基于高通量測序的銹腐病西洋參轉錄組分析
2024-12-31 00:00:00龐玉雪陳磊于紅霞王洪濤
山東農業科學 2024年7期

關鍵詞:西洋參:銹腐病:轉錄組測序;差異表達基因:功能聚類:通路富集
西洋參(Panax quinquefolius)別名花旗參、美國人參等,為多年生五加科人參屬草本植物,具有極高的藥用及經濟價值,原產于北美,20世紀80年代我國引種成功后,種植面積逐漸擴大,目前山東省威海市文登區已成為國內最大的西洋參生產區。西洋參栽培周期較長,生長環境較為復雜,容易受到多種病害的影響。銹腐病是對西洋參危害最大、最常見的病害之一,是一種由毀壞柱孢菌(Cylindrocarpon destructans)引起的土傳病害,一般年發病率在30%左右,患病參根變色腐爛,嚴重影響西洋參產量和品質,也是導致西洋參連作障礙的病害因素之一。化學防治是近年來針對西洋參病害的主要防控方法。但化學農藥的過量使用不僅造成西洋參的農藥殘留超標、加劇土壤微生態的失衡,還會導致致病菌的抗藥性不斷增強,造成用藥量和病害相互遞增的惡性循環。因此,解析西洋參抗銹腐病的機制,選育抗病性優良的西洋參新品種是從根本上減少銹腐病發生的重要方法。
高通量測序技術作為一種快速且高效的研究方法可從轉錄組水平上揭示生物的抗病機制,近年來已成功應用到植物病理研究中。高小寧等研究感染褐銹病甘蔗葉片的基因表達情況.鑒定出差異表達基因4716個,發現一系列與苯丙烷生物合成、類黃酮生物合成等有關的基因可能參與了甘蔗葉片抵御褐銹病菌脅迫,為甘蔗的抗病育種提供了理論支持。蘭黎明等通過轉錄組測序研究湖北海棠抗白粉病機制,發現碳代謝通路只在抗性強的湖北海棠中顯著富集,為蘋果屬植物抗白粉病分子機制研究提供了一定的基礎。Shao等通過轉錄組測序分析了陸地棉對黃萎菌接種的響應,發現代謝途徑、次生代謝產物的生物合成及植物與病原菌互作在陸地棉抵御黃萎菌侵染中起到重要作用,這可為培育抗黃萎菌陸地棉新品種提供理論參考。目前,對西洋參銹腐病的研究大多集中在病原菌的分類鑒定及生物學特性、拮抗菌的篩選與鑒定、根際土壤微生物群落分析等方面,而在轉錄組水平上對異質生境中感染銹腐病的西洋參相關基因的表達特征研究相對有限。因此,本研究基于高通量測序技術,在轉錄組水平上發掘與西洋參抗銹腐病相關的基因,分析與抗病相關的代謝通路,以期為深入解析其抗病機理提供理論參考。
1材料與方法
1.1試驗材料
在2022年7月末西洋參生長旺季,采集山東省威海市文登區侯家鎮同一管理方式下種植的兩年生西洋參為試材,依據西洋參的患病情況,將試材分為感病組BL、BR(患銹腐病西洋參葉片和根)和對照組AL、AR(健康西洋參葉片和根)。對照組取與感病組長勢一致的健康植株作為一個生物學重復。所有樣品采用液氮速凍后-80℃冷凍保存,每組設3次重復。
1.2RNA提取、cDNA文庫構建及測序
將凍存樣品在液氮中研磨后,用天根RNAprep Pure植物總RNA提取試劑盒提取西洋參葉片及根組織的總RNA。采用NanoDrop 2000超微量分光光度計和Agilent 2100生物分析儀分別檢測RNA的純度和質量。cDNA文庫的構建以及轉錄組測序均委托上海歐易生物醫學科技有限公司完成。利用Illumina高通量測序平臺完成西洋參樣品的轉錄組測序,獲得原始序列數據。
