404 Not Found
404 Not Found
大腸桿菌O-抗原血清型鑒定研究進展
劉璨穎,張濟培,王丙云
?
大腸桿菌O-抗原血清型鑒定研究進展
劉璨穎,張濟培,王丙云
大腸桿菌在一定條件下能引發(fā)宿主疾病,其表面O-抗原與毒力有關(guān)。O-抗原的化學(xué)組成和結(jié)構(gòu)具有高度多樣性,并且O-抗原血清型種類與大腸桿菌致病性有一定聯(lián)系。因此,大腸桿菌O-抗原血清型鑒定對流行病學(xué)調(diào)查,防御和控制致病性大腸桿菌病有重要意義。傳統(tǒng)的血清學(xué)分型方法耗時長、費用高、準確度不理想。隨著科學(xué)技術(shù)的發(fā)展,研究者對大腸桿菌196種O-抗原基因簇進行了破譯,比較分析了不同O-抗原基因簇序列,并針對基因簇中特異性DNA序列設(shè)計分子標記,運用PCR方法對O-抗原血清型進行分型,其中包括一般PCR、多重PCR、實時PCR、DNA芯片和微球懸浮列陣法。此外,還有rbf-限制性片段長度多態(tài)性分析法,磁性微球免疫分析法和基于全基因組序列預(yù)測法,這些方法豐富了O-抗原血清型鑒定方法,彌補了傳統(tǒng)血清學(xué)分型方法的不足。本文概述了O-抗原合成基因簇序列和O-抗原血清型鑒定方法相關(guān)研究進展。
大腸桿菌;O-抗原;血清型分型
大腸桿菌廣泛存在于自然界中,致病性大腸桿菌是一類重要的人獸共患病病原菌,依據(jù)致病特性分為腸內(nèi)致病性和腸外致病性大腸桿菌。腸內(nèi)致病性大腸桿菌能分泌毒素,引發(fā)宿主水樣、血樣腹瀉,甚至導(dǎo)致全身性臨床癥狀,如溶血尿毒癥和腎神經(jīng)后遺癥。腸外致病性大腸桿菌能侵襲并定殖于宿主胃腸道外組織,并誘發(fā)感染,包括腦膜炎、敗血癥、泌尿道感染和呼吸道感染等。大腸桿菌菌株間差異大,常按照種系發(fā)育群、生物表型、致病特性、抗原特性等進行分類區(qū)分[1-2]。
菌體O-抗原,表面K-抗原和鞭毛H-抗原是大腸桿菌分型的主要表面抗原[3]。O-抗原是大腸桿菌脂多糖的最外層結(jié)構(gòu),與細菌對環(huán)境的適應(yīng)性有關(guān),是細菌的毒力因子,也是細菌噬菌體和宿主免疫系統(tǒng)的主要目標,能在宿主體內(nèi)誘發(fā)強烈的免疫反應(yīng)[4-6]。?rskov于1977年提出了綜合分型方案,將大腸桿菌O-抗原分為164種,該方案是分類學(xué)、流行病學(xué)調(diào)查、疾病暴發(fā)時菌株區(qū)分以及監(jiān)控中大腸桿菌分型的主要標準[7]。隨著研究成果的不斷更新,報道有O1-O187血清型,其中6種O-抗原血清型O31、O47、O67、O72、O94和O122被撤消,4種被分為亞型O18ab/ac,O28ab/ac,O112ab/ac和O125ab/ac。此外,還有11種OX-型。因此目前大腸桿菌O-抗原血清型共有196種[8]。
O-抗原血清型種類與大腸桿菌的致病性有一定的聯(lián)系,不同致病型大腸桿菌的優(yōu)勢O-抗原血清型不同。例如,腸出血性大腸桿菌的主要血清型為O157,E.coliO157菌株是重要的全球食源性病原菌[9];禽致病性大腸桿菌的優(yōu)勢血清型為O1、O2、O18和O78[10];與尿道感染有關(guān)的大腸桿菌,其血清型主要為O1、O2、O4、O6、O7、O16、O18、O25和O75[11];能引發(fā)新生兒腦膜炎的腸外致病性大腸桿菌菌株,其血清型主要為O18∶K1[12];我國患病豬場分離的豬源腸外致病性大腸桿菌菌株優(yōu)勢血清型依次為O11、O8、O138、O161和O101[13]。因此,快速、準確鑒定大腸桿菌O-抗原血清型將有助于流行病學(xué)調(diào)查,疾病診斷和疫苗研發(fā),對防御并控制致病性大腸桿菌傳播有重要意義。
1 O-抗原概述
