杉木萌芽林結構多樣性研究
2024-11-03 00:00:00薩日娜廉琪胡艷波張弓喬盧彥磊孫洪剛姚甲寶趙中華
林業科學研究 2024年5期

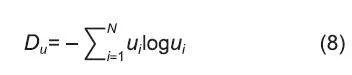





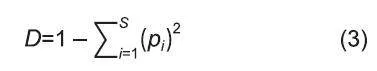



摘要:[目的]多角度分析杉木萌芽林的結構特征,為優化調整林分結構,精準提升杉木萌芽林林分質量提供依據。[方法]以亞熱帶林業實驗中心的山下林場杉木萌芽林為對象,采用3參數Weibull分布擬合直徑分布,用林分空間結構量化分析方法分析林分空間結構參數一元和二元分布特征、樹種優勢度及林分結構多樣性,并用遺傳絕對距離法進行差異顯著性檢驗。[結果]3個林分的樹種組成均為以杉木萌生株為主,混有少量闊葉樹,其Shannon-Winner指數在0.933~1.039之間,Simpson多樣性指數都小于0.5,說明樹種多樣性較低;林分直徑分布呈現左偏單峰山狀曲線,有向倒“J”型發展的趨勢;三板橋、七里坑上坡和下坡林分的平均角尺度分別為0.503、0.486和0與00,均屬于隨機分布的范疇,林分平均混交度分別為0.345、0.404和0.409,林分整體上處于弱度混交向中度混交過渡的狀態;3個林分的結構多樣性指數值較低,分別為1.423、1.349和1.649,角尺度-混交度、大小比數-角尺度二元分布表現出一定的規律性,但混交度-大小比數二元分布變化相對復雜,規律不明顯。[結論]杉木萌芽林樹種組成相對簡單,樹種多樣性和林分結構多樣性低,胸高直徑分布和林木分布格局初步具有天然林的特征,需要通過林分結構優化調整提升林分質量。
關鍵詞:杉木萌芽林;樹種多樣性;空間結構;結構多樣性;結構調整
中圖分類號:S750 文獻標識碼:A 文章編號:1001-1498(2024)05-0148-12
杉木(Cunninghamia lanceolata (Lamb.)Hook.)是我國南方低山丘陵區重要的優0jAf54bfiFjnF40IkcT85A==質鄉土用材樹種之一,在南方16個省(區)均有種植。第九次全國森林資源清查結果表明,杉木人工林無論是面積還是蓄積量均居第一,在我國森林蓄積量和木材生產中占有重要地位。杉木是具有極強萌芽能力的樹種,杉木萌芽更新投入少、技術簡單,因此,萌芽更新是杉木人工林的主要更新方式之一,也是一些杉木產區傳統的育杉方式。森林結構是各種自然生態過程在時間和空間上長期綜合作用的結果,是森林生長及生態過程的驅動因子,在很大程度上決定了森林穩定性、森林質量及碳儲量等。林分結構是研究森林結構的基本單元,即是森林經營與分析的最小單位,也是實施森林經營活動的具體對象,一直是森林經營研究的重點問題。