高溫α-淀粉酶高表達的研究
2010-01-12 09:08:10黃春敏王正祥
微生物學雜志 2010年5期
黃春敏,沈 微,王正祥
(江南大學生物工程學院生物資源與生物能源研究中心和教育部工業生物技術教育部重點實驗室,江蘇無錫 214122)
高溫α-淀粉酶是一種內切型淀粉酶,能隨機水解淀粉質原料及其降解物內部的α-1,4糖苷鍵。因具有反應溫度高、熱穩定性好等特性而被廣泛應用于酒精、白酒、啤酒等食品發酵工業中和紡織業高溫退漿。地衣芽胞桿菌是公認的具有重要工業生產價值的生產菌株,在酶制劑工業中已使用數十年。但目前生產所使用的菌株大多是野生菌株經過傳統遺傳改良獲得的突變株,容易出現平臺效應[1]。在以前的研究已經發現:地衣芽胞桿菌中通過游離質粒增加BLA編碼基因的劑量可以顯著提高BLA的發酵速率和產酶水平[2];通過定向基因整合的方法也可以有效提高BLA的產率[3]。同時在研究中也發現,在特定地衣芽胞桿菌的染色體中,實現BLA編碼基因的整合與基因擴增并不容易發生。為此,本研究構建了一種基于16S rDNA序列為整合位點的am yL重組表達載體,采用原生質體轉化法將此重組質粒轉化入地衣芽胞桿菌CBBD302中,通過16S rDNA序列實現BLA編碼基因的整合與基因擴增,由此獲得了BLA生產水平顯著提高的重組菌。
1 材料與方法
1.1 材料
1.1.1 菌株與質粒 Escherchia coliJM109(CI CIMB0012)、Escherchia coliJM109(pLakr)、B acillus lichenifor m isCBBD302由江南大學中國高校工業微生物資源與信息中心(http://cicim-cu.jiang nan.edu.cn)提供。pBL-amyL[3]、pSKsym-EryR[4]為本實驗室前期構建。
1.1.2 培養基 ①大腸埃希菌和地衣芽胞桿菌的培養基均為LB培養基,需要時加入卡那霉素至50μg/mL或紅霉素至150μg/mL用于重組大腸埃希菌篩選,加入適量紅霉素用于地衣芽胞桿菌的篩選;②地衣芽胞桿菌轉化用培養基:416#培養基、2×S MM、Penassy培養基、S MMP、40%的PEG6000以及再生培養基DM3均參考文獻[5]配置;③發酵培養基:玉米漿40 g/L,硫酸銨30 g/L,乳糖30 g/L;調pH 7.0。發酵試驗在250 mL三角瓶中進行,培養基裝液量為50 mL,200 r/min搖床培養120 h。