1.1 O-抗原組成和特性 革蘭氏陰性菌外膜脂多糖由脂質(zhì)A,核心寡糖和O-抗原多糖3部分組成。脂質(zhì)A在大腸桿菌中非常保守,是毒性成分。核心寡糖區(qū)域分為內(nèi)部和外部區(qū)域,O-抗原多糖與核心寡糖外部區(qū)域的糖基相連,具有血清型特異性,由含有3-7個單糖的寡糖單位重復(fù)連接組成。單糖種類、數(shù)量、排列方式和寡糖間鏈的不同構(gòu)成了O-抗原的多樣性[6,14]。基因水平轉(zhuǎn)移是O抗原高度多樣性的主要原因。基因水平轉(zhuǎn)移、重組和點突變能使O-抗原血清型發(fā)生轉(zhuǎn)變[15-17]。此外,獲得位于細菌噬菌體或質(zhì)粒上的O-抗原修飾基因也能導(dǎo)致細菌O-抗原血清型的差異[18-19]。具有完整O-抗原的大腸桿菌為光滑型表型,而O-抗原發(fā)生突變或丟失時,大腸桿菌失去合成O-抗原的能力,為粗糙型表型[20]。
1.2 O-抗原合成基因簇 大腸桿菌中,大部分編碼O-抗原合成酶的基因在染色體上相鄰排列,形成一個約4.5 kb(O155,含有4個基因)-19.5 kb(O108,含有18個基因)的基因簇,被稱為rfb基因簇。該基因簇兩端常有持家基因galF(UTP-葡萄糖-1-磷酸尿甙基轉(zhuǎn)化酶編碼基因)和gnd(6-磷酸葡萄糖酸脫氫酶編碼基因),在染色體上位于可拉酸合成基因簇(wca基因)和組氨酸(his基因)合成操縱子之間[21]。DebRoy等對大腸桿菌196種O-抗原合成基因簇進行序列分析,結(jié)果顯示其中有21組(46種)序列相似性高達98%~99.9%,插入元件在O-抗原合成基因簇進化過程中發(fā)揮了重要作用[8]。但O-抗原血清型O14和O57型菌株galF和gnd基因間不含有O-抗原合成基因簇[22]。
O-抗原合成基因簇中主要包括3類基因:單糖合成酶基因、糖基轉(zhuǎn)移酶基因和寡糖單位處理酶基因。核苷酸糖合成基因參與O-抗原核苷酸糖前體的合成,在不同O-抗原血清型中有高度相似性,且常聚集成簇[14,8]。最新研究顯示有至少27%種O-抗原血清型,核苷酸糖合成基因不位于O-抗原合成基因簇中[22]。糖基轉(zhuǎn)移酶轉(zhuǎn)移不同糖前體形成寡糖,每種O-抗原合成基因簇中含有2~6個基因編碼假定糖基轉(zhuǎn)移酶。糖基轉(zhuǎn)移酶基因的點突變能導(dǎo)致O-抗原血清型間的差異[16]。O-抗原處理蛋白涉及O-抗原基本單位的轉(zhuǎn)運和聚合。
在大腸桿菌中合成O-抗原多糖的途徑為Wzy聚合酶依賴型途徑和三磷酸腺苷結(jié)合轉(zhuǎn)運盒(ATP-binding cassette, ABC)轉(zhuǎn)運子途徑。這兩種合成途徑中,基因wzx(糖基轉(zhuǎn)移酶編碼基因)/wzy(寡糖單位聚合酶編碼基因)和wzm(O-抗原ABC轉(zhuǎn)運子通透酶編碼基因)/wzt(ABC轉(zhuǎn)運子ATP-結(jié)合蛋白編碼基因)分別與O-抗原基本單元轉(zhuǎn)運和聚合有關(guān)。DebRoy等對大腸桿菌196種O-抗原合成基因簇進行分析,顯示185種基因簇中含有wzx和wzy基因,這兩個基因與運用Wzy聚合酶依賴型途徑合成和轉(zhuǎn)運O-抗原有關(guān),另外11種O-抗原血清型(O162,O101,O89,O92,O97,O52,O95,O60,O8,O99和O9)攜帶wzm和wzt基因,利用三磷酸腺苷結(jié)合轉(zhuǎn)運盒轉(zhuǎn)運子途徑處理O-抗原。并且依據(jù)Lguchi等對大腸桿菌182種O-抗原合成基因簇wzx/wzy或wzm/wzt同源基因序列分析結(jié)果顯示,137種O-抗原合成基因簇wzx/wzy或8種O-抗原合成基因簇wzm/wzt同源基因序列差異大,同源性<70%,具有一定的O-抗原合成基因簇特異性[8,22]。
