茶樹接種AM真菌在干旱脅迫下的生理響應
2024-11-08 00:00:00魯薇鄔曉龍胡賢春郝勇劉春艷
茶葉科學 2024年5期



摘要:為探究干旱脅迫下叢枝菌根(Arbuscular mycorrhizal,AM)真菌對茶樹生長及生理的作用機理,以福鼎大白茶(Camellia sinensis cv. ‘Fuding Dabaicha’)茶樹實生苗為試驗材料,采用溫室盆栽法,在正常水分和干旱脅迫下分別接種或不接種AM真菌幼套近明球囊霉(Claroideoglomus etunicatum),探究AM真菌在不同干旱脅迫時長(4周、6周、8周)下對茶樹實生苗生長、光合、滲透調節(jié)及氣孔開度等的生理響應。結果顯示,在兩種不同的水分條件下,接種AM真菌處理均顯著促進了茶樹生長,增加了地上部和地下部生物量,改善了根系構型,與不接種AM真菌相比,茶樹根系總長度、二級側根和三級側根的數量、葉片滲透勢分別增加了20.00%~38.77%、48.90%~163.33%、60.00%~442.86%、29.77%~41.24%,降低了干旱脅迫下茶樹葉片氣孔密度和相對電導率,與不接種AM真菌相比分別降低了16.00%~42.37%、2.21%~6.67%,且在干旱脅迫下的效果更為顯著。干旱脅迫顯著抑制了AM真菌對茶樹根系的侵染和茶樹的生長,表現為抑制了茶樹根系構型的建立,降低了葉片葉綠素含量、最大光量子效應(QY_max)、葉片相對含水量(RWC)、氣孔開度及滲透勢等。接種AM真菌能顯著緩解這種抑制效應,改善茶樹對干旱脅迫的生理響應,從而促進茶樹生長。研究表明,AM真菌可通過優(yōu)化茶苗根系構型,提高茶苗葉片的保水和光合能力,調節(jié)氣孔及滲透平衡,從而促進植株對水分和養(yǎng)分的吸收,緩解干旱對茶樹的損傷,提高茶樹實生苗的抗旱能力,且隨著干旱時間的延長AM真菌的促進效果更為顯著。
關鍵詞:茶樹生長;生理特性;叢枝菌根真菌;干旱脅迫
中圖分類號:S571.1;S326 文獻標識碼:A 文章編號:1000-369X(2024)05-718-17
Physiological Response of Tea Plants Inoculated with Arbuscular Mycorrhizal Fungi under Drought Stress
LU Wei1, WU Xiaolong1, HU Xianchun1, HAO Yong2*, LIU Chunyan1*
1. College of Horticulture and Gardening, Yangtze University, Jingzhou 434025, China;
2. College of Urban Construction, Yangtze University, Jingzhou, 434023, China
Abstract: To explore the mechanism of arbuscular mycorrhizal (AM) fungi on the growth and physiological characteristics of tea plants under drought stress, tea cultivar ‘Fuding Dabaicha’ was used as experimental material to inoculate with or without (Claroideoglomus etunicatum) under well-watered and drought stress. Plant growth performance, photosynthesis, osmotic regulation and stomatal aperture were determined to investigate the effect of AMF on tea seedlings under different duration of DS (4 weeks, 6 weeks and 8 weeks). The results show that under well-watered and drought stress conditions, AMF inoculation significantly promoted plant growth performance, increased the shoot and root biomass, improved the root system architecture, in particularly increased total root length, secondary and tertiary lateral root numbers, and leaf osmotic potential by 20.00%-38.77%, 48.90%-163.33%, 60.00%-442.86%, 29.77%-41.24%, decreased the stomatal density and relative conductance under drought stress by 16.00%-42.37% and 2.21%-6.67% respectively. The effects were more significant under drought stress. Whereas, drought