紅景天苷對犬細(xì)小病毒體外復(fù)制的抑制效應(yīng)分析
2024-12-31 00:00:00張美雯王成龍劉玉貞趙育桐朱記平李毅
畜牧獸醫(yī)學(xué)報 2024年12期
關(guān)鍵詞:細(xì)胞凋亡




摘" 要: 旨在分析紅景天苷(salidroside,SAL)對犬細(xì)小病毒(canine parvovirus,CPV)的抑制作用及其機制。對病毒感染過程的3個節(jié)點(吸附、侵入、復(fù)制)分別添加SAL進(jìn)行孵育,檢測細(xì)胞中病毒滴度,評價SAL對病毒的抑制作用;TUNEL (Terminal deoxynucleotidyl Transferase dUTP Nick End Labeling)試驗分析細(xì)胞凋亡;實時熒光定量PCR和Western blot分別檢測細(xì)胞凋亡相關(guān)蛋白和炎癥因子的mRNA或蛋白表達(dá),分析和初步驗證SAL抑制病毒復(fù)制的機制。結(jié)果顯示,SAL可顯著抑制CPV復(fù)制,但對CPV的吸附及侵入過程沒有顯著影響。SAL可抑制CPV誘導(dǎo)的細(xì)胞凋亡,且顯著抑制了caspase 8及tBID的蛋白表達(dá)。進(jìn)一步研究發(fā)現(xiàn),CPV可誘導(dǎo)IL-1β及相關(guān)炎癥因子上調(diào)表達(dá),而SAL孵育病毒感染的細(xì)胞后部分炎癥因子下調(diào)表達(dá)。Caspase 1、NLRP3的激活與IL-1β密切相關(guān),病毒感染細(xì)胞后添加SAL進(jìn)行孵育可抑制caspase 1的激活,而對NLRP3無顯著作用。應(yīng)用siRNA-caspase 8下調(diào)細(xì)胞中caspase 8的表達(dá)再孵育病毒,結(jié)果顯示,細(xì)胞中IL-1β表達(dá)被抑制,與SAL的作用效果一致,表明SAL主要通過抑制caspase 8信號通路活化從而抑制炎癥因子的表達(dá)。綜上所述,SAL可有效抑制CPV的復(fù)制,其通過調(diào)控caspase 8信號通路抑制了CPV誘導(dǎo)的細(xì)胞凋亡,且抑制了部分炎癥因子的表達(dá)。本研究為有效治療CPV提供了新的途徑和方法。
關(guān)鍵詞: 犬細(xì)小病毒(CPV);紅景天苷;細(xì)胞凋亡;細(xì)胞因子
中圖分類號: S852.659.2
文獻(xiàn)標(biāo)志碼:A
文章編號:0366-6964(2024)12-5738-13
doi: 10.11843/j.issn.0366-6964.2024.12.036
開放科學(xué)(資源服務(wù))標(biāo)識碼(OSID):
收稿日期:2023-12-06
基金項目:
作者簡介:張美雯(1997-),女,湖北武漢人,碩士,主要從事病毒致病機理及抗病毒研究,E-mail: zmw5868782222@163.com;Tel:027-89648113
*通信作者:李" 毅,主要從事病毒致病機理研究及抗病毒藥物研發(fā),E-mail: liyi@whsw.edu.cn;朱記平,主要從事病毒致病機理研究及抗病毒藥物研發(fā),E-mail: jp_zhu732@126.com
Analysis of the Inhibitory Effect of Salidroside on Canine Parvovirus Replication in vitro
ZHANG" Meiwen1, 2, WANG" Chenglong1, LIU" Yuzhen1, ZHAO" Yutong1, ZHU" Jiping1*, LI" Yi1*
(1.Wuhan University of Bioengineering School of Life Sciences and Technology, Wuhan 430415," China;
2.Jingshan People’s Hospital, Jingshan 431800, China)
Abstract:nbsp; This study analyzes the inhibitory effect and its mechanism of salidroside (SAL) on canine parvovirus (CPV). In vitro, drug effects were performed on three stages of virus infection (adsorption, invasion, and replication). Virus titers were detected in cells to access the inhibitory effect of SAL on the virus were evaluated. TUNEL (Terminal deoxynucleotidyl Transferase dUTP Nick End Labeling) assay was used to analyze apoptosis. Quantitative Real-time PCR and Western blot were used to detect the mRNA or protein expression of apoptosis-related proteins and inflammatory factors, respectively, aiming to analyze and preliminarily verify the mechanism by