2 O-抗原血清型分型方法
2.1 傳統(tǒng)血清學(xué)分型方法 傳統(tǒng)的O-抗原血清型鑒定方法是運用特異的抗血清進行血清凝集實驗,需要從檢測樣品中分離出細菌并進行富集,將細菌于100 ℃加熱2 h釋放出表面O-抗原后,與標準O-抗原血清型細菌的特異性抗血清進行血清凝集反應(yīng),眼觀判斷實驗結(jié)果。若菌體表面無脂多糖呈粗糙型,或者菌體發(fā)生自凝集,則不與抗血清發(fā)生凝集反應(yīng),不能用該方法進行鑒定。如O14型菌株為粗糙型表型,與抗血清不反應(yīng),不能進行血清學(xué)分型。此外血清學(xué)鑒定方法還受到血清質(zhì)量、血清來源、血清交叉反應(yīng)、以及菌體表面其他抗原成分的影響,使檢測準確度、檢出率和靈敏度不太理想[8, 23]。并且傳統(tǒng)的血清學(xué)分型方法耗時長,花費高,不利于大規(guī)模檢測。
2.2 基于特異性DNA序列的PCR分型方法 基于特異性DNA片段的PCR鑒定方法可以實現(xiàn)對生物樣品快速、準確和高效的檢測,能夠很大程度上彌補傳統(tǒng)血清學(xué)鑒定方法的不足。基因wzx/wzy或wzm/wzt具有一定的O-抗原血清型特異性,常被作為PCR和芯片分析鑒定大腸桿菌O抗原血清型的靶標。依據(jù)基因wzx/wzy或wzm/wzt的特異性DNA片段,設(shè)計并篩選出不同O-抗原血清型的特異性引物,運用PCR進行大腸桿菌O-抗原血清型鑒定[24-28]。但19.8%種O-抗原血清型中wzx/wzy基因不具有特異性,或特異區(qū)域過短,無法設(shè)計引物。如O73型、O17型、O44型、O77型與O106型大腸桿菌中wzx/wzy基因的核苷酸序列同源性≥95%;wzx_O46與wzx_O134基因序列同源性為99.7%,wzx_O46基因僅在3′端區(qū)域有2 bp的缺失突變[22]。也可依據(jù)rfb基因簇中差異區(qū)域DNA序列設(shè)計引物進行PCR鑒別。例如wzx/wzy-O62與wzx/wzy-O68基因的核苷酸序列同源性>97%,O62與O68型rfb基因簇序列的差異僅在于O62中含有一個748 bp的插入元件IS1,該插入元件位于rmlA基因末端。因此為了鑒定并區(qū)分大腸桿菌O62與O68型,應(yīng)在擴增出相應(yīng)wzx/wzy基因后,再在O62型rfb基因簇IS1元件兩側(cè)設(shè)計分子標識,依據(jù)PCR產(chǎn)物大小差別區(qū)分兩種O-抗原血清型(表1)[27]。目前,根據(jù)已報道的文獻統(tǒng)計,設(shè)計并篩選出特異性引物的O-抗原血清型種類有68種,DebRoy等于2011年統(tǒng)計了58種[29],本文補充了10種,包括O11,O12,O23,O62,O68,O116,O131,O140,O142和O163(表1)。
表1 基于大腸桿菌O-抗原合成基因簇特異序列的PCR引物
Tab.1 PCR primers based on specific sequence of E. coli O-antigen synthesis gene cluster
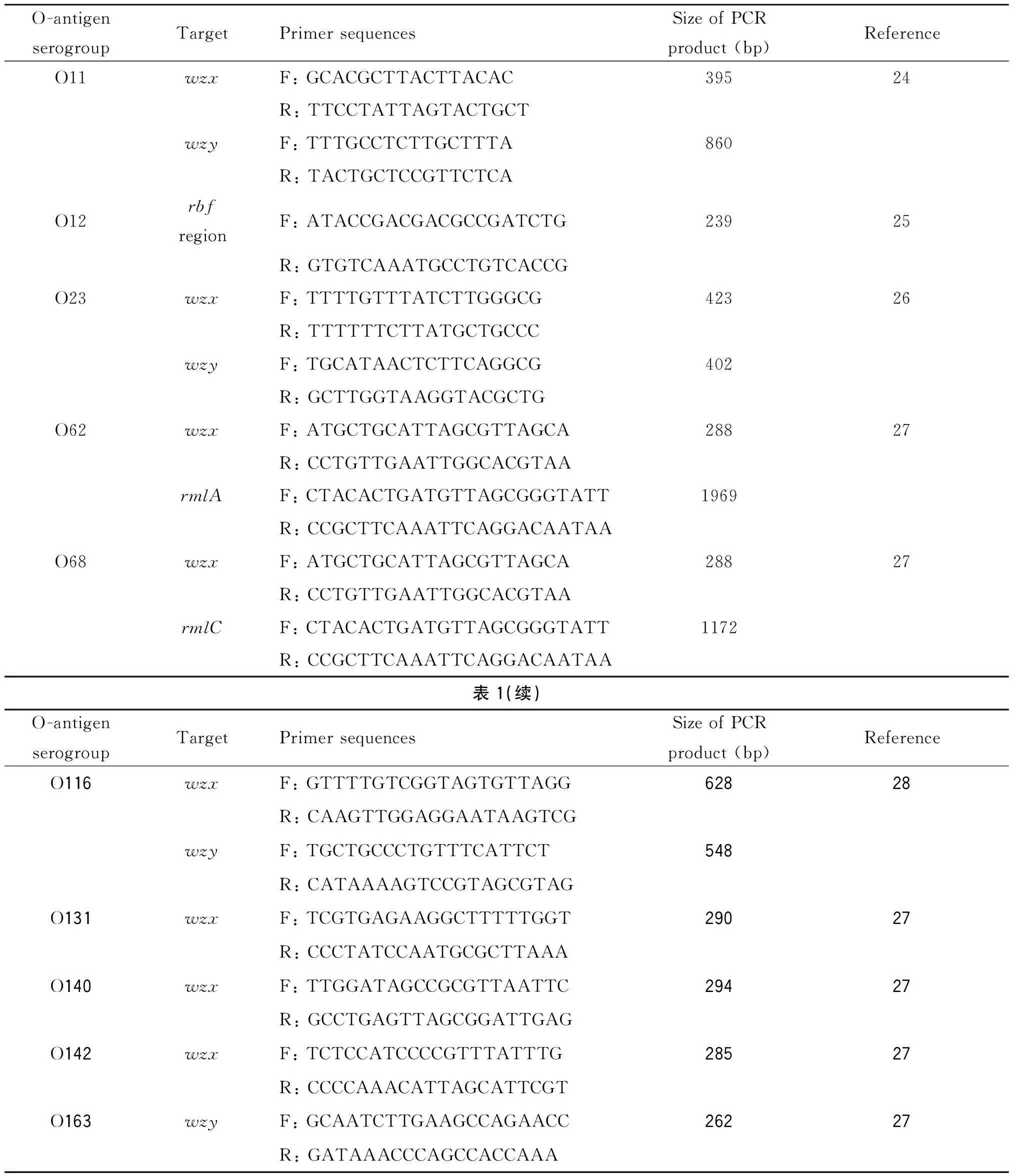
O-antigenserogroupTargetPrimersequencesSizeofPCRproduct(bp)ReferenceO11wzxF:GCACGCTTACTTACAC39524R:TTCCTATTAGTACTGCTwzyF:TTTGCCTCTTGCTTTA860R:TACTGCTCCGTTCTCAO12rbfregionF:ATACCGACGACGCCGATCTG23925R:GTGTCAAATGCCTGTCACCGO23wzxF:TTTTGTTTATCTTGGGCG42326R:TTTTTTCTTATGCTGCCCwzyF:TGCATAACTCTTCAGGCG402R:GCTTGGTAAGGTACGCTGO62wzxF:ATGCTGCATTAGCGTTAGCA28827R:CCTGTTGAATTGGCACGTAArmlAF:CTACACTGATGTTAGCGGGTATT1969R:CCGCTTCAAATTCAGGACAATAAO68wzxF:ATGCTGCATTAGCGTTAGCA28827R:CCTGTTGAATTGGCACGTAArmlCF:CTACACTGATGTTAGCGGGTATT1172R:CCGCTTCAAATTCAGGACAATAA表1(續(xù))O-antigenserogroupTargetPrimersequencesSizeofPCRproduct(bp)ReferenceO116wzxF:GTTTTGTCGGTAGTGTTAGG62828R:CAAGTTGGAGGAATAAGTCGwzyF:TGCTGCCCTGTTTCATTCT548R:CATAAAAGTCCGTAGCGTAGO131wzxF:TCGTGAGAAGGCTTTTTGGT29027R:CCCTATCCAATGCGCTTAAAO140wzxF:TTGGATAGCCGCGTTAATTC29427R:GCCTGAGTTAGCGGATTGAGO142wzxF:TCTCCATCCCCGTTTATTTG28527R:CCCCAAACATTAGCATTCGTO163wzyF:GCAATCTTGAAGCCAGAACC26227R:GATAAACCCAGCCACCAAA
針對O-抗原合成基因簇特異性序列設(shè)計引物進行一般PCR、多重PCR和實時PCR鑒定O-抗原血清型外[30-31],目前還延伸至DNA芯片法和微球懸浮列陣法。
DNA芯片法是將待檢大腸桿菌O-抗原合成基因簇中基因的PCR產(chǎn)物或寡聚核苷酸點在玻璃片上,然后與帶有標簽的特異O-抗原合成基因簇長片段PCR產(chǎn)物進行雜交,依據(jù)雜交信號判斷并鑒定大腸桿菌O-抗原血清型種類。DNA芯片法已用于腸產(chǎn)毒素大腸桿菌O-抗原血清型分型[33]。DNA芯片法優(yōu)勢在于能在同一平臺上一次性鑒定多種O-抗原血清型。
磁性微球懸浮列陣法中,微球外部結(jié)合有O-抗原血清型特異性DNA序列探針,微球內(nèi)部染有不同比例紅色和紅外熒光,可以產(chǎn)生100種獨特的光譜,因此每個樣本理論上可以分析高達100種不同的結(jié)合反應(yīng)。懸浮列陣由獨特?zé)晒馕⑶蚪M成。該方法,首先提取樣品DNA為模板,用生物素標記的多種O-抗原血清型檢測引物進行多重PCR擴增,將PCR反應(yīng)產(chǎn)物變性后與微球上帶有的特異性探針進行雜交反應(yīng),再用偶聯(lián)有強熒光藻類色素R藻紅蛋白的鏈霉素進行顯色檢測[34]。Lin等用該方法檢測了10種與產(chǎn)志賀氏毒素大腸桿菌相關(guān)的O-抗原血清型[35]。
2.3 rbf-限制性片段長度多態(tài)性分析法 rbf-限制性片段長度多態(tài)性分析法是利用長片段PCR方法擴增出待檢大腸桿菌整個rfb基因簇后,用限制性內(nèi)切酶MboII對PCR產(chǎn)物進行酶切,通過酶切后DNA片段凝膠電泳圖譜鑒定大腸桿菌O-抗原血清型。圖譜中條帶數(shù)目為5到25條,共有147種不同O-抗原限制性片段長度多態(tài)性圖譜。但其中有13種圖譜分別對應(yīng)兩種以上O-抗原血清型[36]。此外,由于圖譜中條帶較多,不易區(qū)分,且同一條帶中可能含有多條相同大小的酶切片段,結(jié)果分析易受干擾,且該分析方法耗時較長,使得rbf-限制性片段長度多態(tài)性分析法有一定的限制性。