眾多研究表明,林分結構調控是提升森林質量的有效手段,比如林分密度調控,已有大量的文獻研究表明,密度調控能夠影響各生長時期林分的樹高、胸徑、林分木材產量等因子,在很大程度上決定了林分的內部結構,從而決定了林分的產量結。林分結構包括非空間結構和空間結構,非空間結構包括樹種組成、樹高結構、直徑結構、林分密度和年齡結構等;林分空間結構從我國學者惠剛盈等提出了基于相鄰木關系的林分空間結構參數體系以來,由于該體系能夠量化和多層次分析林分空間結構,且能夠指導經營實踐而得到廣泛應用。同時近年來,學者們對于林分結構多樣性的研究也越來越多。但林分結構及結構多樣性評價調控應用于萌芽林的研究十分缺乏。
從上世紀80年代以來,我國學者針對杉木萌芽林開展了大量的研究,主要集中在采伐年齡、采伐季節、采伐工具等對杉木萌芽能力的影響,杉木萌芽林的經營技術措施、二代實生林與萌芽林的生長特性、萌芽幼齡林群落特征及杉木萌芽林改造針闊葉混交林等方面、杉木萌芽更新研究及杉木萌芽更新經營技術研究;關于杉木萌芽林的空間結構特征及經營對空間結構的影響研究極少。周紅敏等對對浙江龍泉萌生杉木林的空間結構進行分析,根據萌生杉木林的結構特征提出經營對策。張煜林等以不同密度杉木萌生林自然恢復初期林分為研究對象,分析了林分碳密度與樹種多樣性、結構多樣性及林分空間結構參數之間的關系。本研究以位于江西省新余市分宜縣的杉木萌芽林為研究對象,從林分直徑分布、樹種多樣性、林分空間結構參數一元分布、二元分布及林分結構多樣性等方面精細分析杉木萌芽林的結構特征,并在此基礎上,針對杉木萌芽林林分結構存在的主要問題提出經營建議,以期為杉木萌芽林結構優化調控,精準提升杉木萌芽林質量提供依據。
1 研究區概況及研究方法
1.1 研究區概況
江西省新余市分宜縣位于江西中部偏西,位于珠三角、長三角和閩東南三角交匯點,袁河中游(114°29'~114°51'E.27°33'~28°08'N之間),縣境南北長約65 000 m,東西寬約36 000 m,兼具我國南北方氣候特征,屬亞熱帶濕潤性氣候,年均勻氣溫17.2℃,年均勻降雨量1 600 mm,全年無霜期270 d。植被系江南山地丘陵常綠栲楠林,油茶林、松杉林區,常綠栲楠、松杉亞區。全縣森林覆蓋率65.76%,有林地面積57 135.2 ha。
研究樣地位于中國林業科學研究院亞熱帶林業實驗中心的山下林場七里坑72-4和三板橋22-7林班,林分年齡為20年,均為一代杉木林皆伐后萌芽形成的林分,其中,七里坑72-4林班在林緣和山脊部人工栽植了少量的樂昌含笑(Micheliachapensis Dandy)、檫木(Sassafras tzumu(Hemsl.) Hemsl),桉樹(Eucalyptus robustaSmith)等,三板橋22-7林班,在林緣和山脊部人工栽植了木荷(Schima superba Gardn&Champ.),在林中還有少量的馬尾松(Pinus massonianaLamb.)