which SAL inhibits virus replication. The results showed that SAL significantly inhibited CPV replication, but had no significant effect on the adsorption and invasion processes of CPV. SAL could inhibit apoptosis induced by CPV and significantly inhibit the protein expression of caspase 8 and tBID. Further research revealed that CPV could induce the upregulation of IL-1β and inflammatory factors, while SAL downregulated the expression of some inflammatory factors in virus-infected cell. The activation of caspase 1 and NLRP3 was closely related to IL-1β. After viral infection of cells, incubation with SAL could inhibit the activation of caspase 1, but had no significant effect on NLRP3. When virus was incubated in cells with down-regulated expression of caspase 8 by siRNA-caspase 8, the expression of IL-1β in cells was inhibited, consistent with the effect of SAL, indicating that SAL mainly inhibits the expression of inflammatory factors by inhibiting the activation of the caspase 8 signaling pathway. In summary, this study shows that SAL can effectively inhibit the replication of CPV. SAL inhibits CPV-induced apoptosis by regulating the caspase 8 signaling pathway and inhibits the expression of some inflammatory factors. This study provides a new approach and method for the effective treatment of CPV.
Key words: canine parvovirus(CPV); salidroside; apoptosis; cytokines
*Corresponding authors: LI Yi,E-mail: liyi@whsw.edu.cn; ZHU Jiping,E-mail: jp_zhu732@126.com
犬細(xì)小病毒(canine parvovirus,CPV)是導(dǎo)致幼犬發(fā)病和死亡最常見的原因之一。CPV屬于細(xì)小病毒科原細(xì)小病毒屬,是一種小的、無囊膜的單鏈DNA病毒[1]。病毒粒子呈正20面體對稱結(jié)構(gòu),由60個結(jié)構(gòu)蛋白裝配而成,直徑為21~24 nm[2]。CPV可感染快速分裂的胃腸道細(xì)胞、骨髓、淋巴組織和心肌細(xì)胞,導(dǎo)致發(fā)熱(40℃以上)、精神沉郁、不食、嘔吐、腹瀉、排出惡臭的糞便,并伴有白細(xì)胞大量減少,引發(fā)急性出血性腸炎和急性心肌炎[3]。目前,犬細(xì)小病毒的防治主要為疫苗預(yù)防和藥物治療。滅活疫苗、弱毒疫苗、基因工程疫苗和CPV多肽疫苗等可用于預(yù)防犬細(xì)小病毒。最為常用的是CPV弱毒疫苗,但弱毒株對一些極易感染的動物存在一定的危險性,毒力易返強,其免疫效果容易受到多種因素的影響[4],且運輸和保存要求較為嚴(yán)格。對發(fā)病犬治療以對癥治療為主,通過靜脈輸液補充間質(zhì)液體丟失,并保持水化,同時補充白蛋白、服用止吐藥、服用抗生素進(jìn)行治療等[5]。
中藥對病毒感染的治療被廣泛關(guān)注,其具有顯著的療效且一般都具有較廣泛的抗病毒作用。紅景天又名羅森根,屬于景天科的多年生草本植物,在亞洲和歐洲的傳統(tǒng)藥物中被廣泛用作補藥、抗抑郁和抗炎藥物[6]。紅景天苷(salidroside,SAL)是紅景天屬植物中普遍存在的一類天然酚的次生代謝產(chǎn)物[7]。近年來,越來越多的研究表明,紅景天苷具有多種藥理活性,包括抗氧化、抗應(yīng)激、抗菌、免疫調(diào)節(jié)、血管調(diào)節(jié)、抗腫瘤、抗病毒、增強免疫、抗糖尿病、抗骨質(zhì)疏松等,是一種毒性低、副作用少的天然藥物[8]。SAL可通過阻斷NF-κB和ERK/MAPKS的激活抑制促炎細(xì)胞因子的產(chǎn)生[9]。SAL也可通過SIRT1/NF-κB途徑減少炎癥因子的表達(dá),如減少TNF-α和IL-1β的表達(dá)從而顯著改善D-半乳糖誘導(dǎo)的記憶障礙[10]。SAL可通過調(diào)節(jié)IFN-γ、IL-10、TNF-α和IL-2基因的表達(dá)顯著抑制柯薩奇病毒B3對小鼠心肌細(xì)胞的作用,說明SAL有可能成為防治病毒性心肌炎的有效藥物[11]。SAL可在體外抗EV71病毒,抑制EV71的穿入和吸附,對EV71的復(fù)制有較好的抑制作用且對細(xì)胞毒性較小[12]。SAL還可作為免疫佐劑調(diào)節(jié)小鼠的體液免疫和細(xì)胞免疫。SAL與卵清蛋白聯(lián)合使用可誘導(dǎo)小鼠脾細(xì)胞增殖,從而增強疫苗的免疫效果[13]。