1.1.3 主要試劑 DNA限制性內切酶EcoRⅠ、Sm aⅠ、B amHⅠ、B glⅡ、PstⅠ以及卡那霉素、紅霉素、PEG6000,上海生工生物工程技術服務有限公司;T4DNA連接酶,深圳晶美生物工程有限公司;X-Gal、IPTG,寶生物工程有限公司;PCR產物(小量)純化試劑盒、膠回收試劑盒,博大泰克公司;其他試劑藥品皆為國產或進口的分析純和生化試劑。
1.1.4 主要儀器 PCR擴增儀(B IO-RAD公司生產),紫外分光光度計(UN ICO公司生產產品,型號UV-2000),垂直板狀電泳系統(B IO-RAD公司生產),Alpha Innotech凝膠成像系統(美國Alpha Innotech公司生產),臺式高速離心機(美國Sigma公司生產,型號1-15),臺式高速冷凍離心機(美國Sigma公司生產,型號4K15)。
1.2 方法
1.2.1 DNA操作技術 地衣芽胞桿菌染色體DNA的提取,質粒提取、酶切,大腸埃希菌感受態制備、CaCl2法轉化等操作參照文獻[6]進行。DNA片段膠回收、PCR產物純化用博大泰克試劑盒進行。B.lichenifor m is16S rDNA的PCR通用引物L1:AGAGTTTGATCCTGGCTCAG、L2:ACGGCTACCTTGTTACGACTT,PCR反應基本條件:95℃,5 min;94℃,50 s,56℃,90 s,72℃,60 s,35個循環;72℃,10 min。
1.2.2 原生質體的轉化 參照文獻[7]的方法,并根據B.licheniformis CBBD302的特性對原生質體制備和轉化條件進行了優化。挑取單菌落接種于25 mL 416#培養基中,過夜培養。轉接1 mL到新鮮的416#培養基中,振蕩培養約4 h。離心收集菌液,用5 mL S MMP懸浮菌體,加入適量溶菌酶,37℃培養至90%的細胞變為原生質體,1 900 r/min離心10 min。棄上清,將收集的菌體重懸于5 mL S MMP中,隨后依次加入5~10μL質粒、等體積的2×S MM緩沖液和2 mL 40%PEG6000,輕輕混勻后,再加入10 mL的S MMP,2 000 r/min離心10 min。棄上清,再加入500μL S MMP,30℃,100 r/min培養150 min。涂布于含抗性的DM3再生培養基上。