2.4 磁性微球免疫分析法 磁性微球免疫分析法,與上述微球懸浮列陣法都是使用Luminex技術(shù),是一種復(fù)合微球感應(yīng)系統(tǒng)。采用該技術(shù)的分析,用微球表面共價固定的特異O-抗原血清型單克隆抗體來捕獲抗原。通過帶有生物素的二抗和紅藻蛋白報告子檢測到結(jié)合分析物。Luminex分析儀對結(jié)合分析物進行量化,從而進行多重檢測分析。微球免疫分析法的優(yōu)勢是可以同時進行多重分析,因此能夠降低檢測的費用,并且檢測時間較短,只需要3 h,此外該方法具有高靈敏度和特異性。Clotilde等利用該方法同時檢測了產(chǎn)志賀氏毒素大腸桿菌O-抗原血清型O157、志賀氏毒素1和2,并且證實與標準的酶聯(lián)免疫吸附實驗相比,該方法有上述優(yōu)勢[37]。
2.5 基于全基因組序列預(yù)測法 SerotypeFinder在線預(yù)測工具通過收集已知O-抗原血清型大腸桿菌菌株中wzx/wzy或wzm/wzt基因序列建立數(shù)據(jù)庫,將待測大腸桿菌菌株的全基因組草圖或完整序列與數(shù)據(jù)庫中序列進行BLAST比對,利用序列相似性來預(yù)測待測大腸桿菌菌株的O-抗原血清型。該方法相比于傳統(tǒng)血清學(xué)方法更為高效,但對于不同O-抗原血清型大腸桿菌中wzx/wzy或wzm/wzt基因有高度同源性時,鑒定結(jié)果不理想,且比對wzx或wzy基因可能有鑒定結(jié)果不一致的情況[38]。
3 結(jié) 語
大腸桿菌O-抗原血清型種類繁多,而血清型種類與大腸桿菌致病型有一定的聯(lián)系。因此,實現(xiàn)大規(guī)模準確、快速、便捷鑒定O-抗原血清型將有助于流行病學(xué)調(diào)查,疾病診斷和疫苗研發(fā),對防御并控制致病性大腸桿菌傳播有重要意義。隨著越來越多大腸桿菌基因組序列的公布和O-抗原合成基因簇的破譯,可以基于DNA序列,利用現(xiàn)代分子生物學(xué)方法彌補傳統(tǒng)血清學(xué)方法的不足,不斷完善O-抗原血清型鑒定方法。
目前已破譯196種O-抗原合成基因簇,但其中只有68種設(shè)計并篩選出了特異性分子標識。除去O-抗原合成基因簇序列同源性>98%的21組血清型(共46種),還有82種O-抗原血清型特異性分子標識有待確定。O-抗原合成基因簇序列高度相似的這21組血清型(46種)中,有4組(8種)不發(fā)生血清學(xué)交叉反應(yīng),包括O2型和O50型,O46型和O134型,O118型和O151型,OX19型和O11型,可以聯(lián)合現(xiàn)代分子生物學(xué)和傳統(tǒng)血清學(xué)方法進行O-抗原血清型鑒定。O-抗原合成基因簇序列高度相似的O-抗原血清型并不發(fā)生血清學(xué)交叉反應(yīng),可能是因為抗原決定簇相關(guān)蛋白在翻譯后期進行了不同的修飾,也可能是與O-抗原免疫學(xué)反應(yīng)有關(guān)的基因位于基因組中O-抗原合成基因簇以外的位置。對于不同O-抗原血清型大腸桿菌中O-抗原合成基因簇序列同源性>98%,并且傳統(tǒng)血清學(xué)鑒定又出現(xiàn)交叉反應(yīng)時,應(yīng)將這些O-抗原血清型進行融合,以免影響鑒定,包括O42型與O28ac型,O13型、O129型與O135型,O107型與O117型,O123型與O186型,O125ab型與O125ac型,O18ab型與O18ac型,OX6型與O168型,OX10型和O159型,OX21型和O163型,O38型和O128型,OX43型和O19型[8]。
大規(guī)模O-抗原血清型鑒定方法,如DNA芯片法,磁性微球懸浮列陣法,磁性微球免疫分析法和基于全基因組序列預(yù)測法等,提高了O-抗原血清型鑒定的效率和準確度,將是今后血清型鑒定發(fā)展的方向。