當(dāng)前對SAL抗犬細(xì)小病毒的研究較少,本研究旨在分析SAL的體外抑制CPV復(fù)制作用,并初步探討其通過調(diào)節(jié)細(xì)胞凋亡以及天然免疫從而抑制病毒感染的機制,為臨床治療犬細(xì)小病毒病提供理論依據(jù)。
1" 材料與方法
1.1" 實驗細(xì)胞、病毒株和菌株
犬腎細(xì)胞(MDCK)由華中農(nóng)業(yè)大學(xué)農(nóng)業(yè)微生物學(xué)國家重點實驗室曹罡老師惠贈;犬細(xì)小病毒(CPV)由武漢生物工程學(xué)院生命科學(xué)與技術(shù)學(xué)院徐鵬老師惠贈。
DMEM培養(yǎng)基、胰酶-EDTA細(xì)胞消化液、轉(zhuǎn)染試劑PEI Max購自源培生物(中國);Nobimpex胎牛血清、無血清細(xì)胞凍存液購自武漢羅德生物(中國);紅景天苷購自源葉生物(中國),分析標(biāo)準(zhǔn)品,純度≥98%(液相色譜法);BCA蛋白濃度測定試劑盒、TUNEL試劑盒購自碧云天(中國);CCK-8、總RNA提取試劑盒、反轉(zhuǎn)錄試劑盒和qPCR Master Mix均購自南京諾唯贊(中國);IL-1β Rabbit pAb、IL-18 beta Rabbit pAb、caspase 8 Mouse mAb、caspase 1 Rabbit mAb、caspase 3 Polyclonal Antibody、caspase 9 Polyclonal Antibody、BID Antibody、NLRP3 Rabbit Polyclonal Antibody、GAPDH Polyclonal Antibody、Phospho-NF-κB、p65 Rabbit mAb購自正能生物(中國);熒光標(biāo)記二抗、HRP標(biāo)記二抗購自北京全式金生物(中國);抗CPV鼠源單克隆抗體購自山東綠都(中國)。
1.2" 紅景天苷最大無毒劑量的測定
稀釋SAL至終濃度分別為10、15、20、25、30、35、40、45、50、55、60 mmol·L-1。將各濃度SAL孵育細(xì)胞,以未加藥物組作為陰性對照,以無細(xì)胞的DMEM作為空白對照,每個濃度6復(fù)孔。在37 ℃,50 mmol·L-1 CO2條件下孵育48 h后,向每孔內(nèi)加入10 μL CCK-8試劑,培養(yǎng)3 h。在450 nm波長處測定吸光度。細(xì)胞存活率大于90%的藥物濃度即為該藥物最大無毒劑量。
1.3" CPV在MDCK細(xì)胞中的增殖動態(tài)
0.1 MOI CPV孵育MDCK細(xì)胞1 h后棄上清更換為維持液。在感染后6、12、24、36、48、60、72 h分別收集細(xì)胞懸液,反復(fù)凍融3次后,12 000 r·min-1,離心15 min,取上清通過TCID50測定病毒滴度,獨立重復(fù)3次。
1.4" 紅景天苷抑制CPV復(fù)制
0.1 MOI的CPV孵育MDCK細(xì)胞1 h后,用PBS清洗細(xì)胞3次,更換培養(yǎng)基為含SAL(終濃度分別為30、35、40 mmol·L-1)的細(xì)胞維持液,同時設(shè)置未加藥物的空白對照組,在感染后12、24、36 h分別收集樣本,通過TCID50測定病毒滴度。
1.5" 間接免疫熒光技術(shù)檢測細(xì)胞內(nèi)病毒
0.1 MOI CPV孵育細(xì)胞1 h后更換培養(yǎng)基為分別含40 mmol·L-1 SAL、40 mmol·L-1 SAL加4 μmol·L-1 TAT-scFV、4 μmol·L-1 TAT-scFV的細(xì)胞維持液,同時設(shè)置僅孵育病毒的對照組,24 h后PBS清洗細(xì)胞3次。預(yù)冷的40 mL·L-1多聚甲醛固定細(xì)胞30 min,PBS清洗細(xì)胞3次。5 mL·L-1 Triton X-100室溫穿透5 min,PBS清洗細(xì)胞3次。加入50 mL·L-1 BSA封閉過夜。特異性CPV抗體(1∶300)4 ℃孵育過夜,PBS清洗細(xì)胞3次。FITC標(biāo)記的鼠二抗(1∶3 000)室溫避光孵育1 h,PBS清洗細(xì)胞3次。避光孵育DAPI染色液(1∶5 000)15 min,PBS清洗細(xì)胞3次。樣品封片后通過熒光顯微鏡觀察。
1.6" 紅景天苷抑制CPV感染階段
分析不同加藥方式下藥物對病毒感染的抑制作用。A組:病毒感染細(xì)胞后孵育藥物,0.1 MOI CPV孵育細(xì)胞1 h后棄上清,40 mmol·L-1 SAL孵育細(xì)胞2 h后更換為維持液;B組:藥物預(yù)處理,40 mmol·L-1 SAL孵育細(xì)胞2 h后,加入0.1 MOI CPV,1 h后棄上清更換為維持液;C組:同時孵育藥物和病毒,40 mmol·L-1 SAL與0.1 MOI CPV共同孵育細(xì)胞,2 h后更換為維持液培養(yǎng)。分別在感染后12、24、36 h收集樣本測定病毒滴度。