1.3測序數據處理
采用Trimmomatic軟件對測序所得的原始數據進行預處理,去掉接頭及不合格序列,最終獲得高質量的Clean reads。利用HISAT2對質控后的Clean reads與西洋參基因組進行比較,獲取其在參考基因組上的定位信息以及測序樣本特有的序列特征信息。
1.4基因表達水平分析
以FPKM(Fragments Per Kilobase per Millionmapped reads)作為檢測基因表達水平的指標繪制基因表達箱線圖,分析表達數據分布的分散程度。基于基因的表達水平進行主成分分析(PCA),考察基因表達的組間相似性和差異性。
1.5差異表達基因篩選、聚類分析及注釋
使用R語言程序包DESeq 2軟件,以Plt;0.05且log2(FoldChange)lgt;1為條件,篩選感病組與對照組間的差異表達基因( DEGs),得到感病組葉片(BL)與對照組葉片(AL)之間差異表達基因組別BL-vs-AL,感病組根(BR)與對照組根(AR)之間差異表達基因組別BR-vs-AR。同時采用非監督層次聚類的方法,獲得各樣本間的DEGs熱圖。對篩選出的DEGs進行GO功能分析和KEGG通路分析,確定DEGs參與的主要生物學功能和途徑。
1.6差異表達基因的qRT-PCR驗證
隨機選取7個DEGs(上調4個、下調3個),以西洋參GAPDH為內參基因,進行qRT-PCR驗證。利用軟件Primer Premier 5.0設計引物,采用天根FastKing cDNA合成試劑盒將總RNA反轉錄合成cDNA。基因相對表達量通過2-△△法進行計算,每個樣本3次重復,與RNA-seq表達水平進行對比分析。qRT-PCR引物如表1所示。
2結果與分析
2.1轉錄組數據質量分析及比對統計
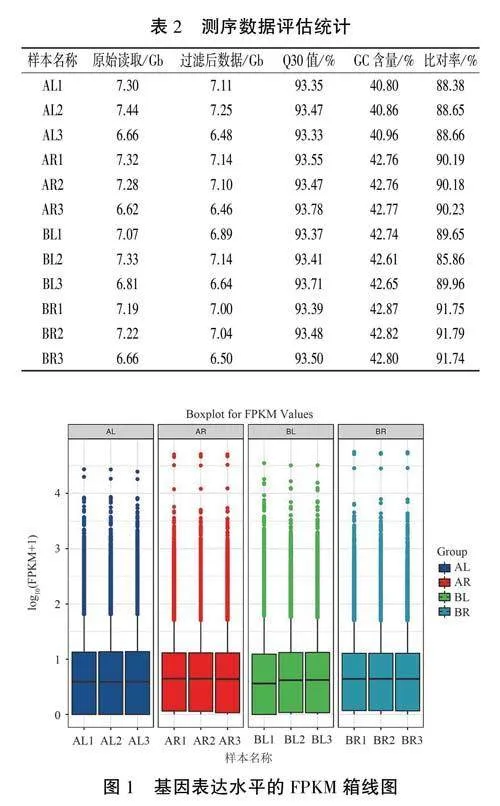
對12個樣本進行測序質量篩選,共獲得Clean data 84.90Gb,所有樣本的有效數據量分布在6.46~7.25Gb之間,平均GC含量為42.28%,Q30堿基百分比均超過93.30%。分別將12個樣本的Clean reads與西洋參參考基因的基因組進行序列比對,比對率在85.86%~91.79%之間,說明各樣本測序質量良好,可用于后續分析(表2)。
2.2基因表達分析
以FPKM作為量化基因表達水平的指標,對12個樣本的FPKM做箱線圖(圖1),FPKM密度分布表明,在離散度和總體分布度上,感病組(BL、BR)與對照組(AL、AR)的基因總體表達量均有不同程度的差異,說明感染銹腐病后西洋參葉片和根中某些基因表達發生了變化。
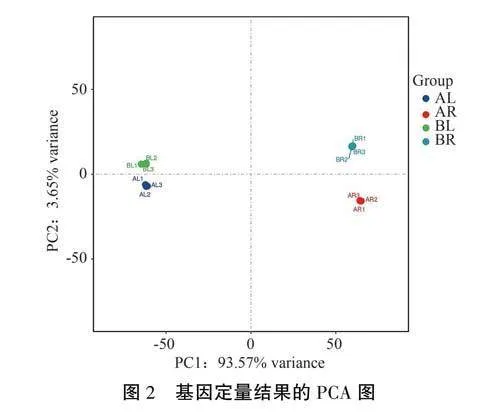
對12個樣本進行主成分分析,結果(圖2)表明,同一組樣本的3個生物學重復聚集在一起,說明各生物重復之間的相似度較高,可以滿足后續差異基因分析的需要。此外,感病組(BL、BR)和對照組(AL、AR)的樣品出現明顯分離,分別分布于差異極顯著的不同群組,說明西洋參感染銹腐病后基因表達譜發生了顯著變化。