stress significantly inhibited the root AMF colonization and plant growth, as evidenced by impaired establishment of root system architecture, reduced leaf chlorophyll content, maximum light quantum effect (QY_max), leaf relative water content, stomatal aperture and osmotic potential, etc. AMF inoculation could significantly alleviate this inhibitory effect, improve the physiological response of tea plants under drought stress and thus promote tea plant growth. The results indicate that AMF could promote the absorption of water and nutrients, alleviate the damage of drought stress and improve the drought resistance of tea seedlings by improving root morphology, promoting the water retention and photosynthetic capacity, adjusting the stomatal and osmotic balance, and the promotion effect of AMF became more significant with the extension of drought time.
Keywords: tea plant growth, physiological character, AMF, drought stress
茶樹[Camellia sinensis (L.) O. Kuntze]是山茶科山茶屬的多年生常綠木本植物,是我國重要的園藝及經濟作物。茶樹起源于我國西南部,具有喜濕怕澇的特性,對土壤水分含量十分敏感,土壤缺水或漬水都會影響茶樹的生長、代謝、產量和品質[1]。近年來,高溫干旱等極端天氣頻發(fā),嚴重影響著茶樹生長和茶產業(yè)的發(fā)展。
干旱是影響全球農業(yè)可持續(xù)發(fā)展的限制性因素之一,嚴重影響植物的生長發(fā)育。我國多數茶產區(qū)氣候條件為四季分明,降雨量周年分布不均,夏季高溫少雨、日照強烈,許多茶區(qū)都會出現不同程度的干旱。一旦遭受干旱,茶樹葉片便會逐漸變黃卷曲、出現褐斑、老嫩葉焦枯脫落,嚴重干旱時會使茶樹整體缺水,枝條枯死甚至整株死亡。不僅如此,干旱還會使茶樹體內活性氧增加,葉綠素含量和相對含水量降低,氣孔關閉,從而限制茶樹光合速率[2]。此外,在干旱脅迫下,茶葉中總蛋白、淀粉和雙糖含量降低,可溶性糖、可溶性蛋白和纖維素含量增加[3],嫩葉中茶多酚含量減少,兒茶素含量下降,從而影響茶葉的產量和品質[1]。因此,探索茶樹耐旱性機制,提高茶樹的抗逆栽培,對茶產業(yè)的可持續(xù)發(fā)展具有重要的意義。
叢枝菌根(Arbuscular mycorrhizal,AM)真菌是土壤中的一類有益微生物,它能侵染植物根系并與超過80%的陸生植物建立共生關系。研究證實,AM真菌強大的根外菌絲能夠幫助植物根系吸收根系以外區(qū)域的水分和養(yǎng)分,從而改善宿主的營養(yǎng)狀況,促進植株的生長發(fā)育[4]。AM真菌還能夠通過提高宿主植物體內葉綠素含量、光合酶活性提高植物的光合性能,進而提高植物固碳能力、改善其源庫關系[5]。大量研究表明,AM真菌在提高植物抗逆性方面具有廣闊的應用前景,可通過調控滲透調節(jié)物質的含量[6]、提高抗氧化酶活性[7]、調節(jié)植物激素水平[8]、調控植物脅迫相關基因表達水平[9]等來提高宿主植物的抗逆性。在逆境脅迫下,AM真菌還可以通過改善宿主植物的根系構型、保護光系統(tǒng)、增強抗氧化系統(tǒng),改善植株的生理生化特性,提高宿主植物對干旱的適應性,進而提高產量和品質[10]。
與大多數陸生植物一樣,茶樹根際具有極為豐富的AM真菌資源。前人研究發(fā)現,在野生茶樹和栽培茶樹的根際中存在多種叢枝菌根物種,在自然栽培條件下無梗囊霉屬(Acaulospora)、巨孢囊霉屬(Gigaspora)、球囊霉屬(Glomus)、近明球囊霉屬(Claroideoglomus)和盾巨孢囊霉屬(Scutellospora)的AM真菌均能與茶樹根系形成良好共生體[11]。我國茶園土壤中的AM真菌資源也十分豐富,目前,已報道鑒定的AM真菌達36種,占世界已知AM真菌種類數的17.4%[12]。茶樹為少根毛植物,AM真菌能在茶樹根際形成大量外生菌絲,從而擴展根系吸收面積,促進茶樹對水分和營養(yǎng)元素的吸收利用。王玉娟等[13]研究發(fā)現,接種幼套球囊霉(Glomus etunicatum)后,茶樹葉片數量和各部分(葉片、莖、根系)生物量顯著增加,茶樹葉片蔗糖、葡萄糖、果糖、兒茶素、氨基酸和茶多酚的含量提高,表明AM真菌在茶樹生長和改善茶葉品質方面有一定的促進作用。此外,研究發(fā)現AM真菌能促進水分脅迫下茶樹的生長,增強水分脅迫下茶樹葉片中超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)及抗壞血酸過氧化物酶(APX)的活性,緩解茶樹葉片受氧化程度[14]。這表明AM真菌在緩解逆境脅迫,提高茶樹抗逆性方面也有重要作用。