為分析紅景天苷對CPV吸附細(xì)胞的影響,將MDCK細(xì)胞置于4 ℃預(yù)冷1 h后加入0.1 MOI CPV和40 mmol·L-1 SAL,4 ℃下孵育1 h,棄上清,用冷PBS洗3次,加入維持液繼續(xù)培養(yǎng),12、24、36 h后分別收集樣本測定病毒滴度。
為分析紅景天苷對CPV侵入細(xì)胞的影響,細(xì)胞置于4 ℃預(yù)冷1 h,之后接種10 MOI CPV,于4 ℃孵育1 h,以確保所有細(xì)胞都被感染并防止多輪感染。之后用冷PBS洗3次,加入含40 mmol·L-1 SAL的維持液于37 ℃培養(yǎng),以允許CPV內(nèi)化,2 h后更換為不含SAL的維持液,孵育8 h,收集樣本測定病毒滴度。
1.7" TUNEL試驗
0.1 MOI CPV孵育細(xì)胞1 h后更換培養(yǎng)基為分別含40 mmol·L-1 SAL、40 mmol·L-1 SAL與4 μmol·L-1 TAT-scFV、4 μmol·L-1 TAT-scFV的細(xì)胞維持液,同時設(shè)置僅孵育病毒的對照組,24 h后PBS洗滌細(xì)胞3次。預(yù)冷的40 mL·L-1多聚甲醛固定細(xì)胞30 min,PBS洗滌細(xì)胞3次。1 mL·L-1 Triton X-100孵育5 min,PBS清洗細(xì)胞3次。配制TUNEL檢測液:TdT酶10 μL、熒光標(biāo)記液240 μL、TUNEL檢測液250 μL。向細(xì)胞上加入50 μL TUNEL檢測液,室溫避光孵育1 h,PBS清洗細(xì)胞3次。樣品封片后通過熒光顯微鏡觀察。
1.8" 實時熒光定量PCR
設(shè)計引物,序列信息見表1,由武漢奧科鼎盛生物有限公司合成。0.1 MOI CPV孵育MDCK細(xì)胞1 h后換成含有40 mmol·L-1 SAL的維持液,24 h后收取細(xì)胞。提取細(xì)胞總RNA反轉(zhuǎn)錄為cDNA進(jìn)行qRT-PCR測定。以GAPDH作為內(nèi)參,采用2-ΔΔCT法計算基因表達(dá)量。
1.9" Western blot檢測蛋白表達(dá)
向細(xì)胞上加普通型RIPA裂解液(含PMSF),冰上裂解45 min。刮下細(xì)胞,12 000 r·min-1離心10 min,收集上清即細(xì)胞總蛋白。BCA測蛋白濃度后,加入5×SDS Loading Buffer至蛋白樣品中沸水浴10 min,待冷卻后進(jìn)行SDS-PAGE電泳,轉(zhuǎn)膜封閉后分別加入一抗,4 ℃孵育過夜,TBST洗細(xì)胞5次;二抗孵育1.5 h,TBST洗細(xì)胞5次。將清洗干凈的PVDF膜置于化學(xué)發(fā)光儀中,按照說明書配置ECL顯影液顯影。觀察蛋白表達(dá)并拍照記錄。
1.10" siRNA干擾及病毒感染
犬caspase-8的siRNA由武漢奧科鼎盛生物有限公司合成,Negative control:UUCUCCGAACGUGUCACGUTT;siRNA-caspase 8 (16 534~16 554 bp):UCAAAGAAGUCCAGAUGAATT。根據(jù)PEI MAX說明書進(jìn)行細(xì)胞轉(zhuǎn)染,siRNA終濃度為20 nmol·L-1,轉(zhuǎn)染后24 h孵育0.1 MOI CPV,36 h后收集細(xì)胞提取總蛋白進(jìn)行Western blot檢測。
1.11" 統(tǒng)計學(xué)分析
利用軟件GraphPad Prism 5進(jìn)行統(tǒng)計分析、繪圖,且在圖中反映出試驗結(jié)果的標(biāo)準(zhǔn)差。顯著性分析方法為Student t檢驗,Plt;0.05時認(rèn)為差異具有統(tǒng)計學(xué)意義,當(dāng)Plt;0.05時標(biāo)記為*,當(dāng)Plt;0.01時標(biāo)記為**。
2" 結(jié)" 果
2.1" 紅景天苷抑制CPV復(fù)制
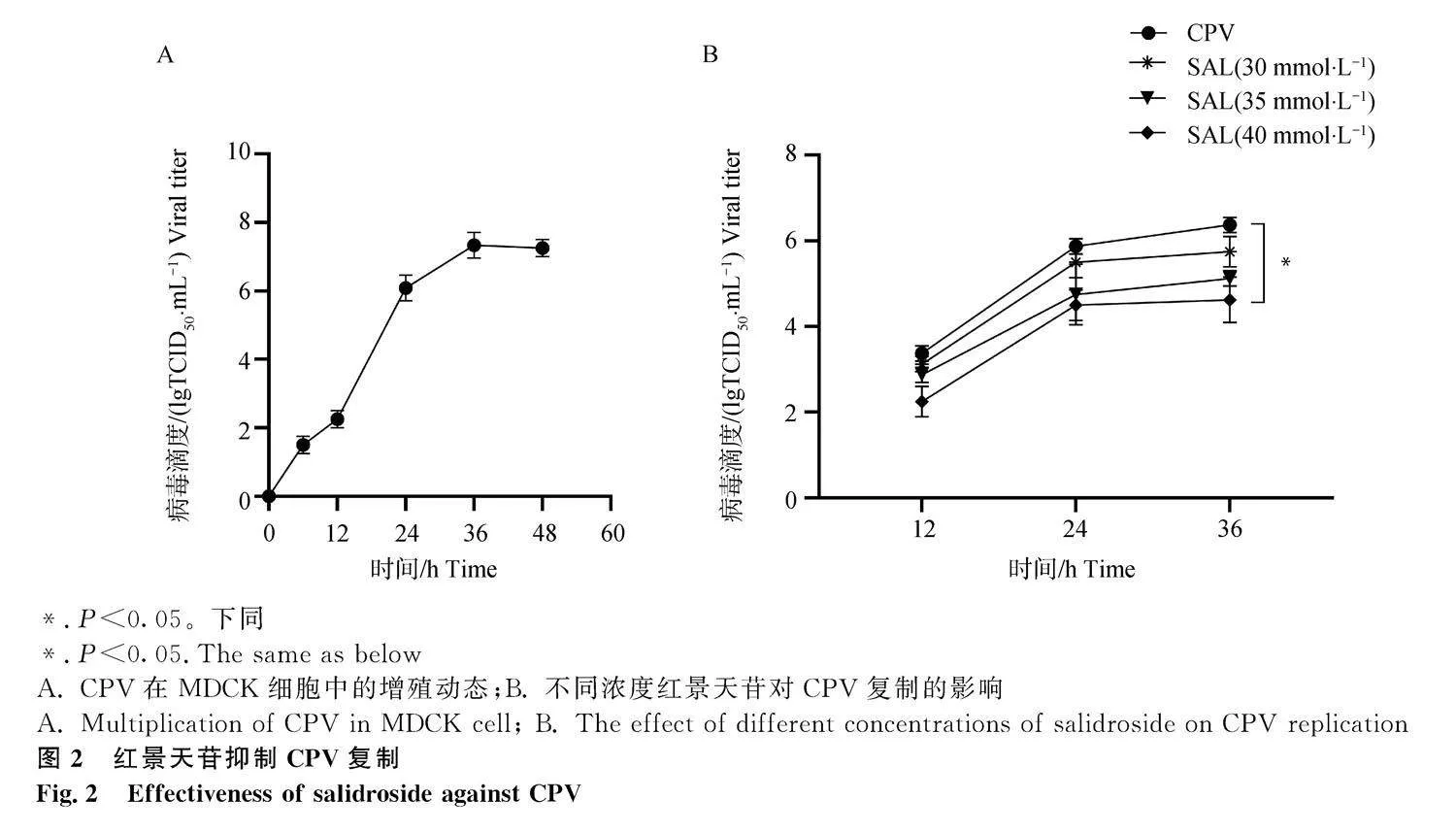
不同濃度的SAL孵育MDCK細(xì)胞,當(dāng)SAL的濃度低于45 mmol·L-1時,對MDCK細(xì)胞活性無明顯影響,當(dāng)藥物濃度大于或等于50 mmol·L-1時,細(xì)胞活性降低到85%以下(圖1)。因此,后續(xù)研究,SAL作用濃度小于或等于45 mmol·L-1。CPV感染MDCK細(xì)胞36 h后病毒滴度達(dá)到峰值(圖2A)。將0.1 MOI CPV孵育MDCK細(xì)胞1 h后,更換為含不同濃度SAL的細(xì)胞維持液,同時設(shè)置未加藥物的對照組。分別檢測感染后不同時間的病毒滴度。結(jié)果如圖2B所示,與對照組相比,SAL可有效抑制病毒復(fù)制,且成劑量依賴性。
2.2" 紅景天苷聯(lián)合單鏈抗體(TAT-scFV)顯著抑制CPV復(fù)制
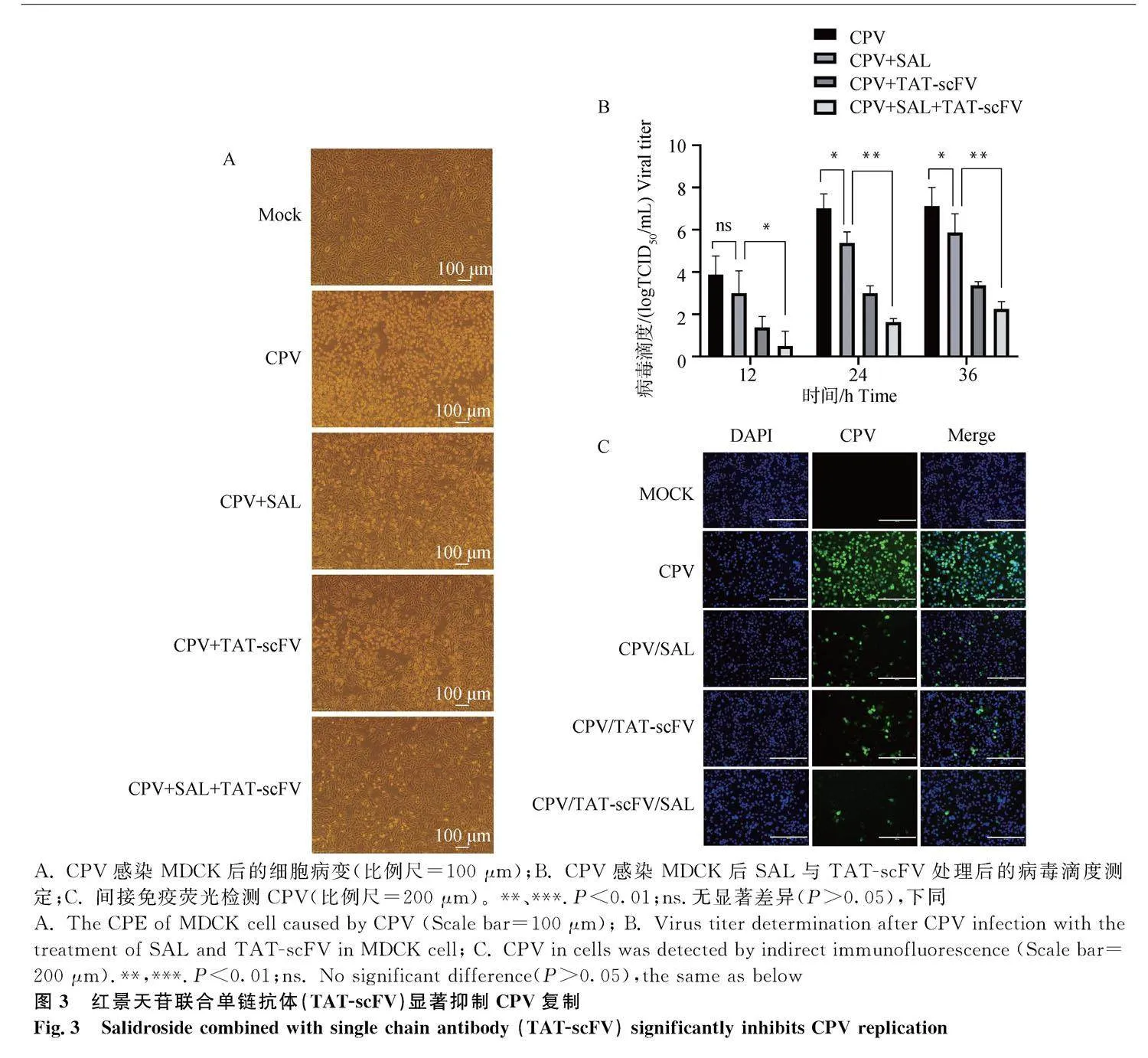
本實驗室前期制備了抗CPV的單鏈抗體(TAT-scFV),其可有效抑制CPV在細(xì)胞中的復(fù)制[14]。為檢測SAL聯(lián)合TAT-scFV的抑制病毒復(fù)