1.2.3 高溫α-淀粉酶酶活的測定以及表達產物的SDS-PAGE分析 高溫α-淀粉酶酶活測定參照國家行業標準QB/T2306-97進行[8]。1個酶活單位定義為在90℃、pH 6.0的條件下1 min液化1 mg淀粉成為糊精所需的酶量。酶活以1 mL發酵液中的高溫α-淀粉酶酶活計算,用U/mL表示。SDS-PAGE分析參照文獻[6]進行。
2 結 果
2.1 整合表達通用性質粒pBli16s-amyL-EryR的構建
以B.licheniform is染色體DNA為模板,用通用引物L1、L2擴增出B.licheniform is的16S rDNA,純化后與經Sm aⅠ酶切的載體pLakr連接,連接產物轉化E.coliJM109感受態細胞,在含有50μg/mL卡那霉素的藍白平板上篩選陽性轉化子,獲得重組質粒pLa-Bli16s。重組質粒pLa-Bli16s經B amHⅠ酶切、pBL-amyL經BglⅡ酶切后膠回收1.88 kb的am yL片段,兩者連接并轉化E.coliJM109,在含有50μg/mL卡那霉素的淀粉平板上篩選陽性轉化子,獲得轉化子pB-li16s-amyL,并提取質粒用PstⅠ酶切驗證。將pB-li16s-amyL用Sm aⅠ酶切,pSKsym-EryR經Sm aⅠ酶切回收1.1 kb的EryR片段,兩者連接并轉化E.coliJM109,在含有50μg/mL紅霉素的淀粉平板篩選陽性轉化子,得到整合表達質粒pBli16samyL-EryR(圖1)。
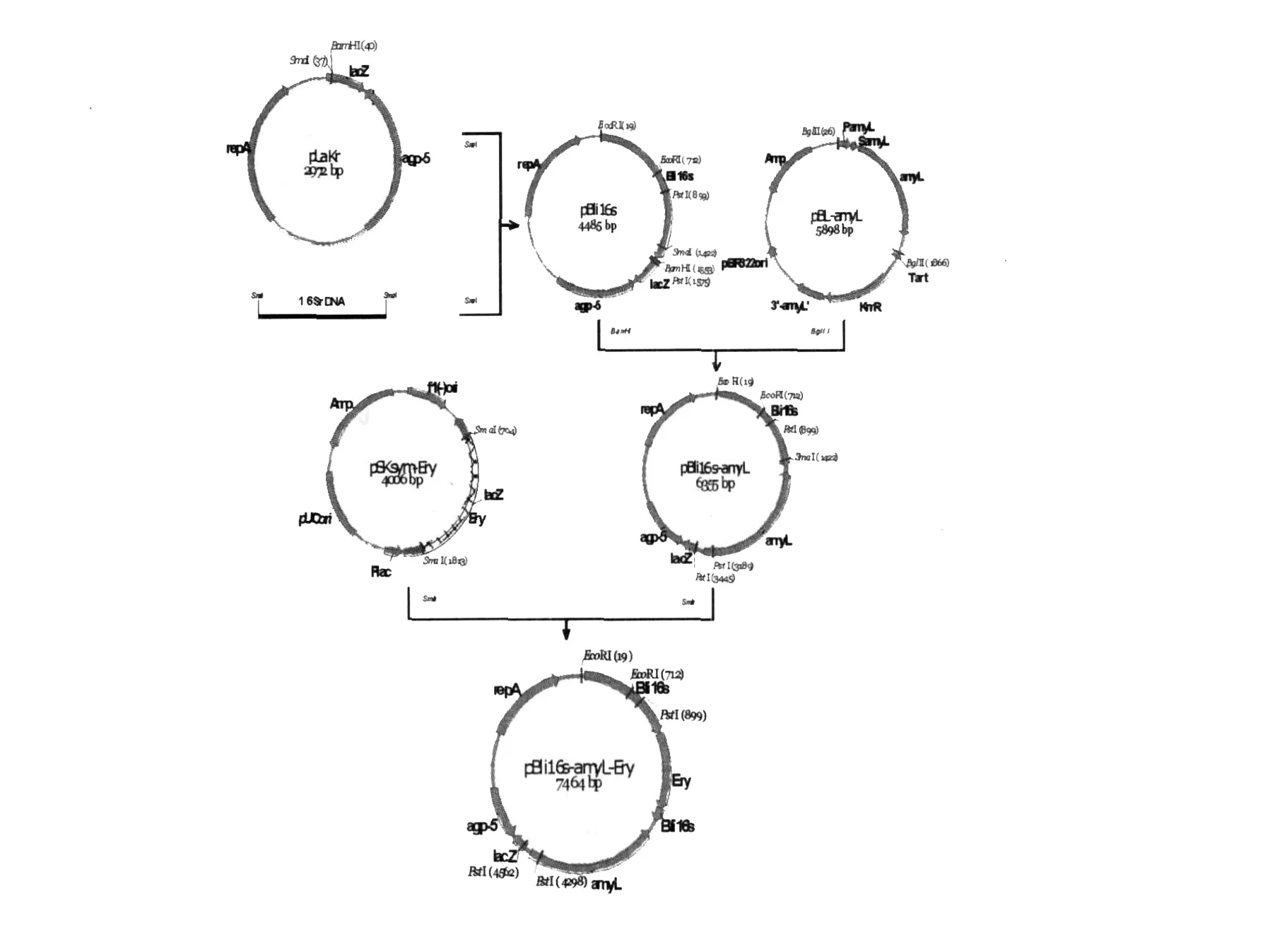
圖1 整合表達通用性質粒pBli16s-amyL-EryR的構建Fig.1 Construction of the common integrated expression plasmid pBli16s-amyL-EryR
2.2 原生質體轉化
用帶有卡那、紅霉素抗性的質粒pBli16samyL-EryR轉化B.licheniform isCBBD302。將所得的轉化子分別點種到無抗和含有卡那霉素、紅霉素雙抗的淀粉平板上,觀察透明圈大小,挑取透明圈較原菌大的菌落分離純化、發酵測酶活,酶活比原菌高10%及以上的即為陽性轉化子。地衣芽胞桿菌原生質體再生困難、限制修飾系統復雜等特殊生理、遺傳性狀,使其分子改良難度大大提高[9],其遺傳轉化至今仍是一大難點。實驗表明,相同條件下選擇對數中后期的菌體有利于原生質體的再生;在制備原生質體時,溶菌酶終濃度為1μg/mL,作用時間為25~30 min,原生質體形成約90%,其轉化子數量最多為20~50 cfu/μgDNA;當原生質體形成率成過高或過低,都不利于原生質體的轉化或再生;在整個轉化過程中動作要輕柔,采用低速離心,防止原生質體破裂。發酵試驗表明,再生平板上長出的轉化子假陽性極高,可能是在原生質體形成和再生過程中細胞壁結構破壞,使抗生素篩選效果大大降低。
