壇花椒HPLC指紋圖譜研究[J].中國調味品,2020,45(5):19-26.
[4] 李紅,喻陽華.干熱河谷石漠化區頂壇花椒葉片功能性狀的海拔分異規律[J].廣西植物,2020,40(6):782-791.
[5] 喻陽華,鐘欣平,李紅.黔中石漠化區不同海拔頂壇花椒人工林生態化學計量特征[J].生態學報,2019,39(15):5536-5545.
[6] 王開發,王憲曾.孢粉學概論[M].北京:北京大學出版社,1983.
[7] 尹佳蕾,趙惠恩.花粉生活力影響因素及花粉貯藏概述[J].中國農學通報,2005,21(4):110-113.
[8] SALMAKI Y,JAMZAD Z,ZARRE S,et al.Pollen morphology of Stachys (Lamiaceae) in iran and its systematic implication[J].Flora,2008,203(8):627-639.
[9] 武海霞,劉麗婷,廖柏勇,等.20種桉樹及雜交種的花粉形態分析及分類學意義[J].中南林業科技大學學報,2012,32(3):29-36.
[10] 張燕,王春,王克劍.人工創制植物無融合生殖的研究進展[J].科學通報,2020,65(27):2999-3007.
[11] WANG M C,TONG S F,MA T,et al.Chromosome-level genome assembly of sichuan pepper provides insights into apomixis,drought tolerance,and alkaloid biosynthesis[J].Molecular Ecology Resources,2021,21(7):2533-2545.
[12] 劉映紅,王伏雄,錢南芬.花椒和野花椒的無融合生殖[J].遺傳學報,1987,14(2):107-113.
[13] 葉萌,楊德富,趙敏,等.青花椒的無融合生殖現象[C]//中國林學會.第三屆中國林業學術大會論文集,2013.
[14] 李立新,司守霞,魏安智,等.基于花椒轉錄組序列SSR分子標記開發及花椒種質鑒定[J].華北農學學報,2017,32(5):69-77.
[15] FEI X T,SHI Q Q,QI Y C,et al.ZbAGL11,a class D MADS-box transcription factor of Zanthoxylum bungeanum,is involved in sporophytic apomixis[J].Horticulture Research,2021,8(1):23.
[16] 吳淋慧,董家琦,鄭浩,等.利用獼猴桃雌雄花同株材料發掘性別決定相關基因[J].植物遺傳資源學報,2024,25(7),1153-1162.
[17] ALEXANDER M P.A versatile stain for pollen,fungi,yeast and bacteria[J].Stain Technology,1980,55(1):13-18.
[18] 四川植物志編輯委員會.四川植物志:第九卷[M].成都:四川人民出版社,1981.
[19] 《貴州植物志》編輯委員會.貴州植物志:第二卷[M].貴陽:貴州人民出版社,1986.
[20] 韋直,何業祺.浙江植物志:第三卷[M].杭州:浙江科學技術出版社,1993.
[21] 福建省科學技術委員會《福建植物志》編寫組.福建植物志:第二卷[M].福州:福建科學技術出版社,1985.
[22] 中國科學院昆明植物研究所.云南植物志:第六卷[M].北京:科學出版社,1995.
[23] 中國科學院中國植物志編輯委員會.中國植物志:第四十三卷第二分冊[M].北京:科學出版社,1997.
[24] 劉炳侖.中國蕓香科植物的花粉形態[J].植物研究,1987,7(3):11-37.