2.3差異表達基因分析
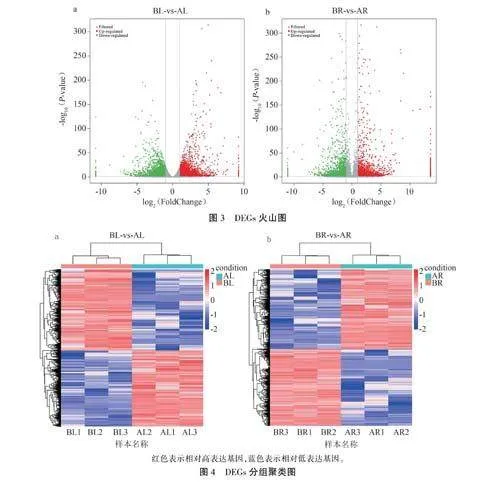
2.3.1差異表達基因篩選及聚類分析以Plt;0.05且llog2( FoldChange)lgt;1為篩選條件,采用DESeq 2檢測感病組與對照組間的DEGs。在BL-vs-AL組中共發現8642個DEGs.包括4463個上調表達,4179個下調表達(圖3a);在BR-vs-AR組中共鑒定出5308個DEGs,包括2609個上調表達,2699個下調表達(圖3b)。表明受到銹腐病菌侵害后,西洋參可通過提高或抑制某些基因的表達來增強自身對抗銹腐病的能力。
在此基礎上對DEGs展開聚類分析,結果表明,感病組和對照組存在著顯著差異。感病組葉片(BL)上調表達的4463個基因分布在上部,而對照組(AL)則呈現出相反的表達模式(圖4a)。感病組根(BR)上調表達的2609個基因分布在下部,而對照組(AR)則相反(圖4b)。說明大量基因在西洋參感染銹腐病后呈現差異表達模式。
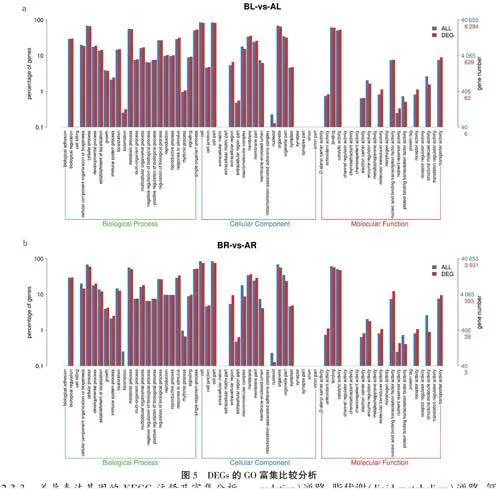
2.3.2差異表達基因GO富集分析利用GO數據庫對DEGs進行功能注釋,分別挑選top23、top20、top21的亞類進行展示。BL-vs-AL組中共有6294個DEGs獲得功能注釋(圖Sa),BR-vs-AR組中共有3931個DEGs獲得功能注釋(圖Sb),其中,生物學過程(biological process)主要富集于細胞過程(cellular process)、代謝過程(meta-bolic process)、單有機體過程(single-organlsmprocess)、生物調節(biological regulation)、應激反應(response to stimulus)等;細胞組成(cellularcomponent)主要富集在細胞(cell)、細胞組分(cellpart)、細胞器(organelle)、膜(membrane)等;分子功能(molecular function)主要富集在黏合(bind-ing)、催化活性(catalytic activity)、核酸結合轉錄因子活性(nucleic acid binding transcription factoractivity)、轉運活性(transporter
activity)等。
2.3.3差異表達基因的KEGG注釋及富集分析
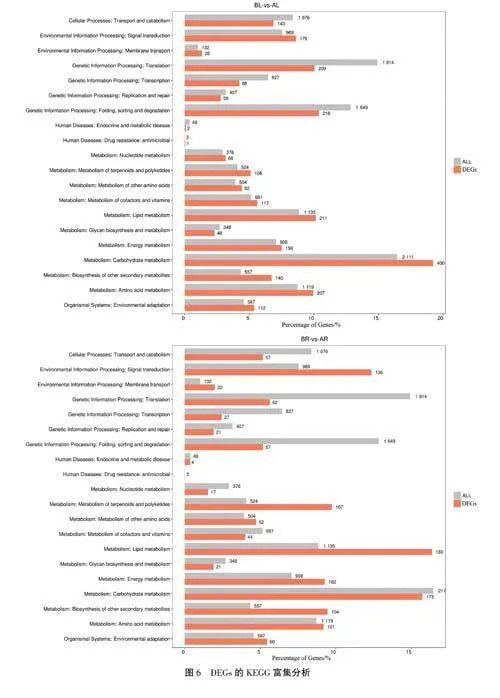
為了解DEGs主要參與的代謝和信號轉導途徑,利用KEGG數據庫對DEGs進行代謝通路分析,選取代謝通路中top20的通路進行展示。BL-vs-AL組中注釋到最多的是有400個DEGs參與碳水化合物代謝(carbohydrate metabolism)通路,其次是折疊、分類和降解(folding,sorting and deg-radation)通路、脂代謝(lipid metabolism)通路、氨基酸代謝(amino acid metabolism)通路等(圖6a)。