、楓香(Liquidambar formosanaHance)、樟樹(Cinnamomum camphora (L.)Presl)、梓樹(Catalpa ovata G.Don)、苦櫧(Castanopsis sclerophyHa (Lindl.) Schot)、刨花楠(Machilus pauhoi kaneh)等;補植樹種與杉木在伐樁上萌發的林木及天然更新的樹種形成了混交林。
1.2 樣地設置與調查
2021年4月和2022年7月,在山下林場三板橋22-7和七里坑72-4林班內,運用TOPCON全站儀(GTS-601)建立了面積分別為60 m×40 m和兩個40 m×60 m定位監測樣地,對樣地內胸高(h=1.3 m)直徑大于5 cm的林木進行定位掛牌,記錄樹種、胸徑、樹高、冠幅、更新狀況、林木健康等基本信息。
1.3 內業分析
1.3.1 胸高直徑分布
林分直徑分布特征是林分結構中最重要的指標之一,是測定、研究林分直徑、斷面積、材積等生長因子的依據,是許多森林經營技術的基礎。本研究起測胸徑為5 cm,徑階寬度為2 cm,采用3參數Weibull分布擬合杉木萌芽的胸高直徑分布,其概率密度函數為:
式中,a、b、c分別為位置參數、尺度參數和形狀參數,d為對應徑階直徑。參數a為林分直徑最小徑階的下限值;當c≤1時,f(x)隨x的增大而減小;1<c<3.6時分布形狀偏于左邊;c=3.6時,近似正態分布;c>3.6時,偏于右邊。
1.3.2 樹種多樣性
樹種多樣性選擇Shannon-Wiener指數和Simpson指數計算。
Shannon-Wiener指數:
式中:ρi為第/個樹種株數在林分樹木總株數中所占百分比,S為林分中樹種的數目;
Simpson多樣性指數:
式中:參數與上同。
1.3.3 林分空間結構
運用森林空間結構分析軟件Winkelmass分析林分空間結構參數一元分布,主要計算的參數有角尺度、混交度和主要樹種優勢度;同時,將角尺度(Wi)、混交度(Mi)和大小比數(Ui)兩兩結合,分析林分空間結構參數二元分布,再利用遺傳絕對距離法對3個林分二元分布進行差異性檢驗。在計算3個空間結構參數時為避免邊緣效應對林分結構的影響,本研究設置緩沖區域為各邊長兩側的5%。
樹種優勢度利用樹種空間優勢度來表達,其計算公式為:
式中:Dg表示樹種相對顯著度,U表示樹種大小比數均值,樹種優勢度在[0,1]之間,越大越好。
1.3.4 林分結構多樣性