[1] Kaper JB, Nataro JP, Mobley HL. PathogenicEscherichiacoli[J]. Nat Rev Microbiol, 2004, 2: 123-140. DOI: 10.1038/nrmicro818
[2] Smith JL, Fratamico PM, Gunther NW. Extraintestinal pathogenicEscherichiacoli[J]. Foodborne Pathog Dis, 2007, 4(2): 134-163. DOI: 10.1089/fpd.2007.0087
[3] Kaufmann F. The serology of the coli group[J]. J Immunol, 1947, 57(1): 71-100.
[4] Raetz CR, Whitfield C. Lipopolysaccharide endotoxins[J]. Annu Rev Biochem, 2002, 71: 635-700.
[5] Lerouge I, Vanderleyden J. O-antigen structural variation: mechanisms and possible roles in animal/plant-microbe interactions[J]. FEMS Microbiol Rev, 2002, 26(1): 17-47.
[6] Wang L, Wang Q, Reeves PR. The variations of O antigens in gram negative bacteria[C]//. Endotoxins: Structure, Function and Recognition, 2010, 53: 123-152
[7] ?rskov I, ?rskov F, Jann B, et al. Serology, chemistry, and genetics of O and K antigens ofEscherichiacoli[J]. Bacteriol Rev, 1977, 41(3): 667-710.
[8] DebRoy C, Fratamico PM, Yan XH, et al. Comparison of O-antigen gene clusters of all O-serogroups ofEscherichiacoliand proposal for adopting a new nomenclature for O-typing[J]. PLoS One, 2016, 11(1): e0147434. DOI: 10.1371/journal. pone.0147434
[9] Paton AW, Paton JC. Direct detection of Shiga toxigenicEscherichiacolistrains belonging to serogroups O111, O157, and O113 by multiplex PCR[J]. J Clin Microbiol, 1999, 37(10): 3362-3365.
[10] Wang SH, Meng QM, Dai JJ, et al. Development of an allele-specific PCR assay for simultaneous sero-typing of avian pathogenicEscherichiacolipredominant O1, O2, O18 and O78 strains[J]. PLoS One, 2014, 9(5): e96904.
[11] Johnson JR. Virulence factors inEscherichiacoliurinary tract infection[J]. Clin Microbiol Rev, 1991, 4(1): 80-128.