雖然關于AMF調控茶樹生長的一系列研究已經開展,但是對不同時長干旱脅迫下AM真菌對茶樹生長及抗性方面的研究較少,相關機理解析還不夠深入。鑒于此,本研究以福鼎大白茶(Camellia sinensis cv. ‘Fuding Dabaicha’)為試驗材料,在正常水分和干旱脅迫下接種AM真菌,分析其對不同時長干旱脅迫下茶樹實生苗生長及生理特性的影響,為探究AM真菌增強茶樹抗旱性機理及茶樹高效栽培提供理論依據。
1 材料與方法
1.1 試驗材料
以福鼎大白茶實生苗為試驗材料,茶種由貴州省農科院茶葉研究所提供。茶種經75%酒精消毒10 min后置于預先經過高壓滅菌(121 ℃,0.1 MPa,2 h)的河沙中,于晝/夜溫度28 ℃/20 ℃,相對濕度為70%的培養(yǎng)箱中催芽。供試AM真菌為幼套近明球囊霉(Claroideoglomus etunicatum),菌種購于中國叢枝菌根真菌種質資源庫(BGC),經白三葉草(Trifolium repens L.)擴繁3個月(每克孢子數約達43個)后用于本研究。選取長勢一致的2葉齡無菌茶苗移栽至塑料盆(盆口內徑×盆底內徑×盆高=15 cm×10 cm×13 cm)中,盆中裝有預先經過高壓蒸汽滅菌消除基質中土著真菌孢子的栽培基質(V土︰V河沙=1︰1),每盆移栽2株茶樹實生苗。
1.2 試驗設計
本研究采用雙因素完全隨機試驗設計。因素一為水分處理,分別為正常水分(田間最大持水量的75%,WW)和干旱脅迫(田間最大持水量的55%,DS)。因素二為AM真菌接種,即在每個水分梯度下分別設置接種C. etunicatum(+AMF)與不接種(-AMF)處理。接種AM真菌的處理在移栽時采用“分層接種法”(即分別在盆土1/2和1/3高度處加入AM真菌菌種,并與盆栽基質混勻)進行接種處理,每盆接種80 g菌劑(包括侵染的根段、AM真菌菌絲的孢子和河沙等),活孢子數量約為3 440個。不接種AM真菌的處理在移栽時加入相同質量經高壓滅菌后的菌種作為對照。試驗共4個處理,每個處理重復6次,共24盆,隨機排列。
于AM真菌接種8周后進行干旱處理,隨機選取一半接種AM真菌(+AMF)和不接種(-AMF)處理茶苗進行干旱處理,另一半則繼續(xù)維持正常水分條件。干旱處理于每天18:00通過稱重法[13]計算基質中的水分含量,并補充相應水分,維持穩(wěn)定的含水量。所有試驗材料均于水分處理4周、6周、8周后分批收獲。
1.3 測定方法
每次植株收獲前測定株高、莖粗、葉片數。同時,自上而下選取第4片完全展開的功能葉,采用FluorCam封閉式熒光成像儀(RAP-FLUO高通量植物熒光表型檢測平臺,武漢谷豐光電科技有限公司)測定葉綠素熒光參數,獲得最大光量子效應(QY_max)、穩(wěn)態(tài)光量子效應(QY_Lss)、穩(wěn)態(tài)非光化學熒光淬滅(NPQ_Lss)等參數。隨后參照王學奎[15]的方法測定葉綠素含量。
植株收獲時將茶樹實生苗從試驗盆中完整取出,分成地上部和地下部,同時收集根際土。小心將根系清洗干凈,吸干表面水分后立即稱取地上部鮮重和地下部鮮重。隨后將完整的根系置于根盤中,采用Epson Perfection V700 Photo Dual Lens System(J221A,Indonesia)對茶樹根系進行掃描,采用WinRHIZO(2007)分析根系構型參數。隨后將根系平鋪在實驗臺上,人工統(tǒng)計各級側根數。隨機選取各級側根根系,去掉根尖,剪成1~2 cm長根段置于FAA固定液中,經曲利苯藍染色后參考Phillips等[16]的方法測定菌根侵染率。土壤菌絲長度參考Bethlenfalvay等[17]的方法,在生物顯微鏡下鏡檢,記錄土壤菌絲長度。
葉片相對含水量參照Bajji等[18]的方法測定。相對電導率、滲透勢以及氣孔密度的測量參考孫琪璐[19]的方法:相對電導率采用電解質外滲法測定;取植物組織液10 μL用露點滲透壓儀測定其滲透勢;氣孔密度測定,自頂部向下選取第3片完全成熟的功能葉,切成體積為1 mm3的小塊,放于預冷的5%戊二醛固定液中,4 ℃過夜,隨后在葉片下表皮涂抹指甲油,自然風干后將指甲油剝離,置于載玻片上于顯微鏡下觀察氣孔形態(tài),在光學顯微鏡40×物鏡下統(tǒng)計每個視野中的氣孔數目,并測量視野中的氣孔器長度、寬度。參照楊妮等[20]的方法計算氣孔開度。
1.4 統(tǒng)計分析
運用SAS10.1.3v軟件(SAS Institute Inc.,Cary,NC,USA)進行雙因素差異分析(Two way ANOVA),并用Duncan多重比較法進行多重比較分析(P<0.05)。
2 結果與分析
2.1 水分處理前茶樹實生苗生長狀況
水分處理前,接種AM真菌(+AMF)顯
著促進了茶樹實生苗生長,株高、莖粗、葉片數及各部分生物量分別增加了19.95%、6.17%、54.76%、68.29%、75.00%和139.06%(表1)。
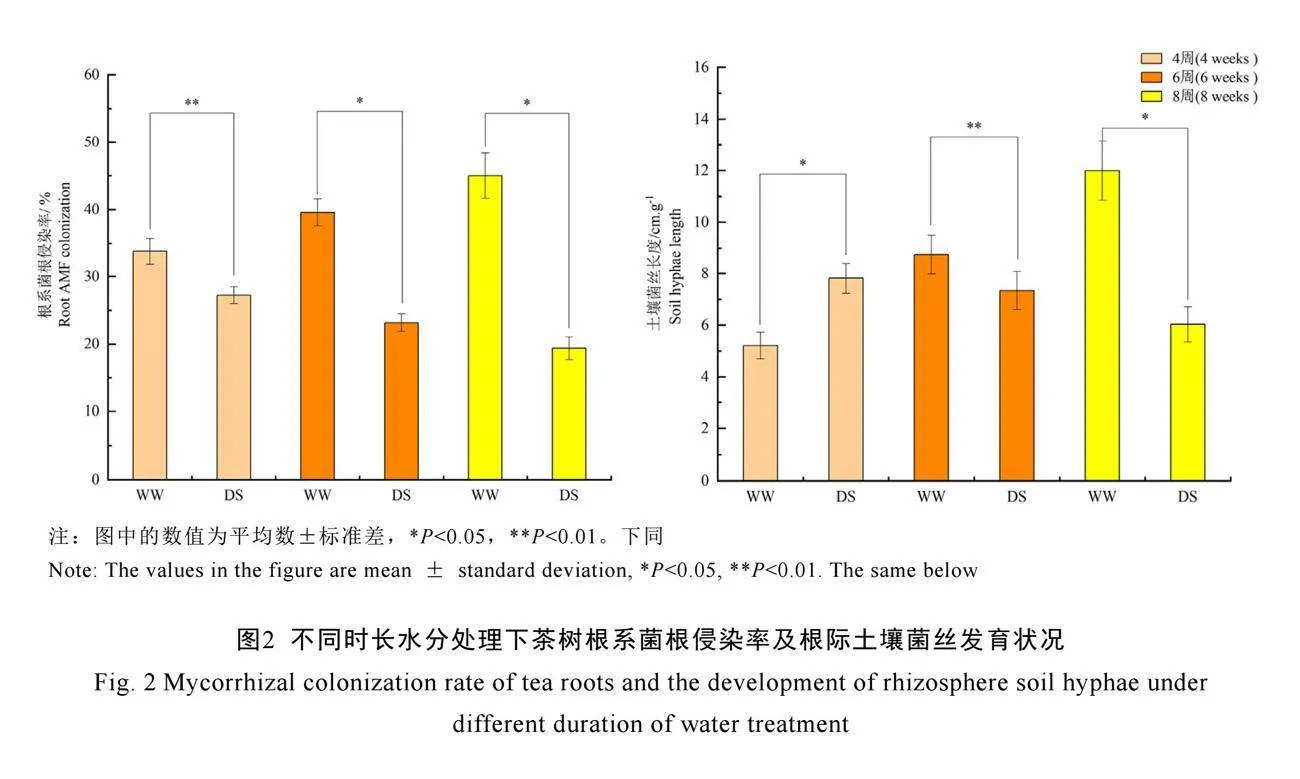
2.2 不同水分處理下茶樹菌根發(fā)育狀況