林分結構多樣性能表達不同林分和林型的結構異質性,包括樹種多樣性、分布和大小分化。采用Zhao等提出的林分結構多樣性表達方法分析。其計算公式如下:
式中,SD是結構多樣性指數,S是林分中的樹種數量,Pi是林分中第j個樹種的比例;N是結構單元類型的數量,ui是第i個結構單元類型所占的比例。H是香農指數;DU是結構單元類型的多樣性;CV是樹木大小的變異系數。SD的值由林分中樹種的多樣性、結構單元類型的多樣性和單株大小的變化決定。
CV=σ/μ(7)
式(7)為林木大小的變異系數,式中σ為林分中林木胸徑大小的標準偏差,μ是林分中林木胸徑平均值。
式(8)為結構單元多樣性,式中Du是結構單元類型的多樣性,Ui是第i個結構單元類型在所有組合情況下的比例,N是結構單元類型的數量;結構單元類型是根據結構單元中混交度、大小比數、角尺度和樹種數量取值進行聯合,把取值不同的結構單元看作是不同類型;混交度、大小比數、角尺度3個結構參數5種不同取值可能會出現的組合有C15×C15×C15=125種情況;當考慮結構單元中的樹種時,可能出現的結構單元類型種類為(C11+C11+C2+C13+C14)×C15×C15=275種,因此,結構單元類型最多不超過275。當森林中只有1種結構單元類型(N=1)時,Du的最小值為零。結構單元類型越多,比例越均勻,Du的值就越大;當N=275時,Du的最大值為5.617。
1.3.5 數據處理
運用Winkelmass分析林分的水平結構,并在Excel表格中進行林分空間結構參數二元分布特征分析,運用遺傳絕對距離法進行顯著性檢驗。并用R語言對林分胸高直徑進行3參數Weibull函數分布擬合。本研究內所有圖均使用origin2021作圖。
2 結果分析
2.1 杉木萌芽林樹種組成及多樣性
由表1可以看出,三板橋、七里坑上坡和七里坑下坡樣地中樹種數分別為11種、9種和11種,Shannon-Winner指數在0.933~1.039之間,Simpson多樣性指數都小于0.5,說明3個林分樹種多樣性均較低。3個林分中杉木的相對多度均達到了70%以上;其中,三板橋樣地中,木荷和馬尾松相對多度分別為13.2%和7.7%,其余8個樹種相對多度不到10%;七里坑上坡樣地中含笑的相對多度較高,達到了14%,下坡樣地含笑的相對多度則不到10%,其余樹種總占比不足10%。整體而言,3個林分中杉木萌芽林木在數量上占絕對優勢,其它混生的樹種主要集中在小班林緣和山脊部,因而導致樹種組成多但樹種多樣性較低。
2.2 杉木萌芽林胸高直徑分布
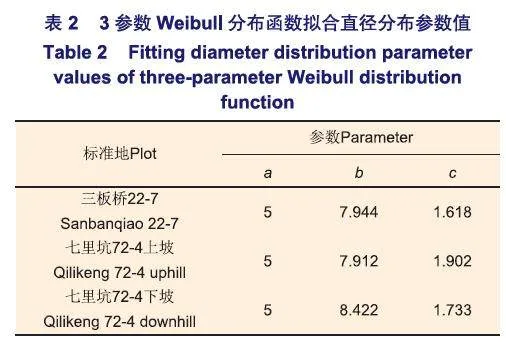