BR-vs - AR組中注釋到DEGs最多的是有180個DEGs參與脂代謝(lipid metabolism)通路,其次是碳水化合物代謝(carbohydrate metabolism)通路、信號轉導(signal transport)通路等(圖6b)。
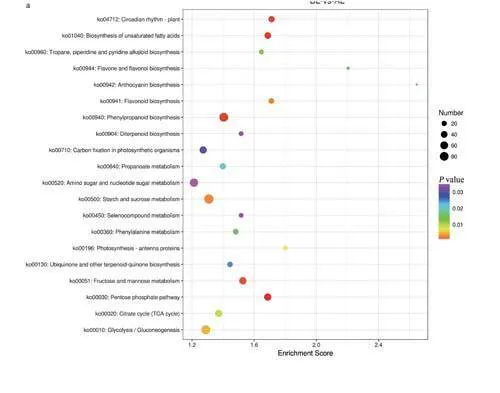
結合KEGG注釋結果,對DEGs進行信號通路富集分析,計算各條目中DEGs富集的顯著性,篩選出P值最低的前20條KEGG通路進行展不。BL-vs-AL組中花青素生物合成(anthocyaninbiosynthesis)通路的富集水平最顯著,其次是黃酮和黃酮醇的生物合成(flavone and flavonol biosyn-thesis)。苯丙素生物合成(phenylpropanoid bio-synthesis)、淀粉和蔗糖代謝(starch and sucrosemetabolism)、氨基糖和核苷酸糖代謝(amino sugarand nucleotide sugar metabolism)是富集到DEGs最多的3個通路(圖7a)。
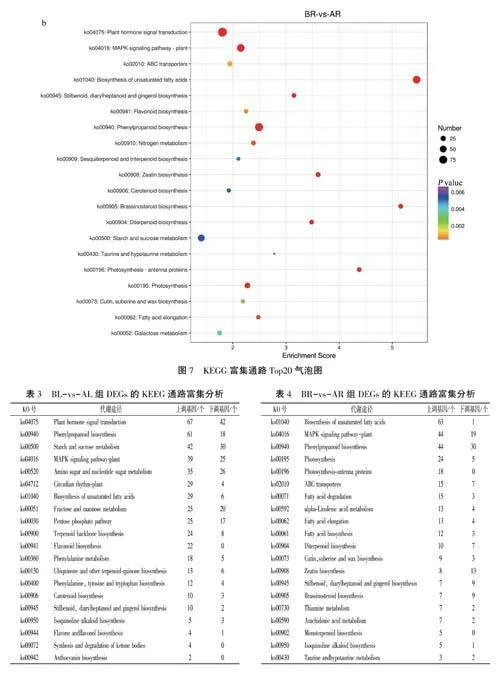
BR-vs-AR組中不飽和脂肪酸的生物合成(biosynthesis of unsaturated fatty acids)通路的富集水平最顯著,其次是油菜素內酯生物合成(brassinosteroid
biosynthesis)、光合作用一天線蛋白(photosynthesis-antenna
proteins)。苯丙素生物合成(phenylpropanoid biosynthesis),不飽和脂肪酸的生物合成(biosynthesis of unsaturated fattyacids)、MAPK信號通路一植物(MAPK signalingpathway-plant)是富集到DEGs最多的3個通路(圖7b)。
BL-vs-AL、BR-vs-AR兩組KEGG通路中上調及下調DEGs的富集情況見表3和表4。從上調DEGs的富集水平可以看出,西洋參感染銹腐病時,苯丙素生物合成(phenylpropanoid biosynthe-sis)、MAPK信號通路一植物(MAPK signalingpathway-plant)、不飽和脂肪酸的生物合成(bio-synthesis of unsaturated fatty acids)等與植物抗逆性相關的代謝途徑在兩組中共同顯著富集。
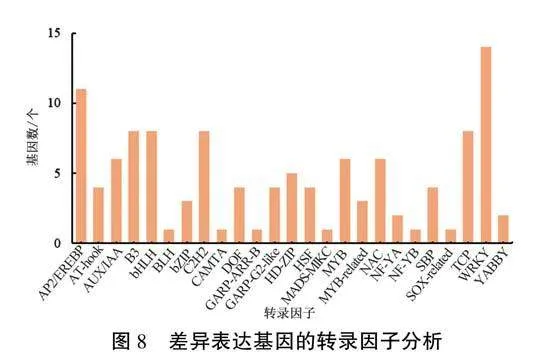
2.4差異表達基因的轉錄因子分析
對BL-vs-AL與BR-vs-AR兩組中共有的1458個DEGs進行轉錄因子分析(圖8),共預測到轉錄因子116個,分別歸屬到25類轉錄因子家族中。其中,WRKY家族的基因最多,包含14個,其次是AP2/EREBP家族的基因,共11個,B3家族、bHLH家族、TCP家族和C2H2家族的基因各有8個,MYB家族與AUX/IAA家族的基因分別為6個,其余家族擁有的基因數目較少。這些轉錄因子家族大多與植物的抗病性密切相關。