不論在正常水分還是在干旱脅迫下,C. etunicatum均能侵染茶樹根系(圖1),侵染率為19.4%~45.0%(圖2)。在正常水分下,茶樹根系菌根侵染率和根際土壤菌絲長度均隨著處理時間的延長而不斷增加,在8周時,分別達到45.01%和11.99 cm·g-1。然而,干旱脅迫下茶樹根系菌根侵染率和土壤菌絲長度會隨著處理時間的延長而不斷降低,并在8周時顯著降低(P<0.05),與同時期正常水分組相比,分別降低了57.78%和50.00%。表明干旱脅迫會抑制AM真菌對茶樹根系的侵染,限制菌絲在土壤中的擴繁,進而影響AM真菌促生功能的發(fā)揮。
2.3 AM真菌對不同時長水分處理下茶樹實生苗生長的影響
由表2可知,在不接種AM真菌(–AMF)處理下,干旱脅迫僅在6周和8周時顯著降低了株高,在4周和8周時顯著降低了葉片數(P<0.05),對莖粗無顯著影響。但不同干旱脅迫時長均顯著降低了茶樹葉片、莖和根系的生物量。與不接種AM真菌相比,接種AM真菌顯著增加了正常水分下4周、8周時茶樹的株高,以及4周和6周時的莖粗;增加干旱脅迫下6周、8周時茶樹的株高以及8周時的葉片數。此外,接種AM真菌還顯著增加了正常水分和
干旱脅迫下茶樹莖干鮮重以及8周時根系和葉片鮮重,且對干旱脅迫下各時長葉片、莖和根系的生物量增加尤為顯著(P<0.05),在4周時分別增加35.29%、50.00%和103.79%;在6周時分別增加25.00%、37.14%和138.46%;在8周時分別增加28.00%、42.11%和74.24%(表2)。
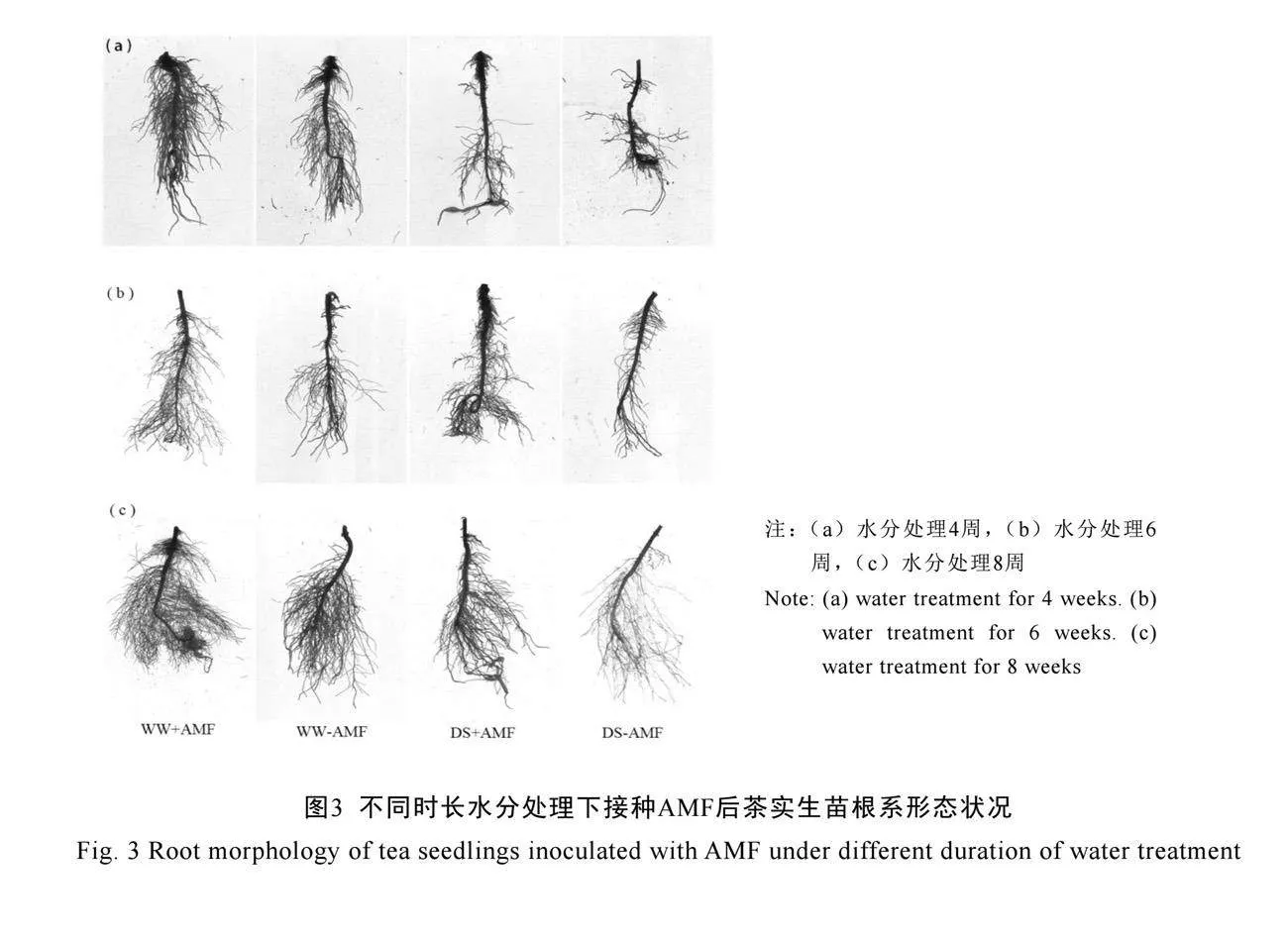
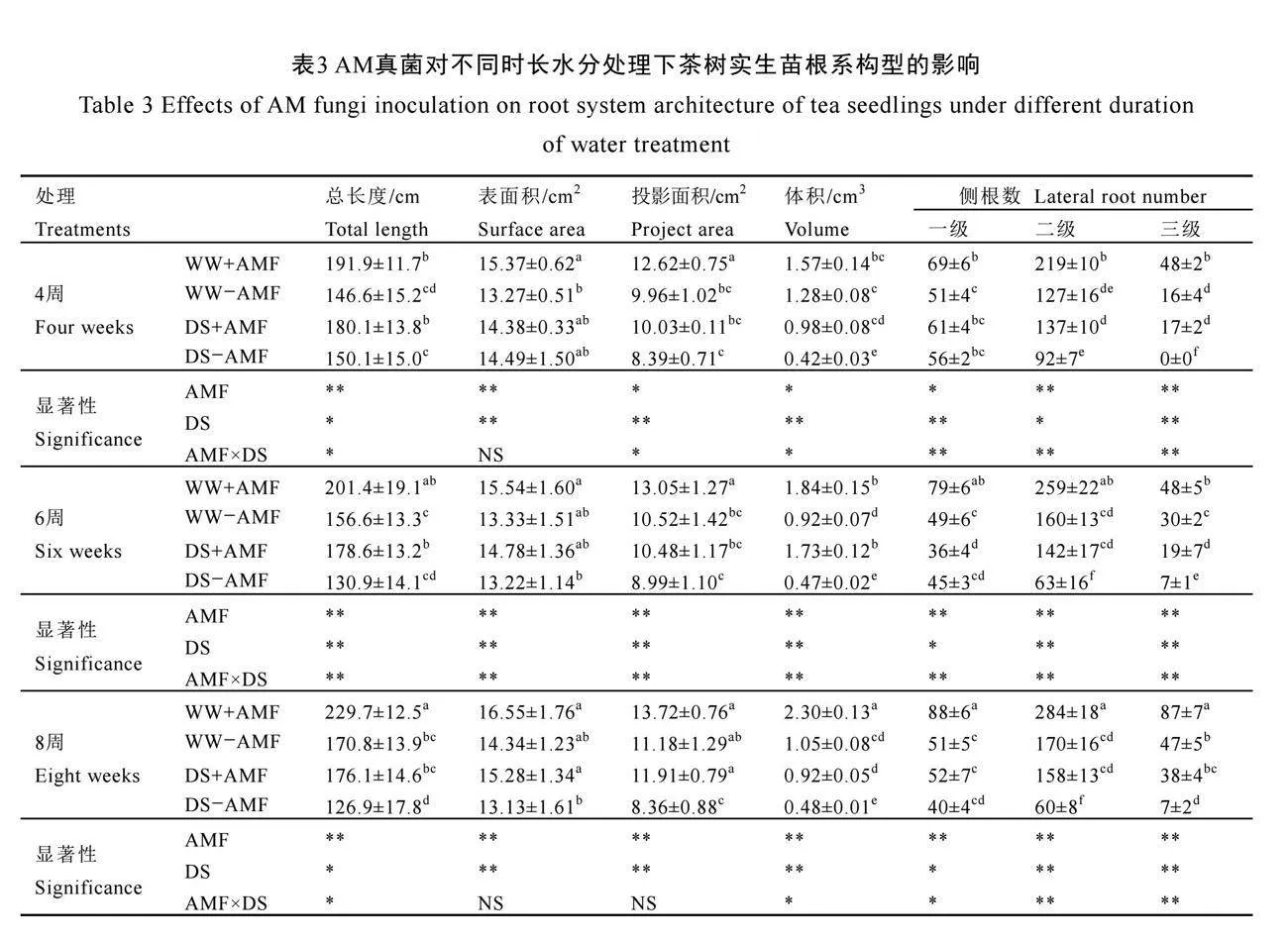
2.4 AM真菌對不同時長水分處理下茶樹實生苗根系構型的影響
不同水分處理下AM真菌對茶樹實生苗根系構型的影響如圖3所示。不接種AM真菌(-AMF)處理的茶苗,在干旱脅迫下僅在第4周、6周時的根系體積、三級側根數量以及6周時二
級側根數量(P<0.05)顯著降低;根系的發(fā)育在第8周時才被顯著抑制,根系總長度降低、根系體積及各級側根數量減少(表3)。與未接種AM真菌(-AMF)相比,接種AM真菌(+AMF)后顯著促進了茶樹根系發(fā)育。在正常水分下,接種AM真菌(+AMF)顯著增加了根系總長度及一級、二級和三級側根數量,在4周時分別增加了30.9%、35.3%、72.4%和200.0%,6周時分別增加了28.7%、61.2%、61.9%和60.0%,在8周時分別增加了34.5%、72.5%、57.1%和85.1%;在干旱脅迫下,接種AM真菌(+AMF)顯著增加了茶樹根系總長度、根系體積、二級側根和三級側根的數量(P<0.05),在4周時分別增加了20.0%、133.3%、48.9%和170.0%,在6周時分別增加了36.4%、268.1%、125.4%和171.43%,在8周時分別增加了38.8%、91.7%、163.3%和442.9%。由此可見,AM真菌對干旱脅迫下根系構型的促進作用更為顯著,能有效緩解干旱脅迫對根系發(fā)育的抑制作用。
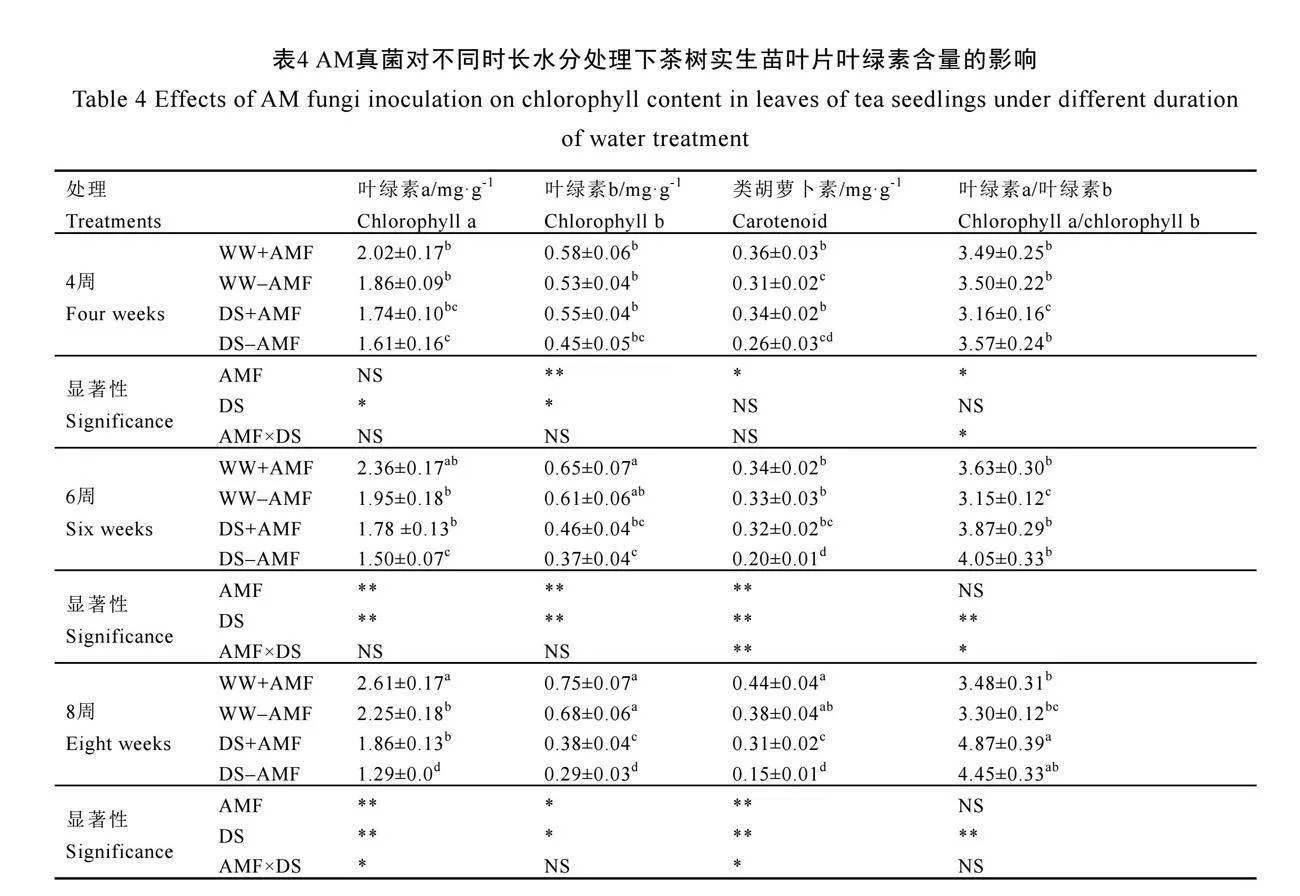
2.5 AM真菌對不同時長水分處理下茶樹葉片葉綠素含量的影響
由表4可知,在未接種AM真菌(-AMF)下,干旱脅迫4周時僅顯著降低了茶樹葉片葉綠素a的含量,在6周和8周時顯著降低了葉綠素a、葉綠素b和類胡蘿卜素的含量,卻顯著提高了葉綠素a/葉綠素b的值(P<0.05)。與不接種AM真菌相比,在4周時,接種AM真菌(+AMF)顯著增加了茶樹葉片類胡蘿卜素的含量,在正常水分和干旱脅迫下分別增加了16.13%和30.77%;與此同時,顯著增加了干旱脅迫下葉綠素a/葉綠素b的值(P<0.05)。在6周時,接種AM真菌(+AMF)顯著提高了干旱脅迫下茶樹葉片葉綠素a和類胡蘿卜素的含量,以及正常水分下葉綠素a/葉綠素b的值,對正常水分下色素含量和干旱脅迫下葉綠素a/葉綠素b的值無顯著影響。在8周時,接種AM真菌僅顯著增加了正常水分下葉綠素a的含量以及干旱脅迫下葉綠素a、葉綠素b和類胡蘿卜素的含量,與不接種AM真菌相比,分別增加了16.00%、44.19%、31.03%和106.7%(P<0.05)。
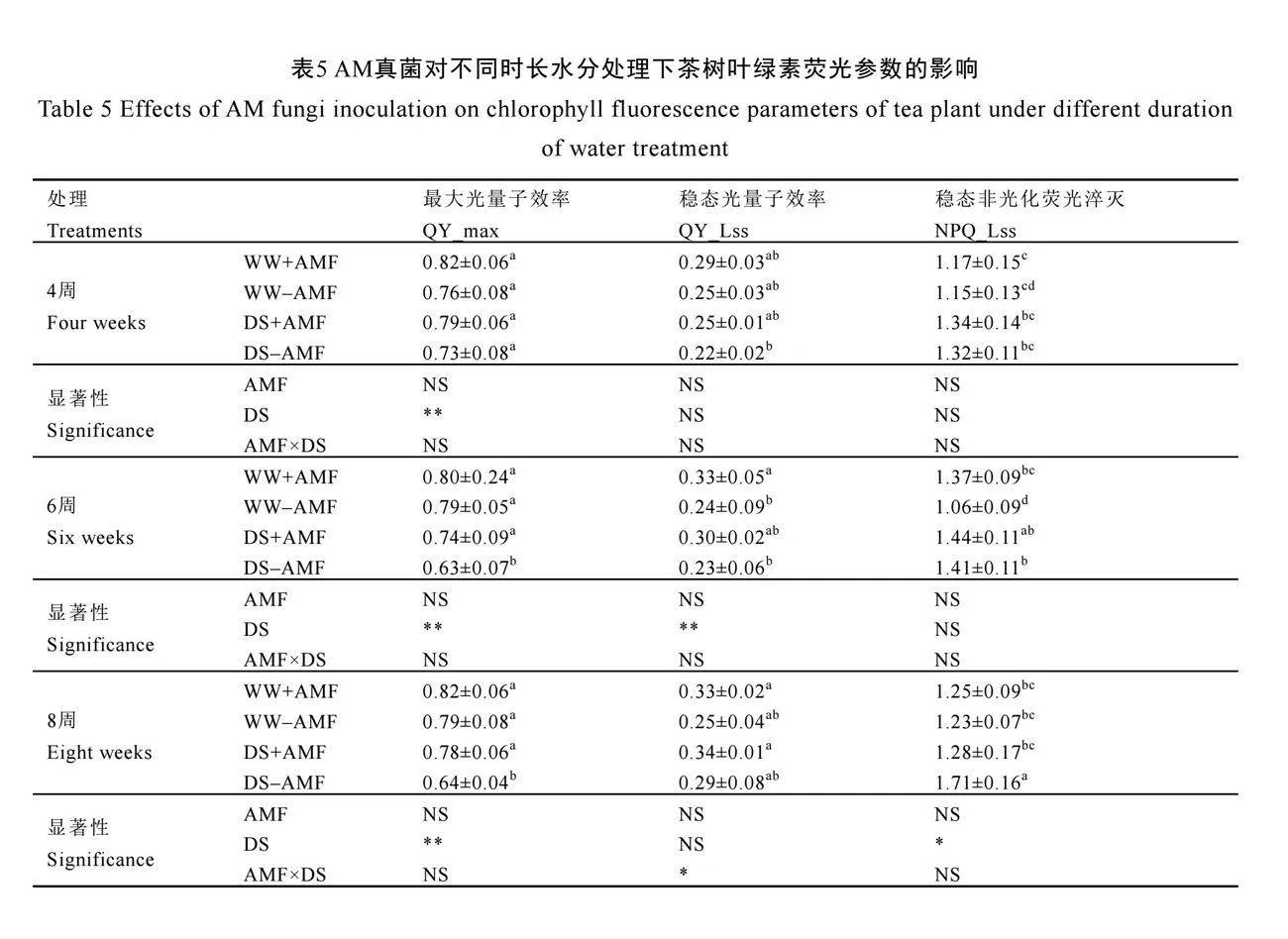
2.6 AM真菌對不同時長水分處理下茶樹葉綠素熒光參數的影響
在未接種AM真菌(-AMF)下,干旱脅迫(DS)顯著降低6周和8周時茶樹葉片QY_max,與正常水分組相比分別降低了20.25%、18.99%,但是卻顯著增加了NPQ_Lss,分別增加了33.02%和39.02%(P<0.05,表5)。與不接種AM真菌相比,接種AM真菌(+AMF)顯著增加了干旱脅迫(DS)下6周和8周時茶樹葉片QY_max(P<0.05),分別增加了17.46%和21.88%,同時降低了在8周時茶樹葉片NPQ_Lss,降低了25.15%;但是在正常水分下,接種AM真菌顯著增加了6周時茶樹葉片QY_Lss和NPQ_Lss(P<0.05),對其他熒光參數無顯著影響(表5)。
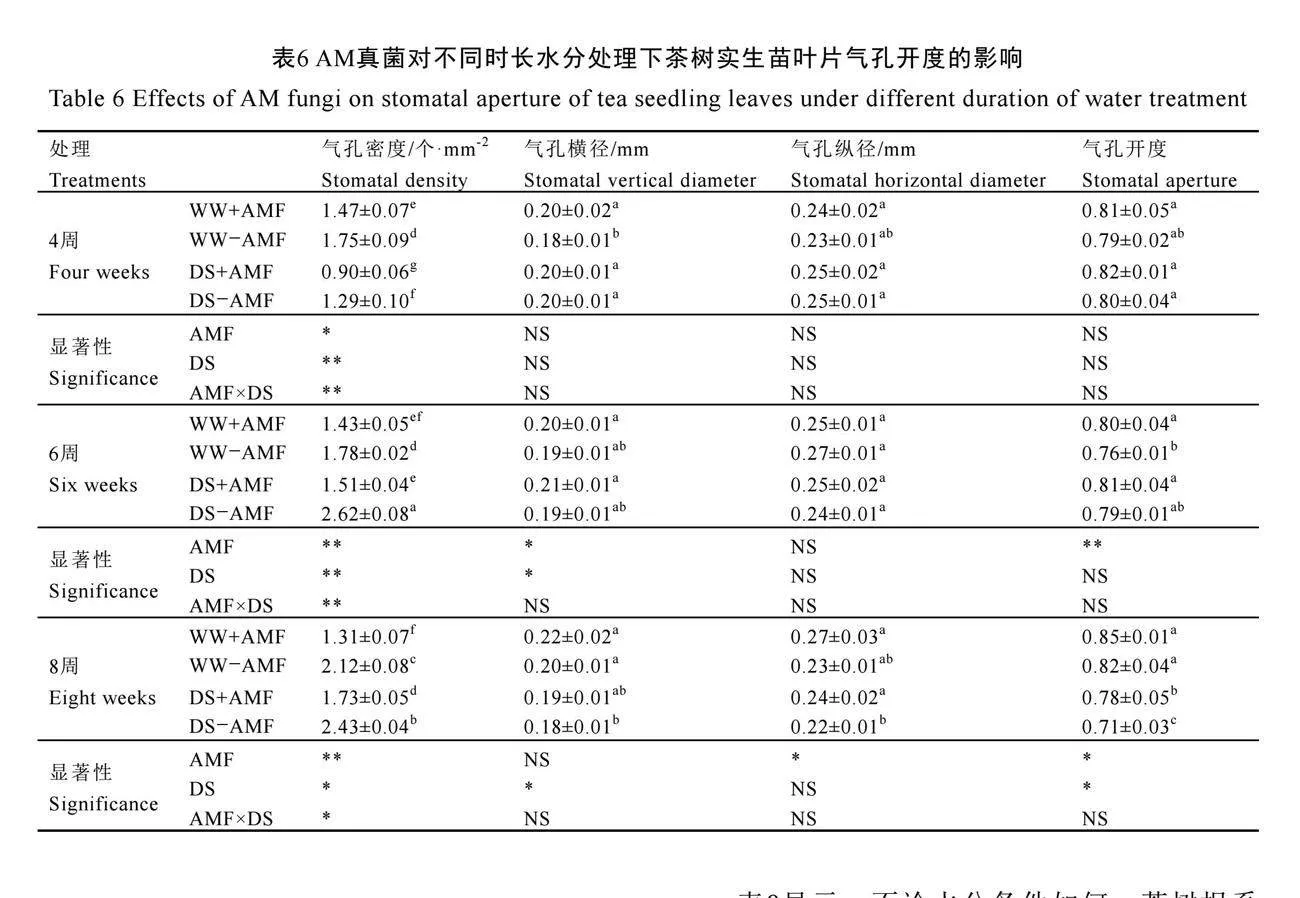
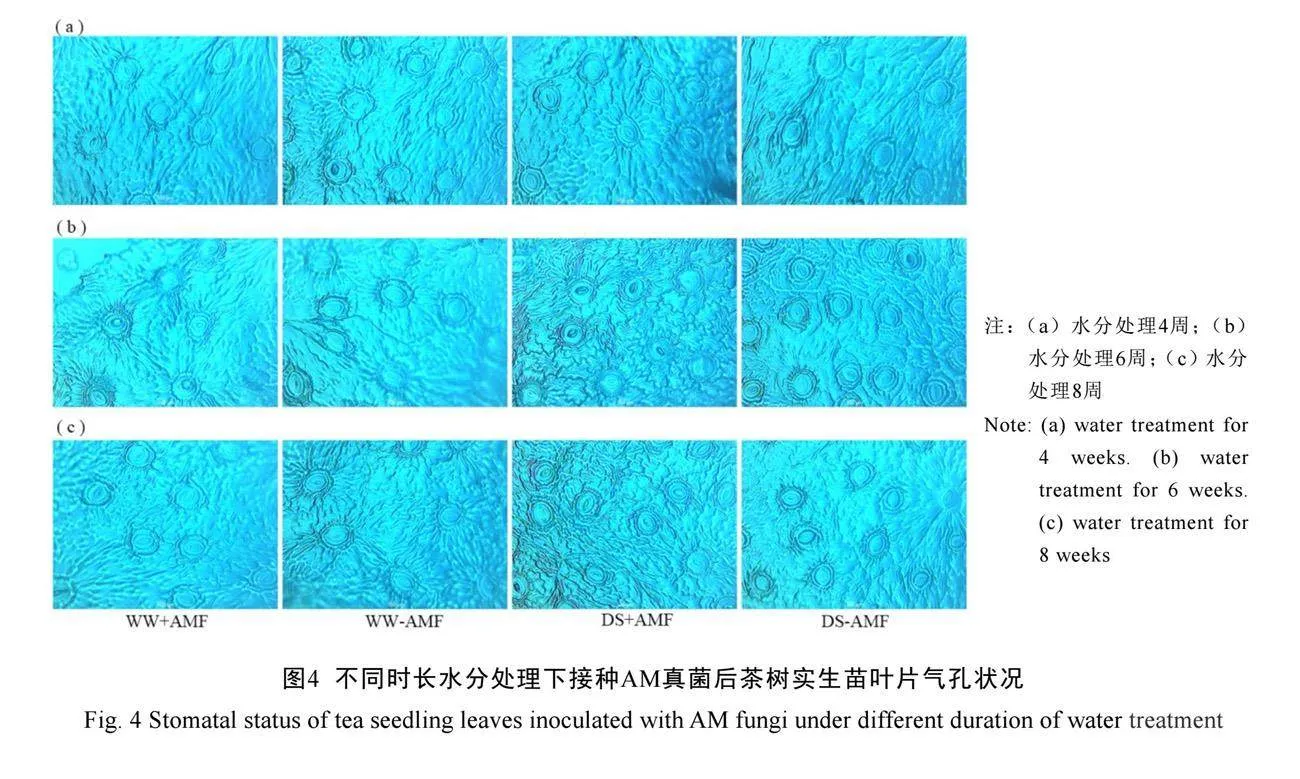
2.7 AM真菌對不同時長水分處理下茶樹實生苗葉片氣孔開度的影響