一般而言,沒有干擾的人工林胸高直徑結構呈正態分布,即中等林木占大多數,而小徑級及大徑級林木較少,而天然林呈倒“J”型分布,小徑級木占大多數,隨著徑階的增加,林木個體逐漸減少。三板橋22-7林班杉木萌芽林林木胸高直徑分布呈單峰分布,胸徑為8~14 cm的林木個體較多;七里坑72-4林班上坡林分呈多峰分布,林分中胸徑為10 cm和14 cm徑階的林木較多;七里坑72-4林班下坡林分整體上呈單峰分布,胸徑為10~14 cm的林木較多(圖1)。從3個林分的胸高直徑利用3參數Weibull分布函數擬合結果的形狀參數值(c)可以看出,三板橋22-7林班和七里坑72-4林班下坡林分的林木胸高直徑呈明顯的左偏單峰山狀分布(表1),72-4林班上坡林分的胸徑分布的柱狀圖顯示為多峰形狀山狀分布,但從擬合參數可以看出(表2),胸高直徑分布也趨于左偏單峰山狀(1<c=1.902<3.6)。以上分析表明,3塊杉木萌芽林胸高直徑集中分布在8~14 cm之間,呈現出明顯左偏單峰分布的特征,隨著徑階的增加,林木株數減少,已經具有了一些天然林徑級結構分布的特征。這一方面可能是因為伐樁上萌生的杉木由于萌發時間不同而產生分化,另一方面可能是由于在林緣和山脊部人工栽植了闊葉樹種,林下部分闊葉樹種天然更新,從而對林分胸高直徑分布產生了影響。
2.3 杉木萌芽林空間結構特征
2.3.1 林木分布格局及樹種隔離程度
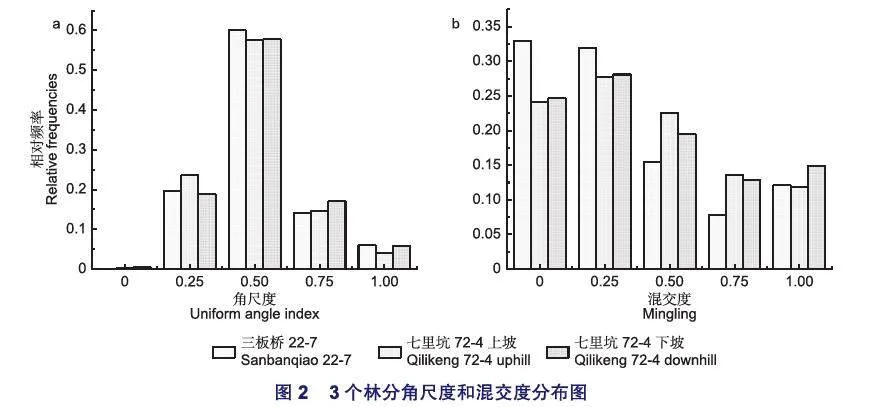
由圖2a可知,七里坑2塊樣地中角尺度值為0.5的林木比例大致相同,約占55%,三板橋樣地角尺度為0.5的比例略高,達到了60%;3塊樣地中,角尺度小于0.5和大于0.5的分布頻率大致相同,但3塊樣地中處于十分均勻(W=0)的比例極少,低于2%,而處于聚集分布(W=1)的比例較低,約為5%左右。三板橋、七里坑上坡以及下坡林分平均角尺度值分別為0.503、0.486和0.500,表明樣地中的林木的分布格局整體上為隨機分布的狀態。從圖2b可知,三板橋樣地中零度混交(M=0)和弱度混交(M=0.25)的林木比例較高,均達到了30%以上,而處于中度混交、強度混交和極強度混交林木的比例均小于15%,表明林分中的林木大多數為同種樹聚集;七里坑2塊樣地的混交度為零的林木比例較三板橋樣地的比例低,而處于中度混交、強度混交和極強度混交較三板橋樣地有所上升,表明七里坑2塊樣地的樹種隔離程度較三板橋樣地高。三板橋、七里坑上坡和下坡3塊樣地林分平均混交度值分別為0.345、0.404和0.409,林分整體上處于弱度混交向中度混交過渡的狀態。
2.3.2 樹種優勢度
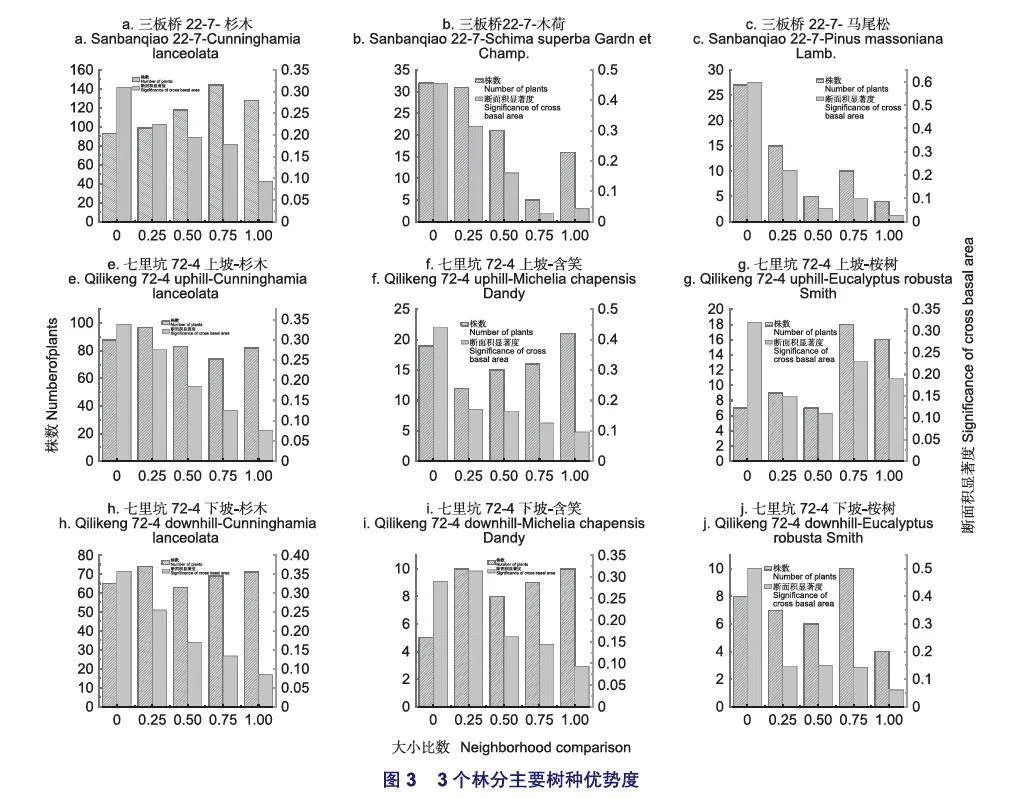