制作用,將CPV孵育MDCK細(xì)胞1 h后更換為含SAL或/和TAT-scFV維持液,24 h后觀察細(xì)胞病變。結(jié)果顯示(圖3A),與正常細(xì)胞相比,病毒感染細(xì)胞后,細(xì)胞大量病變;病毒感染細(xì)胞后單獨孵育SAL或TAT-scFV,細(xì)胞病變部分減少;病毒感染細(xì)胞后共同孵育SAL和TAT-scFV,細(xì)胞病變顯著減少。同時檢測子代病毒滴度,結(jié)果顯示(圖3B),相較于單獨孵育SAL或TAT-scFV細(xì)胞組,SAL和TAT-scFV共同孵育細(xì)胞組后病毒滴度顯著降低。間接免疫熒光技術(shù)檢測細(xì)胞中CPV,結(jié)果顯示(圖3C),SAL和TAT-scFV共同孵育細(xì)胞組,有熒光信號的細(xì)胞更少。綜上結(jié)果表明,SAL與TAT-scFV聯(lián)合用藥可更顯著抑制CPV復(fù)制。
2.3 "紅景天苷抗CPV感染階段
分別在病毒感染細(xì)胞后孵育SAL(圖4A)、SAL預(yù)處理細(xì)胞后孵育病毒(圖4B)、SAL和病毒同時孵育細(xì)胞(圖4C)后檢測病毒滴度。結(jié)果顯示,SAL預(yù)處理或同時孵育SAL和CPV組,病毒滴度無顯著差異,而病毒感染后孵育SAL,24 h后病毒滴度顯著下降,表明SAL在病毒感染后處理細(xì)胞的效果更好。
為進(jìn)一步分析SAL對CPV吸附過程的影響,將MDCK細(xì)胞置于4 ℃預(yù)冷1 h,加入CPV和SAL于4 ℃孵育1 h,棄上清,用預(yù)冷PBS沖洗3次以去除尚未吸附的病毒,加入維持液繼續(xù)培養(yǎng),分別測定12、24、36 h后病毒滴度。結(jié)果如圖4D所示,兩組的病毒滴度無顯著差異,表明SAL不能抑制CPV吸附宿主細(xì)胞表面受體,不影響病毒的吸附。
為分析SAL在病毒侵入過程中的影響,將MDCK細(xì)胞置于4 ℃預(yù)冷1 h后接種10 MOI CPV于4 ℃孵育1 h,以確保所有細(xì)胞都被病毒感染并防止多輪感染。之后SAL孵育細(xì)胞2 h后棄上清,添加維持液培養(yǎng)8 h后測定病毒滴度。結(jié)果如圖4E所示,與對照組相比,SAL處理細(xì)胞后,病毒滴度無顯著變化,表明SAL對CPV侵入沒有顯著影響。
2.4" 紅景天苷抑制病毒誘導(dǎo)的細(xì)胞凋亡
將CPV孵育細(xì)胞1 h后棄上清加入SAL或/和TAT-scFV孵育細(xì)胞。應(yīng)用TUNEL細(xì)胞凋亡檢測試劑盒分析細(xì)胞凋亡。結(jié)果顯示(圖5),CPV感染細(xì)胞后,可觀察到明顯的熒光信號,表明細(xì)胞凋亡顯著;病毒感染細(xì)胞后孵育SAL或TAT-scFV,熒光信號較弱,表明細(xì)胞凋亡受到一定抑制,而病毒感染細(xì)胞后共同孵育SAL和TAT-scFV,幾乎觀察不到熒光信號,表明細(xì)胞凋亡受到顯著抑制。
2.5" 紅景天苷抑制CPV活化的caspase-8信號通路
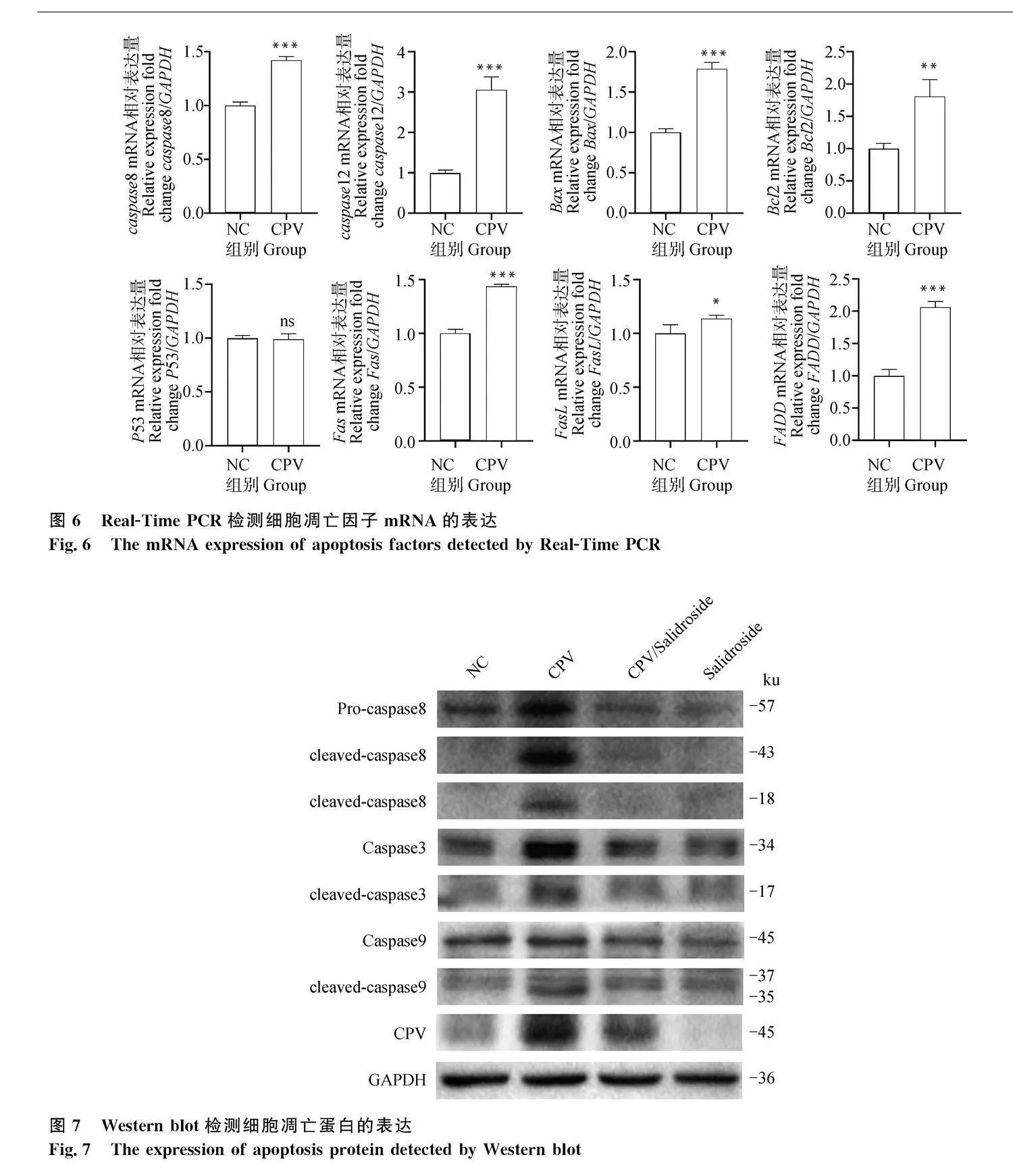
病毒感染MDCK細(xì)胞后,應(yīng)用qRT-PCR檢測凋亡通路相關(guān)蛋白的mRNA表達(dá),結(jié)果如圖6,病毒感染細(xì)胞后,caspase 8、caspase 12、Bcl2、Bax、FADD、FasL、Fas基因的mRNA表達(dá)顯著上調(diào),caspase 8、caspase 12、Bcl2、Bax mRNA表達(dá)上調(diào)表明CPV激活了MDCK細(xì)胞的內(nèi)源性線粒體凋亡途徑以及外源性死亡受體途徑;FADD、FasL、Fas mRNA表達(dá)上調(diào),F(xiàn)as為最典型的死亡受體,細(xì)胞凋亡后會激活FasL從而誘導(dǎo)Fas多聚體化,F(xiàn)as上的死亡結(jié)構(gòu)與FADD結(jié)合形成FADD-Fas復(fù)合物,而這種復(fù)合物則與caspase-8作用激活凋亡通路,推測MDCK細(xì)胞的凋亡可能與caspase 8相關(guān)聯(lián)。
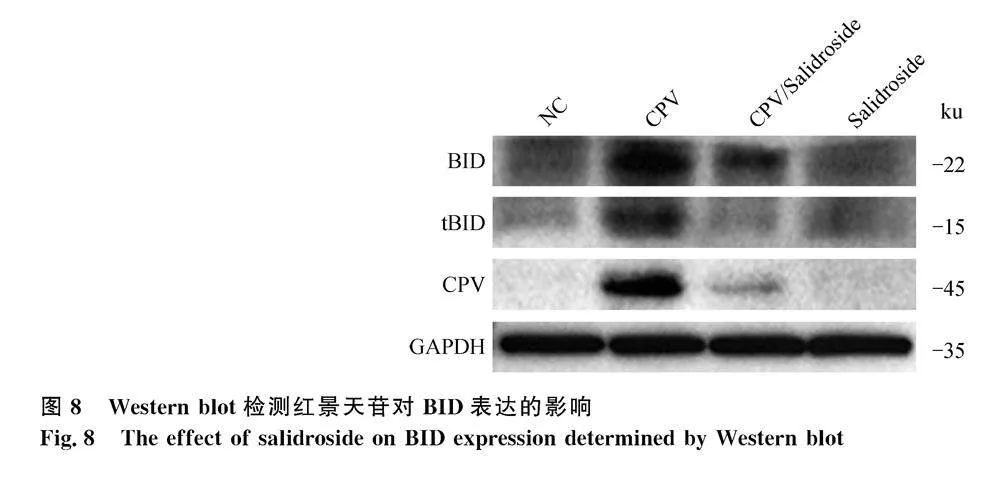
應(yīng)用Western blot檢測凋亡相關(guān)蛋白的表達(dá),如圖7所示,病毒感染后孵育SAL,細(xì)胞中caspase 8蛋白表達(dá)顯著下降,caspase 3、caspase 9蛋白表達(dá)在一定程度上也有所下降。Caspase 8激活主要通過兩條信號通路,其一為caspase 8直接激活caspase 3誘導(dǎo)凋亡;其二為通過間接作用激活線粒體途徑激活caspase 9 誘導(dǎo)凋亡。推測SAL可能主要通過抑制CPV活化的caspase 8信號通路抑制MDCK細(xì)胞凋亡。
Caspase 8激活后可通過激活BID蛋白切割,產(chǎn)生切割產(chǎn)物tBID,進(jìn)而通過激活線粒體途徑誘導(dǎo)凋亡。應(yīng)用Western blot檢測caspase 8通路下游關(guān)鍵蛋白tBID的活化,如圖8所示,CPV感染細(xì)胞后,tBID表達(dá)顯著提高;單獨孵育SAL和病毒感染細(xì)胞后孵育SAL均未檢測到明顯的tBID表達(dá)。結(jié)果表明病毒感染細(xì)胞后激活了死亡受體-FADD-caspase 8通路,而SAL可抑制CPV活化的caspase 8信號通路。
2.6" 紅景天苷對CPV誘導(dǎo)的天然免疫分子的影響
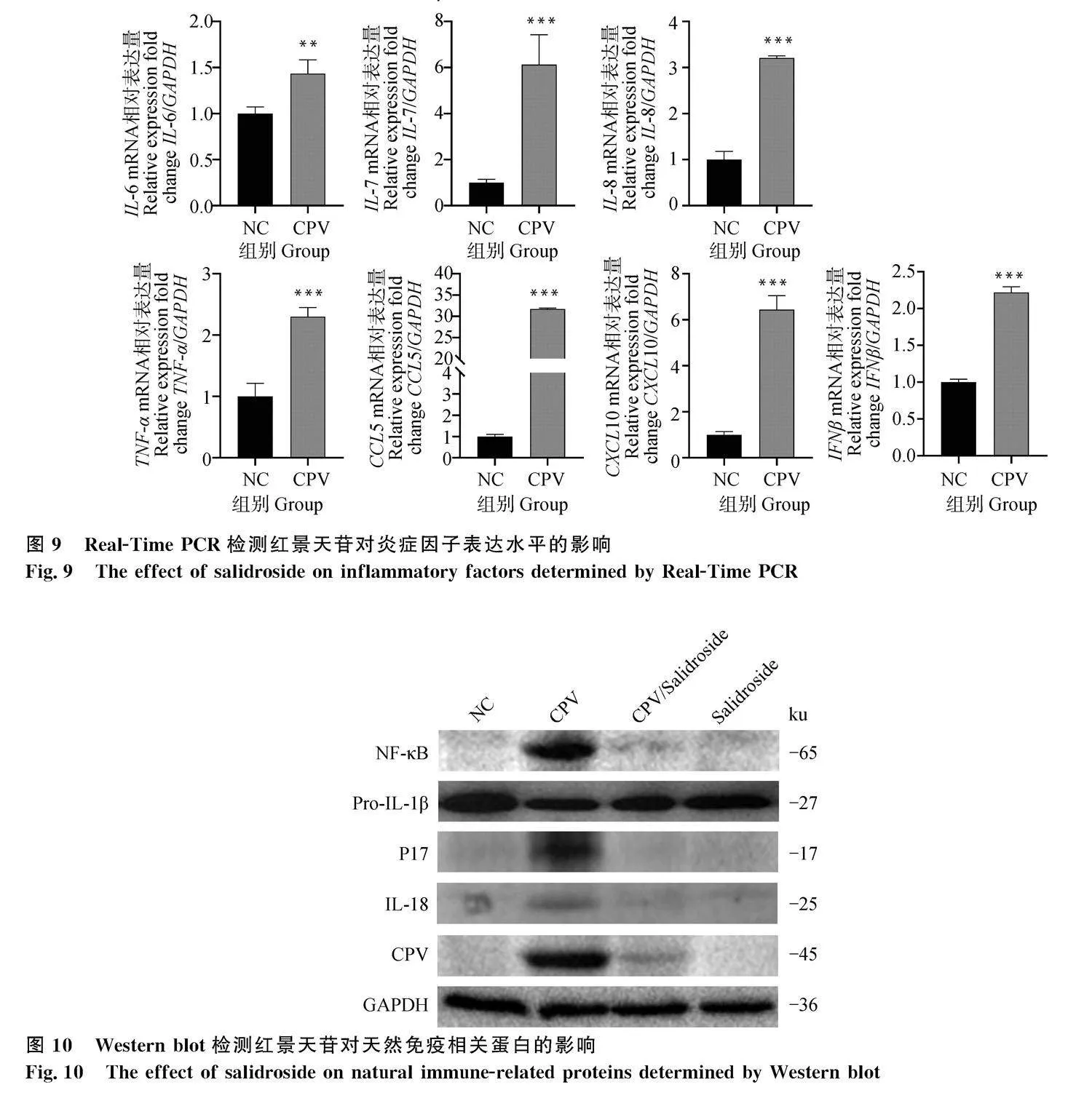
本研究檢測了天然免疫分子的mRNA表達(dá)。如圖9所示,病毒感染細(xì)胞后,細(xì)胞中IFNβ、IL-6、CXCL10、CCL5、IL-7、TNF-α、IL-8 mRNA表達(dá)顯著上調(diào)。如圖10所示,病毒感染細(xì)胞后孵育