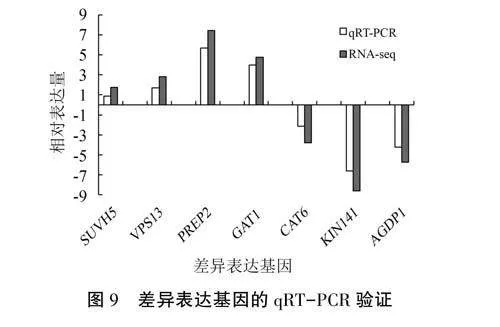
2.5DEGs的qRT-PCR驗證
隨機挑選7個DEGs(SUVH5、VPS13、PREP2、GAT1、CAT6、KIN141、AGDP1),以西洋參GAPDH基因為內參基因,利用qRT-PCR技術對轉錄組測序得到的數據進行驗證。結果(圖9)表明,4個上調及3個下調DEGs的表達模式與轉錄組測序分析結果一致,說明轉錄組數據分析的可靠性較高。
3討論與結論
植物對病原微生物侵染的應答是一個非常復雜的代謝過程,植物體一般通過酶的催化活性來防御病原菌的危害。超氧化物歧化酶(SOD)、過氧化氫酶(CAT)和過氧化物酶(POD)等是一類廣泛分布于多種植物組織中與植物體內抗氧化性能相關的酶,能起到對植物活性氧的催化作用,達到抗氧化效果。植物體內抗氧化酶活力的高低與其自身的代謝能力及對環境的適應能力有很大關系。當受到多種逆境脅迫時,植物體內的防衛酶活力均會升高。孫嘉曼等研究發現,感染銹腐病后,人參根部和葉片中的主要防御酶(POD、CAT和SOD)活性均明顯增強。結合前人的研究成果,本研究對BL-vs-AL、BR-vs-AR組中的DEGs進行GO功能注釋后發現,患銹腐病西洋參的DEGs顯著富集在催化活性、代謝途徑等與酶催化活性有關的GO條目上,且主要為上調表達模式,且與防御相關的CAT等基因的表達量均上調顯著,表明在受到銹腐病菌侵染時,西洋參葉片與根系中相關保護性酶系統的動態平衡被破壞,從而使其能夠抵抗病原菌的危害維持自身的正常功能。
有研究證明,脂類代謝與合成是植物與微生物相互作用中的重要環節,脂類及其代謝途徑在植物與病原菌互作中發揮著重要作用,也是影響植物抗病反應的重要因素。植物體的眾多脂類中,不飽和脂肪酸是植物細胞內一類重要代謝物質,是構成細胞膜的主要成分。本研究發現,兩組中分別有211、180個DEGs被注釋到了脂代謝通路中,且富集到不飽和脂肪酸的生物合成途徑中的DEGs主要呈上調表達。結合前人的研究推測,受到銹腐病菌侵害時,西洋參通過合成不飽和脂肪酸來提高細胞膜的穩定性,從而降低對自身的傷害。
碳水化合物代謝已被證實與植物抗病性緊密相關,增加糖類物質含量可提高植物的抗病能力。本研究KEGG分析發現,受到銹腐病感染后,兩組中有大量的DEGs富集到了碳水化合物代謝通路中。這與Gao等在桃響應可可毛色二孢菌(Lasiodiplodia theobromae)侵染的轉錄組分析結果相吻合。推測西洋參可通過增強體內糖類物質的合成來抵御銹腐病菌的入侵。
KEGG分析還發現,兩組DEGs在MAPK信號通路一植物途徑中顯著富集,并以上調表達為主。MAPK是一種重要的信號轉導分子,可將外界刺激從細胞表面傳導到細胞核內部,植物MAPK級聯在信號轉導植物防御病原體攻擊中起關鍵作用。也有研究表明MAPK組分在水稻激素信號轉導中有著重要的生物學意義。