將樹種的大小比數均值與相對顯著度相結合,可以表達出樹種在林分中的優勢。對3個林分中主要樹種的優勢度進行了分析,結果見表3和圖3。3個林分的主要樹種中均為杉木的樹種優勢度最高,這主要是由于杉木萌生株數量較多,相對顯著度較高而造成的(表3)。結合圖3a-c可以看出,三板橋林分中杉木株數為582株而木荷株數僅有105株,但樹種優勢度僅相差0.134,說明該樣地中杉木大多為劣勢木到中庸木的比例較高,而木荷及馬尾松在林分中大多數為優勢木、亞優勢木,也就是說,以杉木為參照樹及其4株相鄰構成的結構單元中,大小比數為1、0.75和0.5所占的比例較高,而以木荷及馬尾松為參照樹及其4株相鄰木構成的結構單元中,大小比數為0、0.25所占的比例較高。與三板橋林分不同,七里坑上坡和下坡林分雖然杉木優勢度也是最高的,但杉木在結構單元中,大小比數取值較為相對平均,各個等級的比例相差不大,而其它2個主要樹種的大小比數取值則變化較大,而且大多數處于劣勢(Ui=1或Ui=0.75)和中庸狀態(Ui=0.5),因此,它們的優勢度較杉木相差較大(圖3e~j)。以上分析表明,3個林分中杉木萌生株由于數量和相對顯著度占絕對優勢,導致其優勢度總體較高,但其它主要樹種的競爭狀態并不相同,三板橋林分中的木荷與其相鄰木相比處于優勢狀態,而七里坑林發分中的含笑和桉樹大多處于劣勢狀態。
2.3.3 角尺度-大小比數二元分布
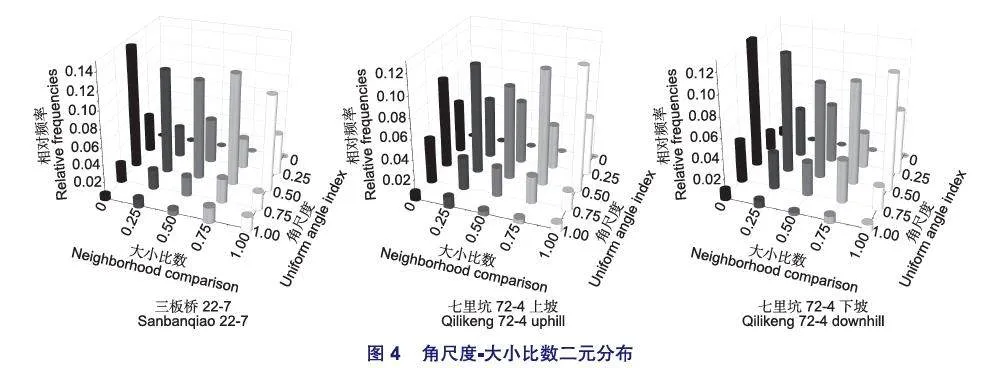
由圖4可以看出,3個林分中結構單元角尺度值為0.5時比例最高,其次為角尺度值為0.25和0.75的結構單元,且不同大小比數等級均有分布;其中三板橋林分同一大小比數等級56.7%以上林木處于隨機分布的結構單元中;林木處于隨機分布且絕對優勢(Wi=0.5,Ui=0)的林木比例最大,達到14.1%,其它大小等級且隨機分布的林木也在10%以上,該林分中沒有結構單元角尺度值為0的林木。在七里坑上坡林分中,同一大小比數等級45.9%以上林木處于隨機分布的結構單元中,林木處于隨機分布且絕對劣勢(Wi=0.5,Ui=1)的林木比例最大,達到12.2%。在七里坑下坡林分中,同一大小比數等級50%以上林木處于隨機分布的結構單元中,林木處于絕對優勢且為隨機分布(Ui=0,Wi=0.5)的林木比例最大,達到12.7%。綜上所述3個林分處于同一大小比數時隨機分布的林木占絕大數,同時絕對優勢林木處于隨機分布的最多,說明林木在隨機分布時長勢較好。3個林分的角尺度-大小比數二元分布遺傳絕對距離檢驗差異不顯著(dab=0.121<da=1.027、dbc=0.091 <da=0.976、dac=0.113<da=l .027)。
2.3.4 角尺度-混交度二元分布