[12] Johnson JR, Oswald E, O'Bryan TT, et al. Phylogenetic distribution of virulence-associated genes amongEscherichiacoliisolates associated with neonatal bacterial meningitis in the Netherlands[J]. J Infect Dis, 2002, 185(6): 774-784.
[13] Ding Y, Tang X, Lu P, et al. Clonal analysis and virulent traits of pathogenic extraintestinalEscherichiacoliisolates from swine in China[J]. BMC Vet Res, 2012, 8: 140.
[14] Stenutz R, Weintraub A, Widemalm G. The structures ofEscherichiacoliO-polysaccharide antigens[J]. FEMS Microbiol Rev, 2006, 30(3): 382-403.
[15] Rump LV, Beutin L, Fischer M, et al. Characterization of a gne: IS629 O rough:H7Escherichiacolistrain from a hemorrhagic colitis patient[J]. Appl Environ Microbiol, 2010, 76(15): 5290-5291.
[16] Rump LV, Feng PC, Fischer M, et al. Genetic analysis for the lack of expression of the O157 antigen in an O Rough: H7Escherichiacolistrain[J]. Appl Environ Microbiol, 2010, 76(3): 945-957.
[17] Wang Q, Perepelov AV, Wen L, et al. Identification of the two glycosyltransferase genes responsible for the difference betweenEscherichiacoliO107 and O117 O-antigens[J]. Glycobiology, 2012, 22(2): 281-287.
[18] Allison GE, Verma NK. Serotype-converting bacteriophages and O-antigen modification inShigellaflexneri[J]. Trends Microbiol, 2000, 8(1): 17-23.
[19] Sun Q, Knirel YA, Lan R, et al. A novel plasmid-encoded serotype conversion mechanism through addition of phosphoethanolamine to the O-antigen ofShigellaflexneri[J]. PLoS One, 2012, 7(9): e46095.
[20] Rittig MQ, Kaufmarm A, Robins A, et al. Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes[J]. J Leukocyte Biol, 2003, 74(6): 1045-1055.
[21] Samuel G, Reeves P. Biosynthesis of O-antigens: genes and pathways involved in nucleotide sugar precursor synthesis and O-antigen assembly[J]. Carbohydr Res, 2003, 338(23): 2503-2519.
[22] Lguchi A, Lyoda S, Kikuchi T, et al. A complete view of the genetic diversity of theEscherichiacoliO-antigen biosynthesis gene cluster[J]. DNA Res, 2015, 22(1): 101-107.
[23] Orskov F, Orskov I.Escherichiacoliserotyping and disease in man and animals[J]. Can J Microbiol, 1992, 38(7): 699-704.
[24] Wang W, Peng X,Wang Q, et al. Sequence ofEscherichiacoliO-11 O-antigen gene cluster and identification of molecular markers specific to O11[J]. Acta Microbiologica Sinica, 2006, 46(3): 341-346. (in Chinese)
王威,彭霞,王荃,等. 大腸桿菌O11 O-抗原基因簇序列的破譯及特異分子標識的鑒定[J].微生物學(xué)報, 2006, 46(3): 341-346.
[25] Clermont O, Johnson JR, Menard M, et al. Determination ofEscherichiacoliO types by allele-specific polymerase chain reaction: application to the O types involved in human septicemia[J]. Diagn Microbiol Infect Dis, 2007, 57(2): 129-136.
[26] Cheng JS, Wang W, Wang Q, et al. Molecular characterization of O-antigen gene cluster ofEscherichiacoliO23 reference strain and identification of UDP-N-acetylglucosamine 4-epimerase[J]. Acta Microbiologica Sinica, 2006, 46(5): 702-708. (in Chinese)
程劍松,王威,王荃,等. 大腸桿菌O23 O-抗原基因簇序列的破譯及UDP-N-乙酰葡萄糖-C4異構(gòu)酶的鑒定[J].微生物學(xué)報, 2006, 46(5): 702-708.
[27] Liu YH, Yan XH, DebRoy C, et al.EscherichiacoliO-antigen gene clusters of serogroups O62, O68, O131, O140, O142, and O163: DNA sequences and similarity between O62 and O68, and PCR-based serogrouping[J]. Biosensors (Basel), 2015, 5(1): 51-68. DOI: 10.3390/bios5010051
[28] Han WQ, Wang W, Wang L. Characterization ofEscherichiacoliO116 O-antigen gene cluster and development of O-serogroup-specific PCR assay[J]. Chin J Zoonoses, 2007, 23(7): 667-671. (in Chinese)
韓巍青,王威,王磊. 大腸桿菌O116 O-抗原基因簇的破譯及PCR檢測方法的建立[J].中國人獸共患病學(xué)報, 2007, 23(7): 667-671.