氨基酸次生代謝產物中,苯丙素類在植物生長調節和抵御病害中發揮了重要作用。其中苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)作為苯丙烷類化合物代謝的關鍵限速酶,可將L-苯丙氨酸催化生成反式肉桂酸,并經其他酶進一步轉化為木質素、類黃酮、酚類等。木質素是細胞次生壁的重要組分,能為細胞提供機械支撐,并能保持水分,進行營養物質的運輸;類黃酮和花青素類物質在植物體內作為一種重要的抗氧化劑,具有抗病毒、抗氧化、抗逆境脅迫等作用,也可作為信號分子介導植物一微生物相互作用,在植物一病原菌互作中發揮著緊要作用。Zhou等研究發現,在患油斑病的青檸中,苯丙素生物合成被顯著激活,鑒定出79個與木質素積累相關的DEGs,其中65個顯著上調,表明柑橘類果實可能通過木質素積累增強自身對油斑病的抵抗能力。本研究顯示,西洋參受到銹腐病菌的侵害后,葉中花青素生物合成途徑及黃酮和黃酮醇的生物合成通路富集水平最顯著:且苯丙素生物合成途徑在根與葉中共同顯著富集,其中PAL基因全部呈現上調表達。但PAL基因是否參與了西洋參對抗銹腐病菌還有待進一步研究。
轉錄因子(transcription factor,TF)作為一類具有調節功能的基因,在植物對病原菌的抗性應答過程中起到了非常重要的作用。它們能夠與基因啟動子區域中的順式元件特異性結合,從而調控靶基因的表達。關于宿主植物對病原菌脅迫應答過程中的一些關鍵轉錄因子如WRKY、MYB、bHLH及AP2/EREBP等已經被大量研究。有文獻表明,WRKY家族成員在植物受病原微生物侵染時,其轉錄、蛋白合成和與寄主的結合均發生顯著變化來調節各種抗性反應,小麥抗葉銹病過程中WRKY基因家族起重要的調控作用。AP2/EREBP TFs為植物所特有,過量表達EREBP/ERF轉錄因子能明顯提高轉基因植株的抗病能力。bHLH轉錄因子在真核生物的生長發育和脅迫響應中扮演著重要角色’bHLH家族有助于調控小麥對條銹菌與假禾谷鐮孢菌脅迫的響應。有研究者發現,C2H2型鋅指蛋白可以激活防衛反應相關基因表達,煙草中的ZFT1基因受煙草花葉病毒侵染后,其表達水平發生了變化以對脅迫做出應答。B3家族及TCP蛋白家族在調控植物的生長發育和多種逆境脅迫應答中發揮重要作用。本研究對轉錄因子進行分析后發現,WRKY家族、AP2/EREBP家族、B3家族、bHLH家族、TCP家族和C2H2家族包含的轉錄因子較多,初步推測可能與西洋參受銹腐病侵害后的發病過程相關。
綜合本研究結果,推測西洋參通過上述代謝途徑提高自身抗病能力,同時也說明其抗病機制十分復雜,多條代謝途徑共同作用以維持其在病原體侵害后的正常功能。目前國內外關于植物抗病機制的轉錄組分析研究多數停留在實驗室階段,主要手段是將植物移栽于實驗室后,通過人工手段使植物感染各種病原菌來分析植物的生理生態響應。然而,實驗室的人工環境不同于植物生長所處的自然環境,無法真實地反映植物在復雜多變的異質生境中特有的抗病機理。本研究通過分析自然環境中西洋參感染銹腐病后的基因表達信息,明確了異質生境中感染銹腐病的西洋參相關基因的表達特征及相關代謝通路,可為深入研究西洋參抗銹腐病的分子機制提供一定的理論參考。