從圖5可以看出,3個林分中林木處于零度和弱度混交的比例較高,合計均超過了50%,且在這些林木中,處于隨機分布的林木比例較高,其次為均勻分布的林木。其中,三板橋林分中,隨機分布且零度和弱度混交的林木占比為39.4%,均勻分布的林木為13.4%;七里坑上坡林分隨機分布且零度和弱度混交的林木占比為25.8%,均勻分布的林木為14.6%;七里坑下坡林分隨機分布且零度和弱度混交的林木占比為30.4%,均勻分布的林木為12.1%。3個林分中處于極強度和強度混交的林木較少,所占比例均沒有超過28%。3個林分的角尺度-混交度二元分布遺傳絕對距離檢驗差異不顯著(dab=0.201 <da=1.1 30、dbc=0.116<da=1.010、dac=0.157<da=1.130)。
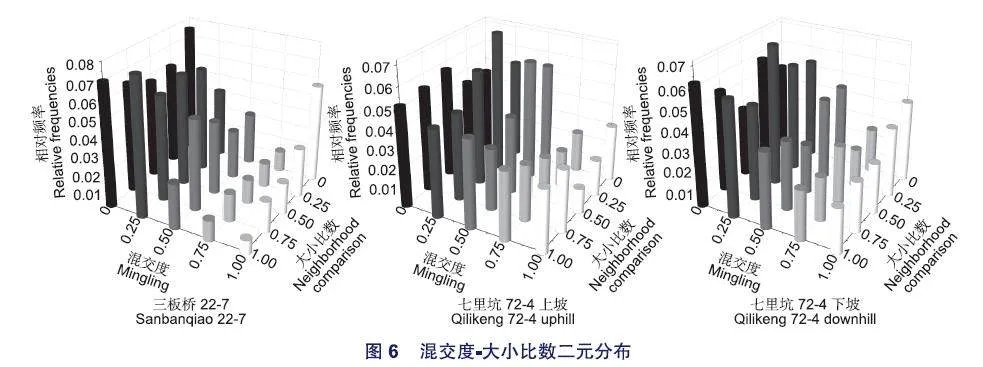
2.3.5 混交度-大小比數二元分布
由圖6可以看出,3個林分混交度-大小比數二元分布比較復雜,盡管可以明顯反映出林分中林木大多數處于中度混交以下,但對于同一大小比數等級林木的混交程度或同一混交程度不同大小比數等級林木的比例變化相對復雜,規律不明顯,例如在三板橋林分中,對于零度混交等級(Mi=0)而言,大小比數從優勢到劣勢的林木比例呈現先減少后增加的趨勢;而對于弱度混交等級(Mi=0.25)而言,大小比數從優勢到劣勢的林木比例呈現雙峰狀。對于同一大小比數等級而言,雖然3個林分總體上為隨著混交程度的增加,林木比例減少的規律,但也出現了一些例外,例如三板橋林分中的大小比數為優勢等級(Ui=0)的林木,呈現出隨著混交度的增加,林木比例先減少后增加,而且零度混交且絕對優勢(Mi=0,Ui=0)和強度混交且處于絕對優勢(Mi=1,Ui=0)的林木比例分別達7.4%和5.6%。這可能主要是由于萌生杉木和人工補植及天然更新的闊葉樹種的年齡、生長特性不同而導致,此外,3個林分的混交度—大小比數二元分布遺傳絕對距離檢驗差異也不顯著(dab-0.1 93<da=597、dbc=0.116<da=0.367、dac=0.1 50<da=0.597)。
2.3.6 林分結構多樣性
Zhao等提出的基于相鄰木關系的林分結構多樣性指數同時考慮了樹種多樣性、林木大小、分布格局、混交等因素,可表達出樹種越豐富、混交程度越高,林木大小分化、分布格局變異越大,林分結構多樣性越高的普遍規律。林分結構多樣性結果表明(表4),3個林分的結構單元類型數均在90個左右,遠低于3個結構參數組合類型的最大值275;3個林分的林分結構多樣性都較低,分別為1.423、1.349和1.649。結合林分的樹種組成和多樣性,造成林分結構多樣性低的主要原因是林分中杉木萌芽株占優勢,大多數林木與同種相鄰,由此造成結構單元的類型多樣性較低,此外,樹種多樣性和胸高直徑大小變異是造成3個林分林分結構多樣性存在一定差異的主要原因。
3 討論