不同時長水分處理下,茶樹實生苗葉片氣孔狀況如圖4所示。在未接種AM真菌(-AMF)下,干旱脅迫4周時葉片氣孔密度顯著降低,氣孔橫徑顯著增加;6周時,僅顯著增加了氣孔密度;8周時,氣孔密度顯著增加,氣孔橫徑和開度顯著降低(P<0.05),且干旱脅迫處理對氣孔縱徑無顯著影響(表6)。與不接種AM真菌相比,接種AM真菌(+AMF)顯著降低了正常水分和干旱脅迫下茶樹葉片氣孔密度(P<0.05),在4周、6周、8周時分別降低了16.00%、19.66%、38.21%和30.23%、42.37%、28.81%。此外,接種AM真菌還顯著增加了4周時正常水分下的氣孔橫徑和8周時干旱脅迫下的氣孔縱徑,以及6周時正常水分處理下和8周時干旱脅迫下的氣孔開度(表6)。
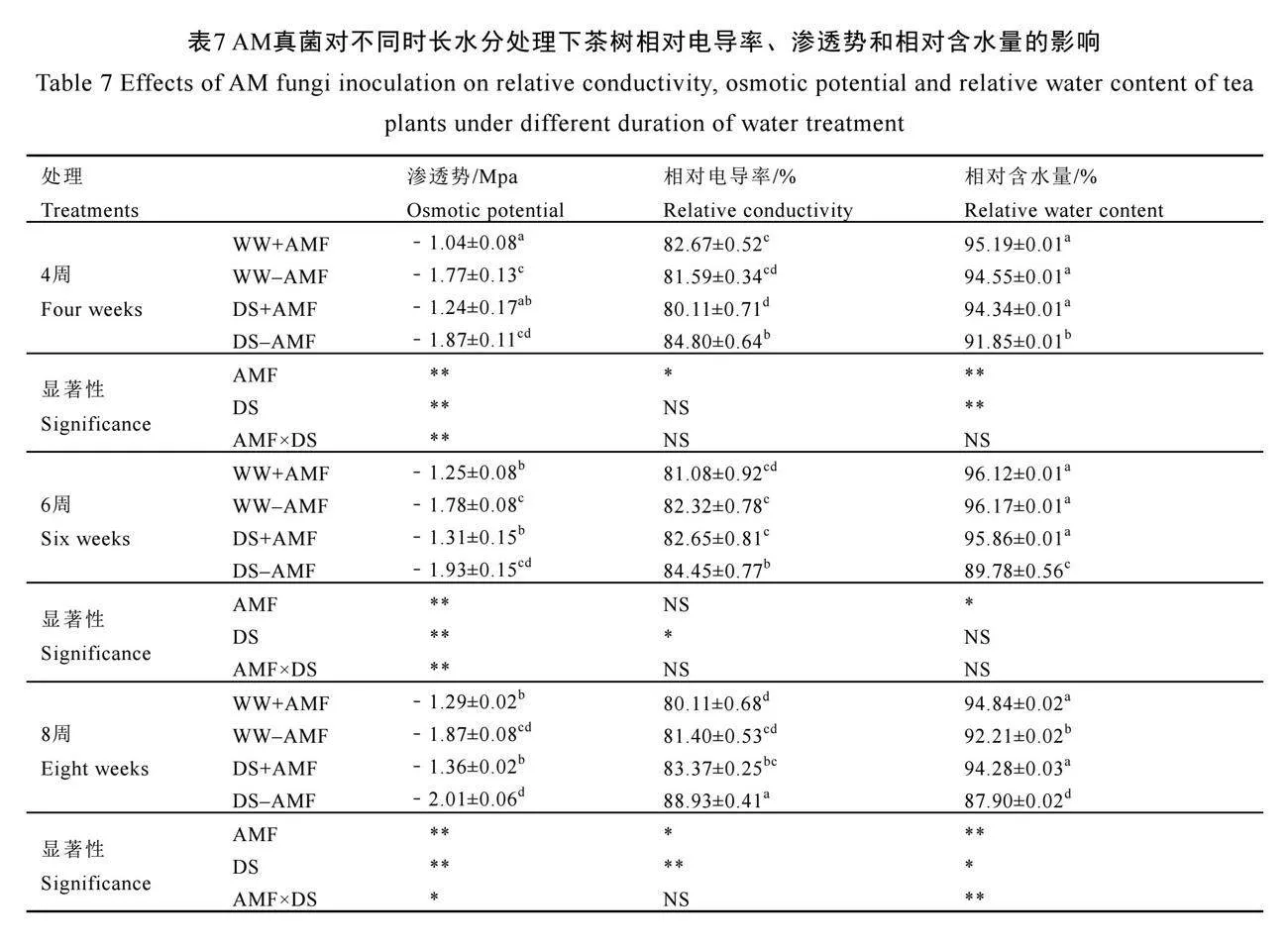
2.8 AM真菌對不同時長水分處理下茶樹相對電導率、滲透勢和相對含水量的影響
由表7可知,在未接種AM真菌(-AMF)的茶苗中,干旱脅迫顯著增加了茶樹相對電導率,但是卻顯著降低了茶樹葉片相對含水量(P<0.05),對滲透勢無顯著影響。與未接種AM真菌相比,接種AM真菌(+AMF)顯著增加了正常水分(WW)和干旱脅迫下滲透勢,在4周、6周、8周時分別增加了41.24%、29.78%、31.02%和33.69%、32.12%、32.34%,表明AM真菌能夠改善植物的水分狀態(tài),增強植株在脅迫環(huán)境下的抵抗能力。與此同時,接種AM真菌還顯著降低了干旱脅迫(DS)下4周、6周、8周時茶葉電導率,但是卻顯著增加了葉片相對含水量,表明AM真菌的接種能減少細胞物質滲出,保護細胞膜的完整性,從而改善葉片水分狀況。
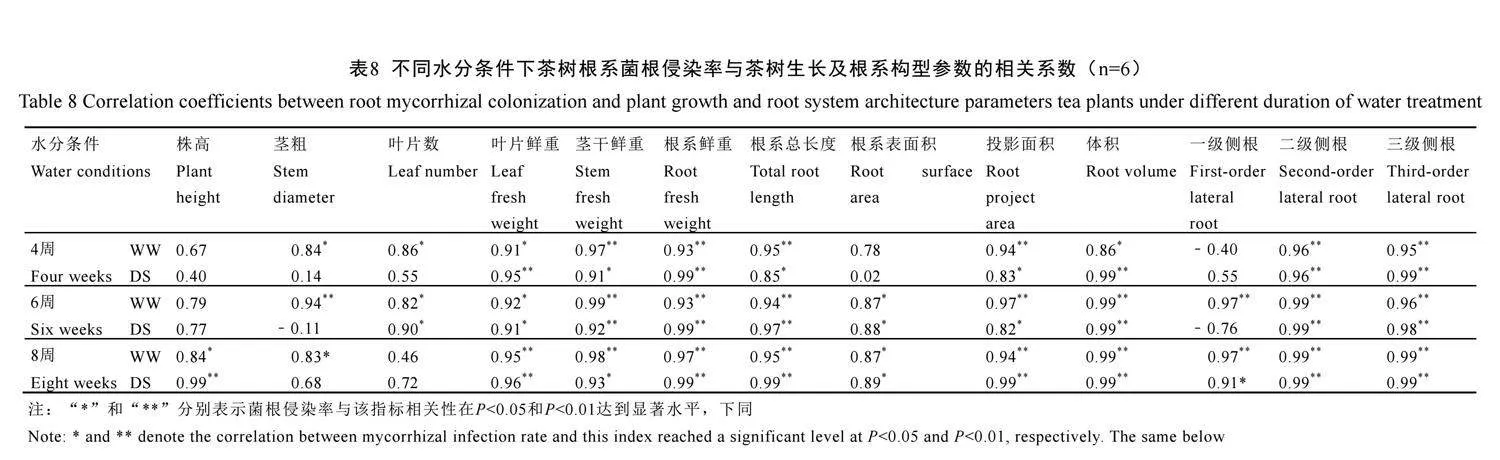
2.9 茶樹根系菌根侵染率與茶樹生長及根系構型參數相關關系
表8顯示,不論水分條件如何,茶樹根系菌根侵染率在4周、6周、8周時均與生物量(葉片、莖干、根系鮮重)、根系總長度、投影面積、體積、二級側根和三級側根的數量呈顯著或極顯著正相關(P<0.05或P<0.01);在6周、8周時均與根系表面積呈顯著正相關(P<0.05)。除此之外,水分處理4周時,在正常水分下菌根侵染率與莖粗、葉片數呈顯著正相關(P<0.05)。6周時,在正常水分下,菌根侵染率與莖粗、葉片數和一級側根數量呈顯著正相關,在干旱脅迫下與葉片數呈顯著正相關(P<0.05)。8周時,在正常水分下,茶樹根系菌根侵染率與株高,莖粗、一級側根呈顯著正相關,在干旱脅下與株高和一級側根數量呈極顯著負相關(P<0.01)。表明水分條件和處理時間均對茶樹根系侵染率與生長和根系構型建立之間產生了顯著的影響,尤其是在生物量、根系總長度和側根發(fā)育方面效果極為顯著,表明AM真菌侵染能顯著促進茶樹生長,即使在干旱脅迫下,也能促進茶樹生長。
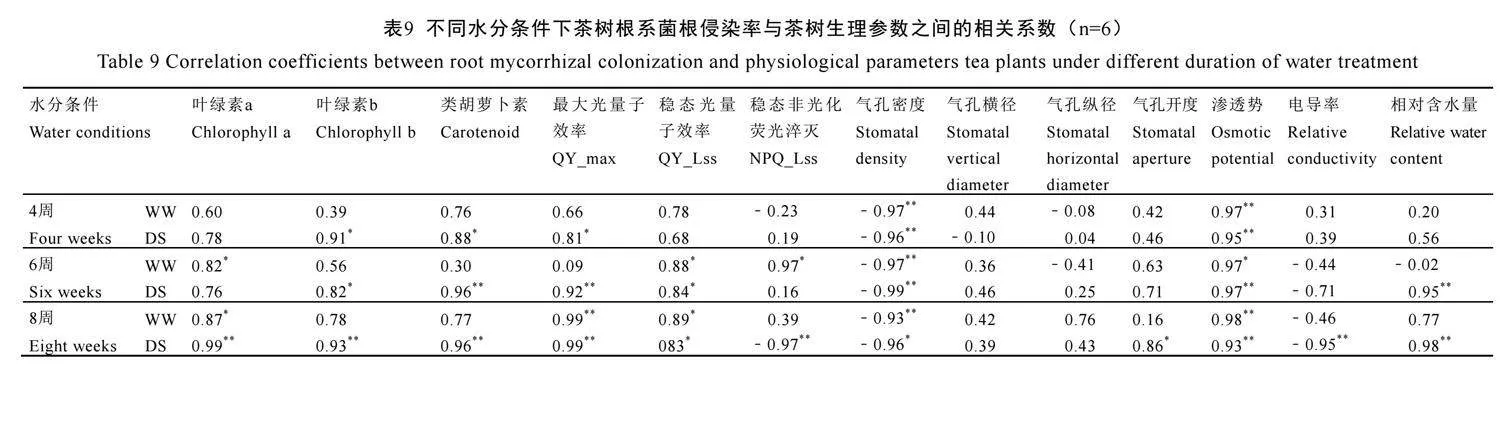
2.10 茶樹根系菌根侵染率與茶樹生理參數之間相關關系
由表9可見,不論水分條件如何,茶樹根系菌根侵染率在4周、6周、8周時均與氣孔密度呈極顯著負相關(P<0.01)。此外,水分處理4周時,在正常水分下菌根侵染率僅與滲透式呈極顯著正相關(P<0.01),在干旱脅迫下與葉綠素b、類胡蘿卜素、QY_max(P<0.05)、滲透式(P<0.01)呈顯著或極顯著正相關。6周時,在正常水分下菌根侵染率與葉綠素a、QY_Lss、NPQ_Lss、滲透式呈顯著正相關(P<0.05),在干旱脅迫下與葉綠素b、QY_Lss(P<0.05)、類胡蘿卜素、QY_max、滲透式、相對含水量(P<0.01)呈顯著或極顯著正相關。8周時,在正常水分下菌根侵染率與葉綠素a、QY_Lss(P<0.05)、QY_max、滲透式(P<0.01)呈顯著或極顯著正相關,在干旱脅迫下與QY_Lss、氣孔開度(P<0.05)、葉綠素a、葉綠素b、類胡蘿卜素、QY_max、滲透式、相對含水量(P<0.01)呈顯著或極顯著正相關,但是卻與NPQ_Lss和相對電導率呈極顯著負相關(P<0.01)。表明水分條件和處理時間影響了AM真菌侵染與茶樹生理參數間的相關性,干旱脅迫下AM真菌與這些生理參數之間相關性更強,表明AM真菌侵染有利于改善茶樹對干旱脅迫的生理響應。
3 討論
干旱脅迫是限制茶樹生長發(fā)育的主要因素之一,極端氣候變化導致干旱頻繁發(fā)生,直接影響著茶葉的產量和品質[1]。AM真菌與植物共生產生的大量根外菌絲能夠促進宿主植物對水分和養(yǎng)分的吸收,從而提高植物對干旱脅迫的抵御能力,增強宿主植物對干旱脅迫的適應性[10]。本研究發(fā)現AM真菌能與福鼎大白茶根系建立良好的共生關系,且在正常水分下
侵染率和土壤菌絲長度隨處理時間的延長而提高。在干旱脅迫下,茶樹根系菌根侵染率顯著降低,且隨著干旱脅迫時間的延長而逐漸下降,根際土壤菌絲長度也顯著降低。這與王玉娟等[13]和Liu等[21]在茶樹上的研究結果一致。雖然短時間的干旱處理也會抑制AM真菌的侵染,但宿主植物根系菌根侵染率仍然呈現增長緩慢的趨勢,這主要歸因于宿主植物體內保存的水分以及積累的碳水化合物還能繼續(xù)促進菌絲的發(fā)育[22],然而長期的水分虧缺抑制了孢子的萌發(fā)和菌絲的擴展,以及碳水化合物的積累[23],同時使得宿主植物生長代謝變緩,AM真菌無法從宿主中獲取充足的營養(yǎng)。因此在本研究中持續(xù)的干旱處理導致根系菌根侵染率和土壤菌絲長度下降,這與Ortas等[24]的研究結果一致。