SAL,NF-κB、IL-1β、IL-18蛋白表達(dá)顯著下降。結(jié)果表明,CPV感染細(xì)胞后,可激活I(lǐng)L-1β相關(guān)炎癥通路,SAL部分抑制了CPV誘導(dǎo)的天然免疫,SAL可能部分緩解了炎性風(fēng)暴引起的細(xì)胞損傷。
本研究檢測了caspase 1、NLRP3的蛋白表達(dá)。如圖11所示,病毒感染細(xì)胞后caspase 1組成型表達(dá)顯著上調(diào),但活化caspase 1的表達(dá)不明顯,而病毒感染細(xì)胞后孵育SAL,caspase 1的組成型表達(dá)和活化表達(dá)均不顯著;同時,NLRP3的表達(dá)在所有細(xì)胞樣品中無顯著差異。結(jié)果表明,IL-1β的分泌不是通過活化的caspase 1信號通路。
2.7" 敲低caspase-8對信號通路蛋白的影響
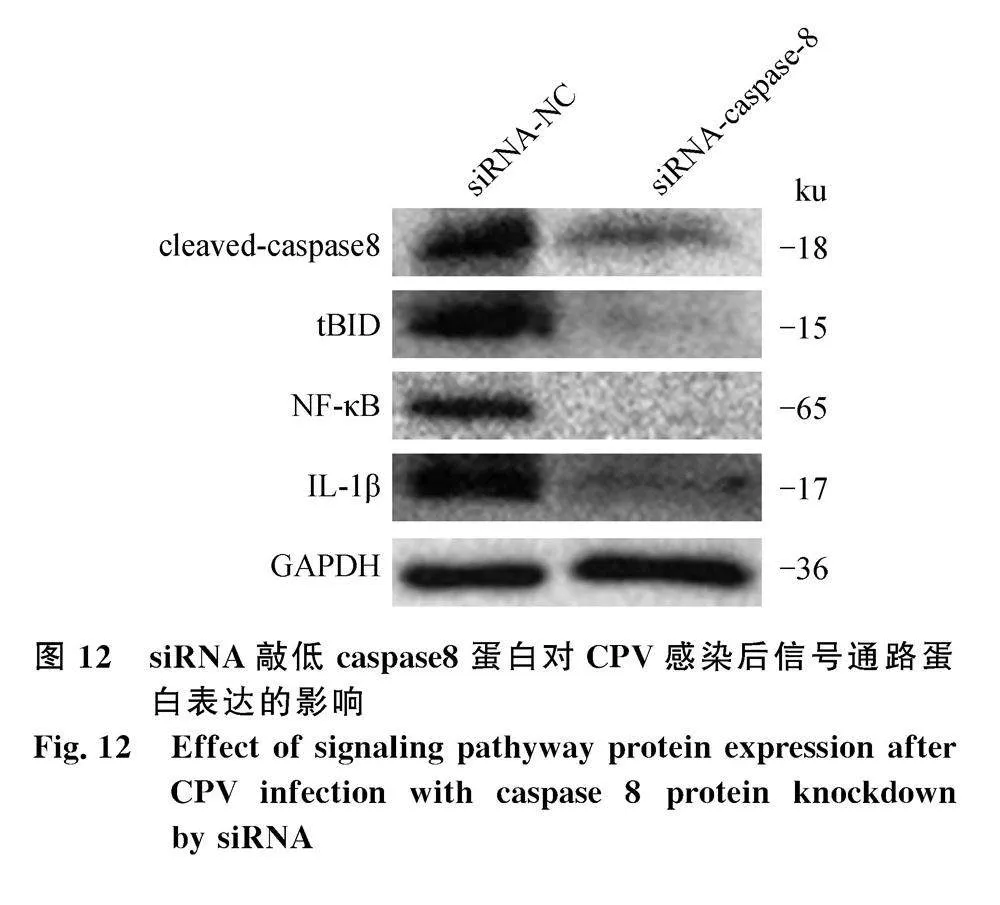
為進(jìn)一步確定caspase 8信號通路在CPV誘導(dǎo)的細(xì)胞凋亡和炎癥因子分泌過程中的作用,通過siRNA敲低細(xì)胞caspase 8的表達(dá),之后CPV感染細(xì)胞,結(jié)果如圖12所示,siRNA-caspase 8可顯著抑制CPV誘導(dǎo)的caspase 8的活化,同時,tBID、NF-κB、IL-1β的表達(dá)也顯著降低。該結(jié)果表明,CPV感染細(xì)胞后誘導(dǎo)caspase 8活化,活化的caspase 8一方面誘導(dǎo)細(xì)胞凋亡,另一方面誘導(dǎo)IL-1β及其信號通路活化,導(dǎo)致炎性因子大量分泌。SAL和siRNA-caspase 8均可有效抑制caspase 8及下游信號通路的活化。
3" 討" 論
CPV是引起犬急性出血性腸炎和心肌炎的主要病原之一,發(fā)病率高達(dá)100%,成年犬死亡率高達(dá)10%,幼犬死亡率高達(dá)91%。目前針對CPV感染的治療方案主要以對癥治療為主。SAL是紅景天的主要生物活性化合物,具有神經(jīng)保護(hù)、抗炎和抗病毒等作用。目前尚無關(guān)于SAL抗犬細(xì)小病毒的報道。
本研究通過檢測SAL對MDCK細(xì)胞的毒性,確定了SAL的藥物使用濃度,并發(fā)現(xiàn)SAL在細(xì)胞水平具有較好的抑制CPV復(fù)制作用。已有研究報道,SAL可通過抑制EV71的吸附和侵入抑制病毒復(fù)制[12]。SAL還可通過調(diào)節(jié)宿主天然免疫因子RIG-I、IRF-3、IRF-7、PKR、P-eIF2α和NF-κB抑制登革病毒[15]。本研究分析了SAL對CPV感染進(jìn)程的影響,結(jié)果顯示SAL對CPV的吸附和侵入階段均無較大的影響,表明SAL并不直接作用于CPV,然而,觀察到了SAL可抑制細(xì)胞凋亡,推測SAL可能通過其他途徑間接影響病毒復(fù)制。
細(xì)胞程序性死亡是維持細(xì)胞穩(wěn)定的狀態(tài)所需要的生理過程[16],也是病毒發(fā)病機理中的主要機制之一[17]。B19、鼠細(xì)小病毒和H-1細(xì)小病毒均能夠誘導(dǎo)相應(yīng)的細(xì)胞發(fā)生凋亡[18]。本研究結(jié)果顯示CPV可以促進(jìn)MDCK細(xì)胞凋亡,并刺激caspase 8、caspase 3、caspase 12、Bcl2、Bax、FADD、FasL、Fas上調(diào)表達(dá)。這與Doley等[19]研究結(jié)果一致。Caspase 8與caspase 12被激活,表明細(xì)胞凋亡存在死亡受體途徑和內(nèi)質(zhì)網(wǎng)途徑。Caspase 9的活化以及Bax/Bcl2比值的增加,表明了線粒體途徑也參與其凋亡過程。p53活化,則可能通過調(diào)節(jié)Bax與Bcl2的上調(diào)或下調(diào)而介導(dǎo)細(xì)胞凋亡[20]。Fas為最典型的死亡受體,細(xì)胞凋亡后會激活FasL從而誘導(dǎo)Fas多聚體化,F(xiàn)as上的死亡結(jié)構(gòu)(death domain,DD)與FADD結(jié)合形成FADD-Fas復(fù)合物,而這種復(fù)合物則與caspase 8作用激活凋亡通路[21]。Caspase 8激活后可通過激活BID蛋白切割,產(chǎn)生切割產(chǎn)物tBID,進(jìn)而通過激活線粒體途徑caspase 9誘導(dǎo)凋亡。2020年Li等[22]報道SARS-CoV-2感染細(xì)胞后可激活Pro-caspase 8,使caspase 8發(fā)生剪切,活化后caspase 8進(jìn)一步直接激活caspase 3,誘導(dǎo)細(xì)胞凋亡。本研究中SAL孵育CPV感染的細(xì)胞后,caspase 8的表達(dá)被顯著抑制,細(xì)胞凋亡的死亡受體途徑相關(guān)蛋白Fasl、Fas、FADD、tBID、caspase 9也顯著下調(diào),且同時觀察到caspase 3蛋白表達(dá)在一定程度上也有所下降,推測SAL通過抑制caspase 8的活化,進(jìn)而抑制了caspase 8和caspase 3相關(guān)死亡受體途徑誘導(dǎo)的細(xì)胞凋亡,但SAL對CPV激活的其它凋亡相關(guān)信號通路的作用機制有待進(jìn)一步研究。
