硫甙葡萄糖苷降解研究進展
2010-03-22 03:50:48繆文華潘思軼
食品科學 2010年9期
關鍵詞:研究
李 晨,薛 峰,繆文華,潘思軼*
(華中農業大學食品科學技術學院,湖北 武漢 430070)
硫甙葡萄糖苷降解研究進展
李 晨,薛 峰,繆文華,潘思軼*
(華中農業大學食品科學技術學院,湖北 武漢 430070)
通過硫苷生物合成、降解途徑、降解產物分析方法和降解產物功能作用對十字花科植物中次級代謝產物硫甙葡萄糖苷降解情況進行綜述。硫苷生物合成以氨基酸為底物經過3個步驟完成;硫苷降解存在酶降解、熱降解、化學降解;硫苷降解產物分析方法主要采用化學分析和色譜分析;硫苷降解產物既有抗癌的積極作用又有抗營養化的消極作用。
硫苷;降解途徑;分析方法;功能作用
硫甙葡萄糖苷(glucosinlates,以下簡稱硫苷)是十字花科植物的次級代謝產物,位于植物液泡內[1]。它是一組含硫化合物的總稱,大部分經由植物體內的α-氨基酸的生物合成得到,結構包括β-D-硫葡糖基,磺酸肟,以及由氨基酸衍生而成的支鏈。根據支鏈種類的不同,可將硫苷分為脂肪族、芳香族和吲哚型3大類[2]。
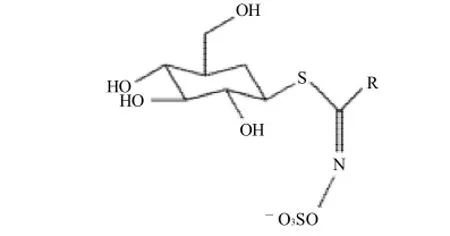
圖1 硫甙葡萄糖苷結構[3]Fig.1 Structure of glucosinolates
當植物組織遭受外界損傷時,位于液泡中的硫苷就會與分布在芥子酶細胞質中的黑芥子苷酶相結合,發生酶解反應,生成具有熱辣氣味的風味物質,主要的3大類風味物質包括異硫氰酸酯、硫氰酸鹽和腈[4]。除酶解反應之外,硫苷還會發生非酶解反應,包括熱降解和化學降解等。根據降解機制或降解條件的不同,生成不同種類的風味物質。
硫苷降解產物由于其特殊的氣味和生化特性,尤其由于其抗癌和植物防御功能[5],從上世紀50年代開始,就引起了研究者們的廣泛關注。到目前為止,已有多篇關于硫苷的降解途徑和各種降解產物的報道,本文對硫苷的降解途徑和降解產物進行綜述,旨在為其降解機制的進一步研究提供參考。
1 硫甙葡萄糖苷的生物合成
硫苷的生物合成可分為3個步驟:首先是氨基酸通過轉氨作用產生α-酮酸,經過異構化使鏈延長形成長鏈酮酸,再通過轉氨作用形成長鏈氨基酸;第2步為葡萄糖苷核心部分的生物合成,即細胞中的氧化酶系將長
鏈氨基酸氧化為不同形式的肟,再由谷胱甘肽轉移酶和CS裂解酶催化肟產生硫甙肟,最后硫甙肟產生的脫硫葡萄糖苷經硫甙轉移酶的催化,產生硫甙葡萄糖苷;第3步為側鏈的修飾,側鏈修飾情況錯綜復雜,不同的修飾途徑可能產生不同硫苷,也可能導致同種產物,因此在側鏈修飾的研究上,還存有很大的空間[6]。
影響硫苷種類和含量的原因有:基因和環境狀況;蔬菜烹飪過程中的工藝影響;包裝和貯藏條件等。由于硫苷是十字花科植物中風味成分的前體物質,其種類和含量將直接決定其降解后生成的風味成分的種類和含量[7]。
2 硫甙葡萄糖苷的降解途徑