干旱脅迫是抑制植物生長,影響產量和品質的限制性因子。在本研究中我們也得到了相似的結果,發(fā)現干旱脅迫處理顯著抑制了茶樹實生苗的生長,降低了茶樹地上部和地下部生物量。然而接種AMF后顯著改善了不同水分處理下茶苗的生長表型,顯著增加了茶樹植株生物量,且在干旱脅迫下的促進效果更為顯著。這與前人在柑橘[25]上的研究結果一致。相關關系分析也顯示,不論水分狀況為何,茶樹根系菌根侵染率與不同時長水分處理下茶樹生物量呈極顯著正相關,表明干旱脅迫雖然抑制AM真菌與茶樹根系共生關系的建立,但是AM真菌的侵染仍然促進了茶苗的生長,這主要歸因于AM真菌龐大的根外菌絲網絡增強了水分和養(yǎng)分的吸收,改善了宿主植物養(yǎng)分狀況,提高了植株的抗旱能力[26]。此外,根外菌絲還會向土壤中分泌球囊霉素蛋白,不僅能夠促進土壤團聚體的形成從而提高土壤的保水能力,還可為土壤提供充足的有機碳、氮等大量元素,為宿主植物有機物質的積累提供保障[27]。
根系可塑性高,能對周圍環(huán)境變化做出快速響應。然而在眾多環(huán)境因素中,土壤水分狀況和養(yǎng)分含量對根系的生長起決定性作用[28]。在本研究中,干旱脅迫顯著抑制了茶樹根系構型的建立,在側根發(fā)育方面的抑制作用尤為顯著,這與Liu等[21]在茶樹上的研究結果一致。然而,AM真菌作為土壤中的一類有益微生物,對根系構型的影響不言而喻。先前的研究發(fā)現AM真菌可通過調控宿主植物根系構型促進植物生長[21]。本研究發(fā)現,接種AM真菌處理顯著改善茶樹根系構型,無論是在正常水分還是干旱脅迫下,都顯著增加了根系總長度以及各級側根的數量。這與前人在柑橘[27]上的研究結果一致。這可能是因為AM真菌提高了根系對水分和養(yǎng)分的吸收,增強了根系活力,從而促進了根系發(fā)育[29]。此外,AM真菌可能通過影響植物生理和代謝過程,主要表現為改變宿主植物內源激素的水平,促進根系發(fā)育[30]。相關關系分析顯示,不論水分狀況為何,水分處理4周、6周、8周時茶樹根系菌根侵染率與茶樹根系總長度、根系投影面積、二級側根以及三級側根的數量呈顯著正相關,在6周和8周時與根系表面積呈顯著正相關,與6周時正常水分下的一級側根以及8周時的一級側根數量呈顯著正相關,這進一步說明AM真菌侵染在改善茶樹根系構型方面具有積極地作用。良好根系構型是地上部分形態(tài)建成的基礎,這也為本研究中干旱脅迫下AM真菌仍然促進茶樹生長提供了證據。
葉綠素含量和葉綠素熒光參數是反映植物光合能力的主要指標,同時也反映著植物在逆境脅迫下的抵抗能力。在干旱脅迫下,植物體內活性氧的積累會致使膜結構發(fā)生變化,從而抑制葉綠素的合成與積累。本研究結果顯示,干旱脅迫處理4周時茶樹葉片葉綠素a含量顯著降低,6周和8周時葉綠素a、葉綠素b及類胡蘿卜素含量顯著降低,表明干旱脅迫會使得葉片細胞器受到損傷,從而影響葉綠素的合成。在4周時,干旱脅迫并未顯著影響葉綠素b和類胡蘿卜素的含量,這可能是因為短時間的干旱脅迫下,茶樹一方面通過抑制葉綠素類物質的消耗,另一方面通過強化胞質抗氧化系統(tǒng),增強了葉綠素的穩(wěn)定性。此外,在本研究中,接種AM真菌顯著促進了干旱脅迫下4周時類胡蘿卜素、6周時葉綠素a和類胡蘿卜素以及8周時茶樹葉片色素的積累,但在正常水分下僅顯著增加了4周時類胡蘿卜素和8周時葉綠素a的含量,這與Dai等[2]在茶樹上的研究結果一致。相關關系分析結果顯示,茶根系菌根侵染率與4周、6周、8周時葉綠素b和類胡蘿卜素的含量呈顯著正相關,與6周和8周時的葉綠素a呈顯著正相關,表明AM真菌能夠有效緩解干旱脅迫對茶樹葉片光合器官的損傷以及葉綠素的破壞作用,保護細胞器免受損傷,從而通過促進宿主植物葉綠素的合成與積累,增強光合作用,促進茶樹地上部分的形態(tài)建成與植株的發(fā)育。
葉綠素熒光參數反映著植物在不同環(huán)境下的光合反應器的化學性能。先前在茶樹上的研究結果顯示,干旱脅迫會導致葉片的最大光化學效率、實際光化學量子產率和光化學猝滅系數呈現降低,然而接種AM真菌能顯著改善這些熒光參數值[2]。本研究結果顯示干旱脅迫處理6周時顯著降低了茶樹葉片QY_max,增加了NPQ_Lss,這與方必君等[31]在野牡丹上的研究結果一致,表明干旱脅迫會降低光合器官活性以及光能轉換效率。與此同時,接種AM真菌僅顯著增加了干旱脅迫處理6周和8周時QY_max以及6周時NPQ_Lss,這與馬坤等[32]的研究結果一致。相關關系分析顯示,茶樹根系菌根侵染率與干旱脅迫處理4周時的QY_max、6周時和8周時QY_max、QY_Lss呈顯著正相關,表明接種AM真菌能夠增強宿主植物光合器官活性,促進光能轉換,從而減輕干旱脅迫對茶樹幼苗光系統(tǒng)的損失,以此增強植物的光合性能,促進宿主植物生長。
氣孔是植物與外界氣體交換和水汽運輸的通道,同時也調控著植物蒸騰速率的快慢和光合作用的強弱。在干旱脅迫下,植物可通過調節(jié)氣孔的開度和密度從而降低蒸騰作用,減少植物體內水分的散失。本研究結果顯示,干旱脅迫處理4周時,茶苗葉片氣孔密度降低,在6周和8周時氣孔密度增加,且在8周時氣孔開度顯著降低,這與吳學蕤等[33]的研究結果一致。在干旱處理前期氣孔密度降低可能是植物對逆境環(huán)境的適應策略,在干旱脅迫下通過減少葉片氣孔開度來減少水分的蒸騰,同時為保證植物通過葉片能吸收更多的水分而增加氣孔數量。在6周以后,可能是因為干旱脅迫抑制了葉片的生長,使得葉面積變小,從而造成單位面積內密度變大。此外,本研究結果還顯示,接種AM真菌處理顯著增加了6周時正常水分下以及8周時干旱脅迫下茶苗葉片氣孔開度,顯著降低了4周、6周和8周時的氣孔密度,這與劉婷等[34]的研究結果一致。表明AM真菌可通過增加茶苗葉片氣孔開度以獲取更大的氣體交換空間。而氣孔密度的降低可能是由于AM真菌使植物體內水分增加,葉面積增大,從而使茶苗葉片氣孔密度減少。相關分析顯示,不論是在正常水分還是在干旱脅迫下,茶樹根系菌根侵染率與氣孔密度呈顯著負相關,與8周時干旱脅迫下氣孔開度呈顯著正相關,進一步證明AM真菌侵染參與了茶樹葉片氣孔的變化,擴大氣體交換空間,從而促進植物生長,增強植物抗性。