研究表明caspase 8發(fā)揮多重調(diào)節(jié)作用。SARS-CoV-2感染觸發(fā)了細(xì)胞死亡的雙重途徑,Caspase 8依賴的炎癥反應(yīng)可能導(dǎo)致新冠肺炎患者的肺損傷[22]。目前認(rèn)為caspase 8還通過調(diào)節(jié)NF-κB的激活調(diào)節(jié)炎癥反應(yīng)[23],活化的caspase 8還通過調(diào)節(jié)IL-1β mRNA表達(dá)調(diào)節(jié)炎癥[24]。本研究通過qRT-PCR檢測了CPV感染MDCK細(xì)胞后炎癥因子mRNA的表達(dá),結(jié)果顯示IFNβ、IL-6、CXCL10、CCL5、IL-7、TNF-α、IL-8、IL-1β、NF-κB、IL-18表達(dá)均上調(diào),SAL處理病毒感染的細(xì)胞后可抑制相應(yīng)炎癥因子表達(dá),表明SAL可抑制病毒引起的炎癥反應(yīng)。紅景天通過調(diào)節(jié)免疫細(xì)胞分化、炎癥信號通路的激活和炎癥因子的分泌,發(fā)揮良好的免疫調(diào)節(jié)作用,減輕各種疾病的炎癥損害[25]。Song等[26]發(fā)現(xiàn)紅景天苷可減輕脂多糖誘導(dǎo)的大鼠肺組織病理損傷,抑制肺組織中TNFα和IL-6的分泌,可降低NR8383肺泡巨噬細(xì)胞產(chǎn)生的TNFα和IL-6,表明紅景天苷可減輕脂多糖誘導(dǎo)的急性肺損傷。
為進(jìn)一步分析CPV感染細(xì)胞后caspase 8是否為細(xì)胞凋亡和炎性反應(yīng)的關(guān)鍵蛋白,本研究中通過siRNA介導(dǎo)caspase 8蛋白下調(diào)表達(dá),結(jié)果顯示caspase 8蛋白下調(diào)后,病毒感染的細(xì)胞中,相關(guān)凋亡蛋白、炎性因子、病毒蛋白表達(dá)均下調(diào)。該結(jié)果表明CPV感染誘導(dǎo)caspase 8活化,活化的caspase 8可能是調(diào)控細(xì)胞炎癥因子表達(dá)的關(guān)鍵蛋白。SAL或siRNA-caspase 8對該過程起到一定抑制作用,同時caspase 8、IL-1β與NF-κB、IL-18具有相互調(diào)節(jié)的作用,但其具體調(diào)節(jié)機制還有待進(jìn)一步研究。
Caspase 8還可調(diào)節(jié)NLRP3。Gurung等[27]揭示了caspase 8和FADD在NLRP3炎癥小體復(fù)合物的啟動和激活中的作用。Man等[28]通過免疫共沉淀證實caspase 8與NLRP3可相互作用,且參與caspase 1的剪切與加工,此外,caspase 8可以直接特異性剪切caspase 1。通過siRNA敲低MLKL導(dǎo)致caspase 8負(fù)調(diào)控PIRK1-RIPK3-MLKL介導(dǎo)的NLRP3[29]。Caspase 1作為炎癥caspase的典型成員,參與IL-1β與IL-18的成熟過程[30-31]。本研究中,CPV誘導(dǎo)MDCK細(xì)胞產(chǎn)生的IL-1β依賴于caspase 8,不依賴于NLRP3炎癥體,SAL可抑制caspse 8誘導(dǎo)的IL-1β表達(dá),但對NLRP3的表達(dá)沒有較大作用,卻對caspase 1的活化有一定影響,其機理還需進(jìn)一步深入研究。
同時,本研究還初步分析了SAL與TAT-scFV聯(lián)合用藥的抑制病毒效果,結(jié)果表明SAL聯(lián)合TAT-scFV抑制病毒作用更顯著。SAL可能成為廣譜抑制病毒藥物候選,同時可能作為免疫應(yīng)答的佐劑,增強免疫應(yīng)答的作用效果。
4" 結(jié)" 論
CPV感染細(xì)胞后激活了死亡受體-FADD-caspase 8通路,caspase 8活化后誘導(dǎo)了IL-1β等一系列炎癥因子的激活,SAL通過抑制病毒誘導(dǎo)的細(xì)胞凋亡和炎癥反應(yīng)從而減輕病毒對細(xì)胞的損傷。SAL可能也抑制了CPV誘導(dǎo)的caspase 1活化。該研究為病毒感染性疾病的治療提供理論基礎(chǔ)和試驗依據(jù)。
參考文獻(xiàn)(References):
[1]" 趙" 航.犬細(xì)小病毒流行病學(xué)調(diào)查及其感染F81細(xì)胞的比較蛋白質(zhì)組學(xué)分析[D].北京:中國農(nóng)業(yè)科學(xué)院,2016.
ZHAO H.Canine parvovirus epidemiological investigation and comparative proteomics analysis of CPV infected F81 cells[D].Beijing:Chinese Academy of Agricultural Sciences,2016.(in Chinese)
[2]" 周宏專,蘇" 霞,徐福洲,等.犬細(xì)小病毒研究進(jìn)展[J].動物醫(yī)學(xué)進(jìn)展,2019,40(12):79-84.
ZHOU H Z,SU X,XU F Z,et al.Progress on canine parvovirus[J].Progress in Veterinary Medicine,2019,40(12):79-84.(in Chinese)
[3]" HOELZER K,PARRISH C R.The emergence of parvoviruses of carnivores[J].Vet Res,2010,41(6):39.