林分結構規律研究是林分結構調控與優化的基礎,制定結構調控措施需遵循林分結構規律。杉木萌芽林是在杉木純林皆伐后自然萌發的基礎上,在林緣與山脊上人工補植闊葉樹后形成的林分。經過20多年的生長,3個林分直徑結構、分布格局已經初步具有天然林的特征、林分的樹種多樣性也有所提高,說明補植鄉土闊葉樹種確實可以人工促進樹種多樣性甚至在很大程度上可以改變林分直徑分布的規律。但整體上樹種組成仍然是以杉木萌生株占絕對優勢,樹種分配的均勻性低。這與其發展階段有密不可分的關系。皆伐自然萌生后同根的較多,補植樹種集中在林緣部,林分內闊葉樹種極少導致杉木仍占比例最高其它樹種比例極少且分配不均勻。
利用空間結構參數能夠準確描述林木空間結構的結構狀態。本研究通過分析林分空間結構一元二元分布發現3個林分存在的問題基本相同,整體林分呈弱度混交向中度混交過度的狀態,林分分布格局為隨機分布;同一混交度及大小比數對應的角尺度先增加再減小,在角尺度處于隨機分布時取得最大值。而同一分布格局的周圍大多是零度混交或弱度混交。上述結構特征反映杉木萌芽林的林分空間異質性較差,林內杉木喬木層同齡同種之間資源競爭激烈并且多株同根需要的資源更多,林下養分、水分、土壤肥力及生存空間不足,限制了林下植被繁殖、更新發育,不利于林下其它樹種的群落構建,這可能是導致3個林分混交度低的主要原因。這與王奕茹等對于杉木—閩楠混交林二元分布結構的研究結論和張崗崗對華北落葉松人工林空間結構的研究結論相似。
本研究采用了Zhao等提出的基于相鄰木關系的林分結構多樣性指數,其同時考慮了樹種多樣性、林木大小、分布格局、混交等因素。研究結果表明3個林分結構多樣性低。4個參數任一種變化都能影響結構多樣性。從上述林分結構得出樹種多樣性、樹種分配的均勻度和混交度較低,因此這也是導致結構多樣性低的主要原因。同時樹種分配的均勻性、樹種多樣性和混交度低是3個林分中共同存在并且主要解決的問題,建議調整和優化的方向是在保持林木整體分布格局隨機分布的前提下,采取去劣留優、見縫插針等方法,補植優良鄉土闊葉樹種,通過伐除周圍影響其生長的杉木萌生株,增加闊葉樹種的競爭優勢從而降低杉木萌生株的比例,增加林分的混交度,提高林分中闊葉樹種的優勢程度和林分結構多樣性。
4 結論
本研究探討杉木萌芽林林分結構多樣性特征情況,主要結論如下:3個杉木萌芽林均呈現樹種組成相對簡單,樹種多樣性和林分結構多樣性低,胸高直徑分布和林木分布格局初步具有天然林的特征,需要通過林分結構優化調整提升林分質量。因此本研究以期分析杉木萌芽林的林分結構和結構多樣性,發現其存在的問題,并針對性的提出優化建議,為杉木萌芽林結構優化調整提供依據。
(責任編輯:崔貝)
基金項目:中央級公益性科研院所基本科研業務費專項資金(CAFYBB2020ZB002-1);國家自然科學基金面上項目(32171780)