相對電導率和滲透勢反映著植物在逆境脅迫下的傷害程度。在非生物脅迫下,植株細胞膜可能會受到不同程度的損傷,膜透性增大,細胞內電解質外滲使細胞相對電導率增大,滲透勢降低[35]。本研究結果顯示,干旱脅迫顯著增加了茶樹的相對電導率,隨著干旱脅迫時間的延長,相對電導率逐漸增大,這與李津津等[36]在核桃上的研究結果一致,說明干旱脅迫處理對茶樹葉片細胞膜造成了一定的損傷。然而,AM真菌侵染后,干旱脅迫下的茶樹葉片相對電導率顯著降低,但無論是在正常水分還是在干旱脅迫下,AM真菌侵染均顯著提高了茶樹葉片滲透勢,這與前人[6]的研究結果一致。相關關系分析顯示,不論水分狀況如何,茶樹根系菌根侵染率均與4周、6周和8周時的滲透式呈顯著正相關,與8周時干旱脅迫下相對電導率呈顯著負相關,表明AM真菌具有緩解植株細胞膜過氧化,維持細胞膜穩(wěn)定性的作用,在一定程度上增強植株的抗旱能力,在逆境脅迫下維持宿主植物健壯生長。
相對含水量是衡量植物體內水分狀況的重要指標,反映了植株體內水分虧缺程度和體內代謝活性。AM真菌與宿主植物共生后,其龐大的菌絲網絡能夠幫助植株吸收更多的水分,同時還能改善根系構型以增加宿主植物根系吸收面積,使植株保持良好的水分狀況[26]。本研究中,干旱脅迫顯著降低了茶樹葉片相對含水量,但是接種AM真菌顯著提高了茶樹葉片相對含水量,這與前人在枳[37]上的研究結果一致。相關關系分析結果顯示,在6周和8周時,茶根系菌根侵染率與干旱脅迫下茶葉片相對含水量呈極顯著的正相關關系,表明AM真菌在促進水分吸收,改善植物體內水分的轉運與分布方面具有重要作用。水分條件的改善不僅為宿主植物的生長提供了非常有利的環(huán)境條件,同時在逆境脅迫下還能增強植株對不良環(huán)境的適應能力。
綜上所述,AM真菌能通過促進干旱脅迫下福鼎大白茶根系形態(tài)的建立、促進側根的發(fā)生,改善植株體內水分的運輸與分布,從而促進茶樹生長。此外,AM真菌還能通過改善茶樹葉片色素的合成與積累、氣孔的數量與開度來改善氣體交換空間、增強最大光量子效應改善光合,同時通過調節(jié)水勢緩解活性氧傷害,從而提高植物的生產性能,提高茶樹對干旱脅迫的抵御能力。
參考文獻
[1] Zhang S Y, Liu J J, Zhong G X, et al. Genome-wide identification and expression patterns of the C2H2-Zinc finger gene family related to stress responses and catechins accumulation in Camellia sinensis (L.) O. Kuntze [J]. International Journal of Molecular Sciences, 2021, 22(8): 4197-4214.
[2] Dai F J, Rong Z Y, Wu Q S, et al. Mycorrhiza improves plant growth and photosynthetic characteristics of tea plants in response to drought stress [J]. Biocell, 2022, 46(5): 1339-1346.
[3] Shen J Z, Wang S S, Sun L T, et al. Dynamic changes in metabolic and lipidomic profiles of tea plants during drought stress and re-watering [J]. Frontiers in Plant Science, 2022, 13: 978531. doi: 10.3389/fpls.2022.978531.
[4] 黃文鏡, 楊樹華, 葛紅, 等. AMF對觀賞植物生長發(fā)育影響的研究進展[J]. 中國農學通報, 2023, 39(7): 55-63.
Huang W J, Yang S H, Ge H, et al. Research of AMF on the growth and development of ornamental plants [J]. Chinese Agricultural Science Bulletin, 2023, 39(7): 55-63.
[5] Bagheri V, Shamshiri M H, Alaei H, et al. The role of inoculum identity for growth, photosynthesis, and chlorophyll fluorescence of zinnia [J]. Bhotosynthetica, 2019, 57(2): 409-419.