[4]" 胡" 楠,鄒翔宇,蔡錦順.犬細(xì)小病毒研究進(jìn)展[J].吉林畜牧獸醫(yī),2016,37(8):13-15.
HU N,ZOU X Y,CAI J S.The research progress of canine parvovirus[J].Jilin Animal Husbandry and Veterinary Medicine,2016,37(8):13-15.(in Chinese)
[5]" 孫" 梅,吳建華,白文軍.犬細(xì)小病毒研究進(jìn)展[J].北京農(nóng)業(yè),2013(36):175-179.
SUN M,WU J H,BAI W J.Research progress on canine parvovirus[J].Beijing Agriculture,2013(36):175-179.(in Chinese)
[6]" 史才興,梁水菁,李建璋,等.紅景天苷藥理作用及其機制研究進(jìn)展[J].菏澤醫(yī)學(xué)專科學(xué)校學(xué)報,2021,33(4):67-70,84.
SHI C X,LIANG S J,LI J Z,et al.Research progress on the pharmacological effects and mechanisms of astragalus polysaccharide[J].Journal of Heze Medical College,2021,33(4):67-70,84.(in Chinese)
[7]" 崔晉龍,付少彬,王夢亮.紅景天苷生物、化學(xué)和生物催化合成的分子理論及應(yīng)用[J].天然產(chǎn)物研究與開發(fā),2013,25(6):851-855,840.
CUI J L,F(xiàn)U S B,WANG M L.Advance in studies on biosynthesis,chemical synthesis and biocatalysis synthesis of salidroside[J].Natural Product Research and Development,2013,25(6):851-855,840.(in Chinese)
[8]" LI Y T,ZHAO Y Y,LI X T,et al.Characterization of global metabolic profile of Rhodiola crenulata after oral administration in rat plasma,urine,bile and feces based on UHPLC-FT-ICR MS[J].J Pharm Biomed Anal,2018,149:318-328.
[9]" GUAN S,F(xiàn)ENG H H,SONG B C,et al.Salidroside attenuates LPS-induced pro-inflammatory cytokine responses and improves survival in murine endotoxemia[J].Int Immunopharmacol,2011,11(12):2194-2199.
[10]" GAO J,ZHOU R,YOU X T,et al.Salidroside suppresses inflammation in a D-galactose-induced rat model of Alzheimer's disease via SIRT1/NF-κB pathway[J].Metab Brain Dis,2016,31(4):771-778.
[11]" WANG H B,DING Y Y,ZHOU J,et al.The in vitro and in vivo antiviral effects of salidroside from Rhodiola rosea L. against coxsackievirus B3[J].Phytomedicine,2009,16(2-3):146-155.
[12]" 張" 霞,任秋生,王海鵬,等.紅景天苷體外抗EV71病毒的作用[J].湖北農(nóng)業(yè)科學(xué),2013,52(15):3605-3608.
ZHANG X,REN Q S,WANG H P,et al.Inhibitory effect of salidroside against enterovirus 71 in vitro[J].Hubei Agricultural Sciences,2013,52(15):3605-3608.(in Chinese)
[13]" ZHAO X J,LU Y,TAO Y,et al.Salidroside liposome formulation enhances the activity of dendritic cells and immune responses[J].Int Immunopharmacol,2013,17(4):1134-1140.
[14]" LIU K,XU P,LI Y C,et al.Inhibition of canine parvovirus 2 (CPV-2) replication by TAT-scFv through targeting of the viral structural protein VP2 of CPV-2[J].New Microbiol,2024,46(4):381-389.
[15]" SHARMA N,MISHRA K P,GANJU L.Correction to:salidroside exhibits anti-dengue virus activity by upregulating host innate immune factors[J].Arch Virol,2022,167(3):1011.
[16]" ELLIS R E,JACOBSON D M,HORVITZ H R.Genes required for the engulfment of cell corpses during programmed cell death in Caenorhabditis elegans[J].Genetics,1991,129(1):79-94.
[17]" GALLUZZI L,BRENNER C,MORSELLI E,et al.Viral control of mitochondrial apoptosis[J].PLoS Pathog,2008,4(5):e1000018.
[18]" ANGELOVA A L,APRAHAMIAN M,GREKOVA S P,et al.Improvement of gemcitabine-based therapy of pancreatic carcinoma by means of oncolytic parvovirus H-1PV[J].Clin Cancer Res,2009,15(2):511-519.
[19]" DOLEY J,SINGH L V,KUMAR G R,et al.Canine parvovirus type 2a (CPV-2a)-induced apoptosis in MDCK involves both extrinsic and intrinsic pathways[J].Appl Biochem Biotechnol,2014,172(1):497-508.
[20]" 張傳鵬.犬細(xì)小病毒SH15株的分離鑒定及其誘導(dǎo)MDCK細(xì)胞凋亡的探究[D].上海:上海交通大學(xué),2017.
ZHANG C P.Isoaltion and identification of canine parvovirus SH15 and studies on apoptosis induced by canine parvovirus in MDCK cells[D].Shanghai:Shanghai Jiao Tong University,2017.(in Chinese)
[21]" MLLER D N,BINGER K J,RIEDIGER F.Prorenin receptor regulates more than the renin-angiotensin system[J].Ann Med,2012,44 Suppl 1:S43-S48.
[22]" LI S F,ZHANG Y L,GUAN Z Q,et al.SARS-CoV-2 triggers inflammatory responses and cell death through caspase-8 activation[J].Signal Transduct Target Ther,2020,5(1):235.
[23]" GURUNG P,KANNEGANTI T D.Novel roles for caspase-8 in IL-1β and inflammasome regulation[J].Am J Pathol,2015,185(1):17-25.
[24]" MAELFAIT J,VERCAMMEN E,JANSSENS S,et al.Stimulation of Toll-like receptor 3 and 4 induces interleukin-1β maturation by caspase-8[J].J Exp Med,2008,205(9):1967-1973.
[25]" WANG Q H,KUANG H X,SU Y,et al.Naturally derived anti-inflammatory compounds from Chinese medicinal plants[J].J Ethnopharmacol,2013,146(1):9-39.
[26]" SONG D,ZHAO M,F(xiàn)ENG L X,et al.Salidroside attenuates acute lung injury via inhibition of inflammatory cytokine production[J].Biomed Pharmacother,2021,142:111949.
[27]" GURUNG P,ANAND P K,MALIREDDI R K S,et al.FADD and caspase-8 mediate priming and activation of the canonical and noncanonical Nlrp3 inflammasomes[J].J Immunol,2014,192(4):1835-1846.
[28]" MAN S M,TOURLOMOUSIS P,HOPKINS L,et al.Salmonella infection induces recruitment of caspase-8 to the inflammasome to modulate IL-1β production[J].J Immunol,2013,191(10):5239-5246.
[29]" KANG T B,YANG S H,TOTH B,et al.Caspase-8 blocks kinase RIPK3-mediated activation of the NLRP3 inflammasome[J].Immunity,2013,38(1):27-40.
[30]" GHAYUR T,BANERJEE S,HUGUNIN M,et al.Caspase-1 processes IFN-γ-inducing factor and regulates LPS-induced IFN-γ production[J].Nature,1997,386(6625):619-623.
[31]" LI P,ALLEN H,BANERJEE S,et al.Mice deficient in IL-1 β-converting enzyme are defective in production of mature IL-1 β and resistant to endotoxic shock[J].Cell,1995,80(3):401-411.
(編輯" 范子娟)
猜你喜歡
中國中藥雜志(2016年22期)2017-02-13 17:28:52
中國當(dāng)代醫(yī)藥(2016年19期)2016-09-30 20:43:59
科技視界(2016年15期)2016-06-30 12:27:37
中國實用醫(yī)藥(2016年11期)2016-05-04 21:56:32
成長·讀寫月刊(2015年9期)2015-11-09 21:32:14
中國醫(yī)藥導(dǎo)報(2015年26期)2015-10-16 20:43:00
中國當(dāng)代醫(yī)藥(2015年18期)2015-08-06 18:09:34
中國現(xiàn)代醫(yī)生(2015年13期)2015-06-17 10:19:05
中國醫(yī)藥導(dǎo)報(2015年1期)2015-01-28 20:28:43