[29] DebRoy C, Roberts E, Fratamico PM. Detection of O antigens inEscherichiacoli[J]. Anim Health Res Rev, 2011, 12 (2): 169-185.
[30] DebRoy C, Roberts E, Davis MA, et al. Multiplex polymerase chain reaction assay for detection of nonserotypable Shiga toxin-producingEscherichiacolistrains of serogroup O147[J]. Foodborne Pathog Dis, 2010, 7(11): 1407-1414.
[31] Fratamico PM, Bagi LK, Cray WC, et al. Detection by multiplex real-time polymerase chain reaction assays and isolation of shiga toxin-producingEscherichiacoliserogroups O26, O45, O103, O111, O121, and O145 in groud beef[J]. Foodborne Pathog Dis, 2011, 8(5): 601-607.
[32] Liu YH, Fratamico PM.EscherichiacoliO antigen typing using DNA microarrays[J]. Mol Cellular Probes, 2006, 20(3/4): 239-244.
[33] Wang Q, Wang S, Beutin L, et al. Development of a DNA microarray for detection and serotyping of enterotoxigenicEscherichiacoli[J]. J Clin Microbiol, 2010, 48(6): 2066-2074.
[34] Dunbar SA. Applications of Luminex○RxMAPTMtechnology for rapid, high-throughput multiplexed nucleic acid detection[J]. Clinica Chimica Acta, 2006, 363(1/2): 71-82.
[35] Lin A, Nguyen L, Lee T, et al. Rapid O serogroup identification of the ten most clinically relevant STECs by Luminex microbead-based suspension array[J]. J Microbiol Methods, 2011, 87(1): 105-110.
[36] Coimbra RS, Grimont F, Lenormand P, et al. Identification ofEscherichiacoliO-serogroups by restriction of the amplified O-antigen gene cluster (rfb-RFLP)[J]. Res Microbiol, 2000, 151(8): 639-654. DOI: 10.1016/S0923-2508(00)00134-0
[37] Clotilde LM, Bernar Clay IV, Hartman GL, et al. Microbead-based immunoassay for simultaneous detection of Shiga toxins and isolation ofEscherichiacoliO157 in foods[J]. J Food Prot, 2011, 74(3): 373-379.
[38] Joensen KG, Tetzschner AM, Iguchi A, et al. Rapid and easy in silico serotyping ofEscherichiacoliusing whole genome sequencing (WGS) data[J]. J Clin Microbiol, 2015, 53(8): 2410-2426.
Research progress on identification ofEscherichiacoliO-antigen serogroups
LIU Can-ying, ZHANG Ji-pei, WANG Bing-yun
(DepartmentofVeterinaryMedicine,CollegeofLifeScience,FoshanUniversity,Foshan528231,China)
Escherichiacolicould cause diseases under certain conditions, O-antigens contribute to the virulence ofE.coli. The chemical composition and structure of O-antigen on the surface ofE.colihave high diversity, while specific O-antigen serotype have some connections with certain pathogenicity ofE.coli. Thus, identification of O-antigen serotypes is important for epidemiological studies and control of diseases caused by pathogenicE.coli. The traditional serological typing method is time-consuming, expensive and inaccurate. With the development of science and technology, 196 O-antigen synthesis gene clusters ofE.coliwere sequenced, different O-antigen synthesis gene clusters were compared and analyzed, and molecular markers for specific DNA sequences were designed. PCR method was applied for serotyping O-antigens, including single PCR, multiple PCR, real-time PCR, DNA microarray and microbead-based suspension array. Besides, restriction of the amplified O-antigen gene cluster, microbead-based immunoassay and whole genome sequencing were also used for serotyping O-antigens. These methods would enrich the identification method ofE.coliO-antigen serogroups and make up the deficiency of traditional serological typing method. This review focuses on the research progress about O-antigen synthesis gene cluster and O-antigen serotype identification methods.
Escherichiacoli; O-antigen; serotype
Wang Bing-yun, Email: bywang63@163.com
10.3969/j.issn.1002-2694.2016.010.015
王丙云,Email: bywang63@163.com
佛山科學(xué)技術(shù)學(xué)院,佛山 528231
Q939
A
1002-2694(2016)10-0928-06
2016-04-28;
2016-08-12
廣東普通高校青年創(chuàng)新人才項目(No.2015KQNCX173)
Supported by the project of the Young Creative Talents in Universities in Guangdong Province (No. 2015KQNCX173)