[6] Ye Q H, Wang H, Li H. Arbuscular mycorrhizal fungi enhance drought stress tolerance by regulating osmotic balance [J]. Australian Journal of Grape and Wine Research, 2023, 2023: 1-13.
[7] Abdurrahim Y, Ertan Y, Hilal Y, et al. Use of arbuscular mycorrhizal fungi for boosting antioxidant enzyme metabolism and mitigating saline stress in sweet basil (Ocimum basilicum L.) [J]. Sustainability, 2023, 15(7): 5982-5996.
[8] Bahadur A L I, Batool A S F A, Nasir F A H A D, et al. Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants [J]. International Journal of Molecular Sciences, 2019, 20(17): 4199-4217.
[9] Liang S M, Li Q S, Liu M Y, et al. Mycorrhizal effects on growth and expressions of stress-responsive genes (aquaporins and SOSs) of tomato under salt stress [J]. Journal of Fungi, 2022, 8(12): 1305-1315.
[10] Yan Q X, Li X Y, Xiao X F, et al. Arbuscular mycorrhizal fungi improve the growth and drought tolerance of Cinnamomum migao by enhancing physio-biochemical responses [J]. Ecology and Evolution, 2022, 12(7): e9091. doi: 10.1002/ece3.9091.
[11] Singh S, Pandey A, Chaurasia B, et al. Diversity of arbuscular mycorrhizal fungi associated with the rhizosphere of tea growing in ‘natural’ and ‘cultivated’ ecosites [J]. Biology and Fertility of Soils, 2008, 44: 491-500.
[12] 盧東升, 吳小芹. 豫南茶園VA菌根真菌種類研究[J]. 南京林業(yè)大學學報(自然科學版), 2005, 29(3): 33-36.
Lu D S, Wu X Q. Species of VAM fungi around tea roots in the southern area of Henan province [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2005, 29(3): 33-36.
[13] 王玉娟, 高秀兵, 吳強盛, 等. 不同水分條件下AM真菌對福鼎大白茶生長和茶葉品質的影響[J]. 茶葉科學, 2020, 40(5): 588-596.
Wang Y J, Gao X B, Wu Q S, et al. Influences of arbuscular mycorrhizal fungi on plant growth and tea quality of Fuding Dabaicha seedlings under different water conditions [J]. Journal of Tea Science, 2020, 40(5): 588-596.
[14] 許平輝, 王飛權, 齊玉崗, 等. 叢枝菌根真菌對茶樹抗旱性的影響[J]. 西北農業(yè)學報,2017, 26(7): 1033-1040.
Xu P H, Wang F Q, Qi Y G, et al. Effect of arbuscular mycorrhiza fungi on drought resistance in tea plant (Camellia sinensis) [J]. Acta Agriculturae Boreali-occidentalis Sinica, 2017, 26(7): 1033-1040.
[15] 王學奎. 植物生理生化實驗原理和技術[M]. 2版. 北京: 高等教育出版社, 2006.
Wang X K. Experimental principles and techniques of plant physiology and biochemistry [M]. 2nd ed. Beijing: Higher Education Press, 2006.
[16] Phillips J M, Hayman D S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection [J]. Transactions of the British Mycological Society, 1970, 55: 158-161.
[17] Bethlenfalvay G J, Ames R N. Comparison of two methods for quantifying extraradical mycelium of vesicular-arbuscular mycorrhizal fungi [J]. Soil Science Society of America Journal, 1987, 51: 834-837.
[18] Bajji M, Lutts S, Kinet J M. Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum Desf.) cultivars performing differently in arid conditions [J]. Plant Science, 2001, 160(4): 669-681.
[19] 孫琪璐. 復水方式對干旱脅迫下茶樹的影響及CsSnRK2基因家族的克隆與表達[D]. 合肥: 安徽農業(yè)大學, 2019.
Sun Q L. Effect of rehydration method on tea plant under drought stress and cloning and expression of CsSnRK2 gene family [D]. Hefei: Anhui Agricultural University, 2019.
[20] 楊妮, 萬綺雯, 李逸民, 等. 外源亞精胺對鹽脅迫下茶樹光合特性及關鍵酶基因表達的影響[J]. 園藝學報, 2022, 49(2): 378-394.
Yang N, Wan Q W, Li Y M, et al. Effects of exogenous spermidine on photosynthetic characteristics and gene expression of key enzymes under salt stress in tea plant [J]. Acta Horticulturae Sinica, 2022, 49(2): 378-394.
[21] Liu C Y, Hao Y, Wu X L, et al. Arbuscular mycorrhizal fungi improve drought tolerance of tea plants via modulating root architecture and hormones [J]. Plant Growth Regulation, 2024, 102(1): 13-22.
[22] Zou Y N, Zhang F, Srivastava A K, et al. Arbuscular mycorrhizal fungi regulate polyamine homeostasis in roots of trifoliate orange for improved adaptation to soil moisture deficit stress [J]. Frontiers Plant Science, 2021, 11: 600792. doi: 10.3389/fpls.2020.600792.
[23] Liu C Y, Wang Y J, Wu Q S, et al. Arbuscular mycorrhizal fungi improve the antioxidant capacity of tea (Camellia sinensis) seedlings under drought stress [J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2020, 48(4): 1993-2005.
[24] Ortas I, Rafique M, ?eki? F ?. Do mycorrhizal fungi enable plants to cope with abiotic stresses by overcoming the detrimental effects of salinity and improving drought tolerance? [J]. Symbiotic Soil Microorganisms: Biology and Applications, 2021: 391-428.
[25] 李雪. 叢枝菌根真菌對干旱脅迫下柑橘幼苗生長及抗旱性的影響[D]. 重慶: 西南大學, 2022.
Li X. Effects of arbuscular mycorrhizal fungi on growth and drought resistance of citrus seedlings under water stress [D]. Chongqing: Southwest University, 2022.
[26] 蒲子天, 張林, 張弛, 等. 叢枝菌根真菌與植物共生影響植物水分狀態(tài)的研究進展[J]. 土壤, 2022, 54(5): 882-889.
Pu Z T, Zhang L, Zhang C, et al. Research progress of arbuscular mycorrhizal fungi and plant symbiosis affecting plant water regime [J]. Soils, 2022, 54(5): 882-889.
[27] Guo X N, Hao Y, Wu X L, et al. Exogenous easily extractable glomalin-related soil protein stimulates plant growth by regulating tonoplast intrinsic protein expression in lemon [J]. Plants, 2023, 12(16): 2955-2967.
[28] Liu C Y, Guo X N, Dai F J, et al. Mycorrhizal symbiosis enhances P uptake and indole-3-acetic acid accumulation to improve root morphology in different citrus genotypes [J]. Horticulture, 2024, 10(4): 339-415.
[29] Liu C Y, Zhang F, Zhang D J, et al. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress [J]. Scientific Reports, 2018, 8(1): 1978-1987.
[30] 王浩, 孫麗英. 植物激素調控叢枝菌根發(fā)育的作用機制研究進展[J]. 微生物學通報, 2022, 49(10): 4448-4466.
Wang H, Sun L Y. Mechanisms of phytohormones in regulating arbuscular mycorrhiza development [J]. Microbiology China, 2022, 49(10): 4448-4466.
[31] 方必君, 卓定龍, 劉曉洲, 等. 干旱脅迫及復水對野牡丹光合和葉綠素熒光參數的影響[J]. 熱帶農業(yè)科學, 2023, 43(2): 44-49.
Fang B J, Zhuo D L, Liu X Z, et al. Effects of drought stress and rehydration on photosynthetic and chlorophyll fluorescence parameters of Melastoma candidum D. Don [J]. Chinese Journal of Tropical Agriculture, 2023, 43(2): 44-49.
[32] 馬坤, 王彥淇, 楊建軍, 等. 不同干旱脅迫條件下叢枝菌根真菌對木棉葉綠素熒光參數的影響[J]. 植物資源與環(huán)境學報, 2017, 26(3): 35-43.
Ma K, Wang Y Q, Yang J J, et al. Effect of arbuscular mycorrhizal fungi on chlorophyll fluorescence parameters of Bombax ceiba under different drought stress conditions [J]. Journal of Plant Resources and Environment, 2017, 26(3): 35-43.
[33] 吳學蕤, 趙慶霞, 蔡銀美, 等. 干旱-復水對構樹葉片水勢和氣孔開閉的影響[J]. 草地學報, 2023, 31(3): 769-776.
Wu X R, Zhao Q X, Cai Y M, et al. Effects of drought-rewatering on leaf water potential and stomatal opening and closing of Broussonetia papyrifera [J]. Acta Agrestia Sinica, 2023, 31(3): 769-776.
[34] 劉婷, 唐明. 叢枝菌根真菌對楊樹生長、氣孔和木質部微觀結構的影響[J]. 植物生態(tài)學報, 2014, 38(9): 1001-1007.
Liu T, Tang M. Effects of arbuscular mycorrhizal fungi on growth and anatomical properties of stomata and xylem in poplars [J]. Chinese Journal of Plant Ecology, 2014, 38(9): 1001-1007.
[35] 楊海莉. 小花堿茅對滲透脅迫與等滲透勢鹽脅迫的生理響應[D]. 蘭州: 蘭州大學, 2019.
Yang H L. Physiological response of Puccinellia tenuiflora to osmotic and isotonic salt stress [D]. Lanzhou: Lanzhou University, 2019.
[36] 李津津, 趙書崗, 安秀紅, 等. 干旱脅迫下核桃生理適應性及抗性指標篩選[J]. 中國果樹, 2023, 84(3): 72-78.
Li J J, Zhao S G, An X H, et al. Physiological adaptation of walnut under drought stress and screening of its resistance indexes [J]. China Fruits, 2023, 84(3): 72-78.
[37] 張菲, 鄒英寧, 吳強盛. AM真菌摩西管柄囊霉對干旱脅迫下枳抗氧化酶基因表達的影響[J]. 菌物學報, 2019, 38(11): 2043-2050.
Zhang F, Zou Y N, Wu Q S. Effects of Funneliformis mosseae on the expression of antioxidant enzyme genes in trifoliate orange exposed to drought stress [J]. Mycosystema, 2019, 38(11): 2043-2050.