2.3 整合表達目的基因拷貝數的提高
本實驗采用地衣芽胞桿菌16S rDNA作為同源臂進行整合,而在地衣芽胞桿菌染色體上16S rDNA的整合位點有幾十個,大大提高了整合轉化的效率,而且可以反復轉化,以提高目的基因的拷貝數。但是由于原生質體轉化效率低、假陽性高,因而本實驗采用在一定范圍內提高液體培養基中抗生素的使用濃度,增加新生子代染色體中同源序列不等位交換,從而增加目的基因在宿主染色體上的基因拷貝數(圖2)。在原生質體轉化獲得的陽性轉化子中,隨機挑取20株在含不同濃度紅霉素的LB培養基上進行培養與誘導,并轉接發酵培養基。酶活測定結果如表1所示。
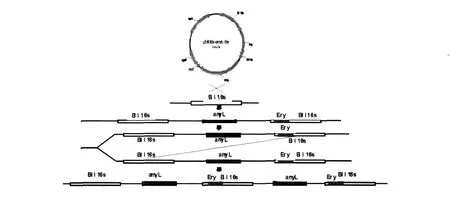
圖2 α-高溫淀粉酶在染色體上的擴增模式Fig.2 The multiplication mode of thermophilicα-amylase in Chromosomes
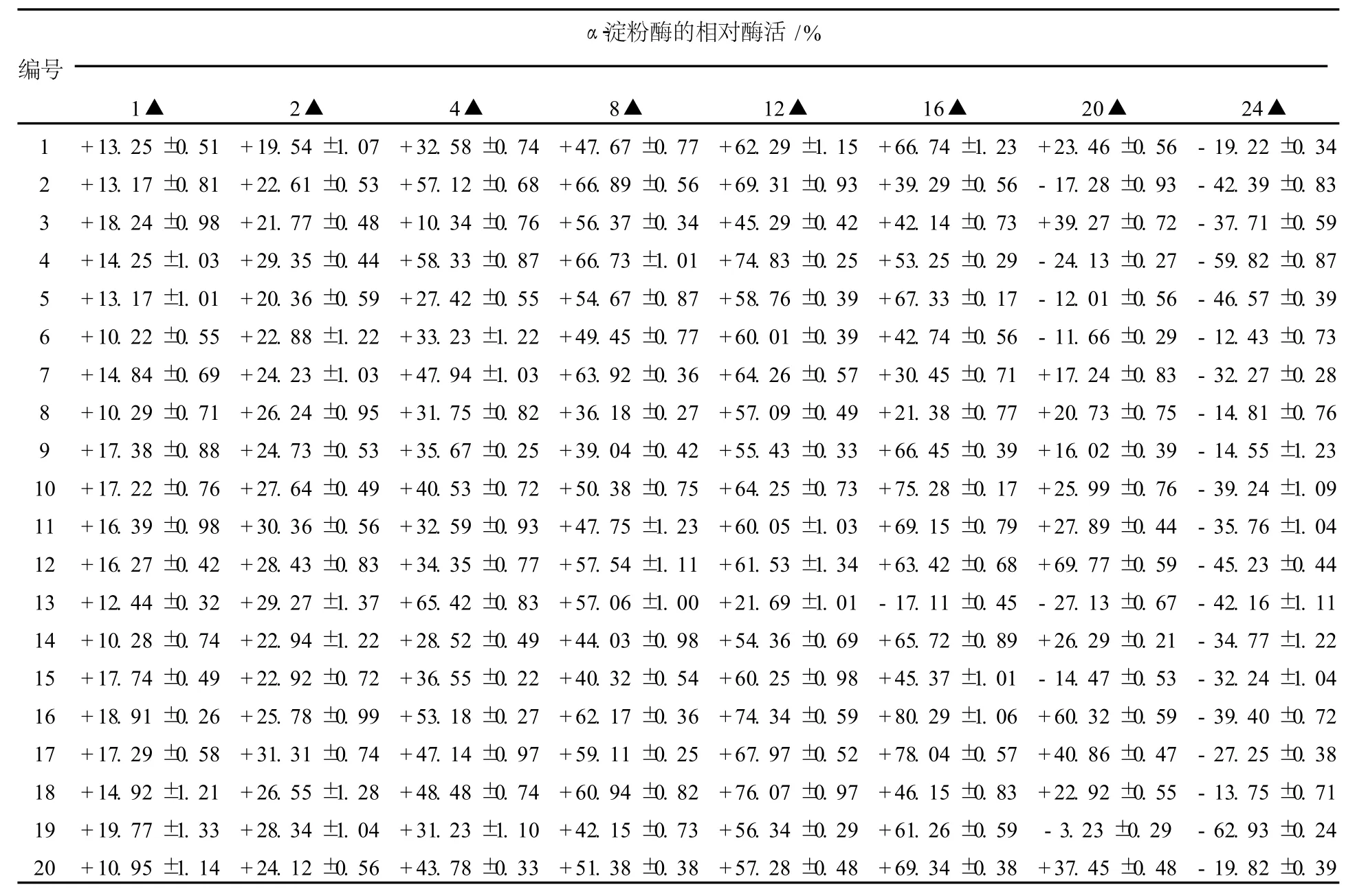
表1 轉化子在不同濃度紅霉素誘導下的α-淀粉酶相對活力Table 1 Relativeα-amylase activity of transformations in induction of different concentrations of erythromycin
由表1可知:不同的轉化子對抗性的敏感性不同,在不同濃度紅霉素的誘導下,轉化子酶活提高了56.37%~80.29%。在低濃度下,酶活隨著抗性增加而非線性提高,這是因為淀粉酶基因胞外分泌表達受到轉錄調節、翻譯效率、肽鏈折疊等諸多因素影響;紅霉素濃度過高,抑制轉化子生長或產生不可逆的變化,使其酶活大大降低。
2.4 表達產物的SDS-PAGE分析
取酶活最高的發酵液與原菌做SDS-PAGE分析表明,目的條帶大小與報道的地衣芽胞桿菌高溫α-淀粉酶分子量55 ku左右相一致,胞外產BLA水平明顯提高(圖3)。由于BLA的大量分泌表達,胞外其他蛋白分泌量也發生了變化。
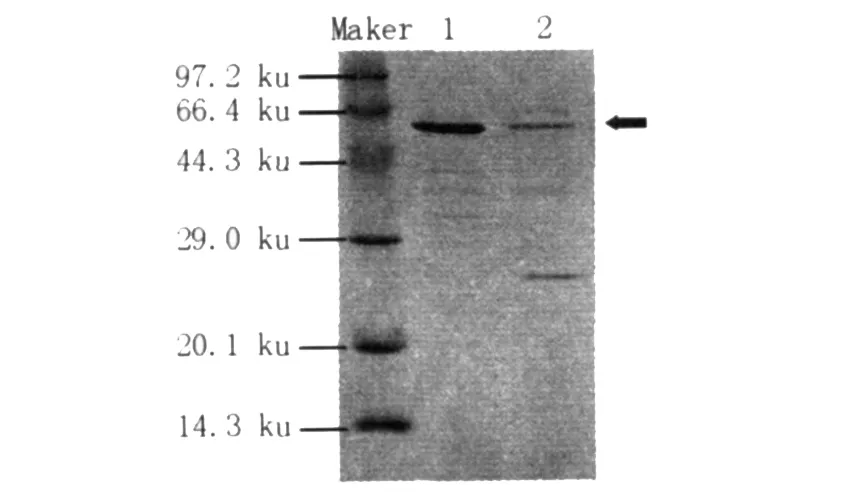
圖3 發酵液的SDS-PAGE分析Fig.3 SDS-PAGE profile of the proteins in broth
3 討 論
通過基因工程手段提高淀粉酶分泌的方法主要有強化啟動子功能、提高基因拷貝數等。Paval等[10]通過提高α-淀粉酶基因拷貝數,使枯草芽胞桿菌野生菌株α-淀粉酶分泌能力提高了2 500倍;牛丹丹等[11]通過多拷貝游離質粒和染色體整合分別將大腸埃希菌、地衣芽胞桿菌分泌的β-甘露聚糖酶活提高了15~18倍和5~7倍;Waldeck J等[12]通過多拷貝游離質粒載體表達異源淀粉酶基因發現突變株表達異源淀粉酶的水平明顯提高,同時細胞自身表達的蛋白酶活力也顯著提高。實驗證明提高酶編碼基因拷貝數是提高其合成與分泌的最有效方法之一。從理論上講,抗性濃度增加,目的基因的拷貝數增加,酶活會相應提高,但是目的基因的分泌表達是個極為復雜的過程,受到操縱子調控、細胞監控、能量再生、分泌調控等因素限制,因此找到不同轉化子的最合適的抗性是實現目的基因高效表達的關鍵。本實驗正是試圖通過改變培養基中抗性使用濃度,篩選出目的基因的高效表達的轉化子。最終轉化子在其合適的抗性濃度下酶活都得到了較大的提高,為56.37%~80.29%。此外,通過SDS-PAGE分析表明地衣芽胞桿菌胞外分泌表達量可能存在極限。
[1] 潘風光,柳增善,劉海學,等.地衣芽胞桿菌耐高溫α-淀粉酶基因工程菌的研究進展[J].內蒙古民族大學學報,2004,35(1):42-44.
[2] NiuDD,Zuo ZR,Shi GY,et al.High yield recombinant thermostable alpha-amylase production using an improved Bacillus licheniform issystem[J].Microbiol Cell Factories,2009,8(1):58-63.
[3] 牛丹丹,石貴陽,王正祥.通過同源介導α-淀粉酶基因擴增改良地衣芽胞桿菌α-淀粉酶生產菌株[J].生物工程學報,2009,3:375-380.
[4] 劉大偉,牛丹丹,張梁,等.便于抗性基因分離和分子重組的系列質粒的構建[J].微生物學通報,2007,34(5):926-928.
[5] Bianca W,Jens W,Susanne W,et al.Generation of readily transformable Bacillus licheniform ismutants[J].ApplMicrobiol Biotechnol,2008,78:181-188.
[6] 奧斯伯FM,金斯頓RE,塞德曼JG,等.精編分子生物學實驗指南[M].北京:科學出版社,2008:42-50.
[7] Chang S,Cohen S.High frequency transformation ofBacillus subtilisprotoplasts by plasmid DNA[J].Mol Gen Genet,1979,168:111-115.
[8] 姜錫瑞.段鋼.酶制劑實用技術手冊[M].北京:中國輕工業出版社,2002:396-403.
[9] Er win HD,Dobek W,Ludolf GB,et al.Novel methods for genetic transformation of natural Bacillus subtilisisolates used to study the regulation of the mycosubtilin and surfactin synthetases[J].Appl Environ Microbiol,2007,73:3490-3496.
[10] Palva I.Molecular cloning of alpha-amylase gene from Bacillus amyloliquefaciensand its expression in B.subtilis[J].Gene,1982,19:81-87.
[11] Waldeck J,Rammes H M,Wieland S,et al.Targeted deletion of genes encoding extracellular enzymes in Bacillus licheniform is and the impact on the secretion capability[J].J Biotechnol,2007,130:124-132.
[12] Dandan Niu,Zheng-Xiang.Wang Development of a pair of bifunctional expression vectors for Escherichia coliand Bacillus lichenifor m is[J].J Ind Microbiol Biotechnol,2007,34:357-362.