硫苷可以通過3種途徑發生降解,分別是酶降解、熱降解和化學降解。發生降解時,硫苷產生等物質的量的葡萄糖、糖苷配基以及硫酸鹽。糖苷配基不穩定,發生進一步降解,生成硫氰酸鹽、腈、異硫氰酸鹽、唑烷以及環硫腈等。其中最主要的具有沖辣氣味的風味成分是異硫氰酸酯,它通過Lossen重排產生,能夠使真菌或昆蟲對植物的侵犯起到防御作用[8]。硫苷的降解產物隨著硫苷結構以及反應條件的變化而變化。
2.1 酶降解
Lüthy等[9]于1984年提出了“芥子油彈”的模型結構。硫苷位于植物細胞的液泡內,能促使硫苷降解的黑芥子苷酶則附著于細胞內膜上,當植物受到損傷,細胞遭到破壞,液泡內的硫苷就會與黑芥子酶相遇,發生降解反應。
在植物中,酶降解的特征是植物體內的抗壞血酸對黑芥子酶活性的控制。Grob等[10]對辣根根部組織進行了檢測,發現有99.5%的抗壞血酸都位于液泡中。硫苷、抗壞血酸、黑芥子酶在植物中有3種分布方式:分別位于不同細胞;位于同一細胞的不同區間;以非活化的形式存在于同一細胞的同一區間。由于高濃度的抗壞血酸抑制酶活性,而低濃度增強酶活[11],因此,當植物未受損傷時,黑芥子酶活性受到高濃度抗壞血酸的抑制,不與硫苷發生反應;而遭受破壞時,液泡膜受損,抗壞血酸會因為細胞液的稀釋而降低濃度,使酶解反應得以發生,因此硫苷、抗壞血酸和黑芥子酶位于同一細胞的同一區間是完全有可能的。
某些昆蟲和微生物體內由于也含有黑芥子酶,能酶解硫苷。硫苷的水解產物尤其是異硫氰酸鹽,對食草昆蟲有毒副作用。為適應這種植物,昆蟲逐漸演化出生化適應性。有不少研究者們從蚜蟲體內檢測到了黑芥子酶。其中Francis等[12]通過硫酸銨分餾、DEAE和凝膠過濾色譜3個步驟從甘藍蚜體內分離出了高純度的蚜蟲黑芥子酶,并利用這種黑芥子酶研究了其對黑芥子糖苷等8種硫苷純品的降解作用及產生的不同降解產物。微生物也會顯示出黑芥子酶活性:人體內腸道細菌含有黑芥子酶,能夠將硫苷在腸道系統內消化[13];真菌如黑曲霉中也被檢測出具有黑芥子酶活性[14]。
硫苷酶酶解的降解產物多樣,酶解最初產生硫甙肟-O-磺酸鹽,通過Lossen重排釋放硫酸鹽后會產生異硫氰酸鹽、硫氰酸鹽、腈和重要的硫化物。不同的降解條件決定了產物的不同。在中性條件下,通常會產生異硫氰酸鹽和硫酮類化合物;pH值較低時,則產生腈和硫氰酸鹽[15]。自然界中能通過酶解過程產生硫氰酸鹽的硫苷只有3種:烯丙基硫苷、苯甲基硫苷和4-丁基硫苷。姜子濤等[16]研究了辣根中酶解反應進行的水浴溫度、時間、pH值和抗壞血酸添加量對酶解反應的影響,得到了最優酶解條件,并檢測出辣根酶解后9種風味成分,其中烯丙基異硫氰酸酯、3-丁烯基異硫氰酸酯、2-戊基異硫氰酸酯和β-苯乙基異硫氰酸酯是主要成分。此外,Rakariyatham等[17]發現曲霉NR-4201能夠對芥末籽餅中的硫苷進行生物降解,固體培養下未檢測出硫苷降解產物,而液體培養下檢測出烯丙基腈的存在。感染了蕪菁花葉病毒的快速增值野生蕓薹屬植物,也檢測出兩種主要的降解產物:3-丁烯基異硫氰酸酯和1-腈基-3,4-環硫丁烷,可能是由此植物體內最豐富的3-丁烯基硫苷轉變而來[18]。
2.2 熱降解
研究熱降解常用的方法為建立模型分析,但由于植物體內的其他因素如金屬離子等會對分析過程產生影響,因此分析數據時就要考慮熱之外的因素。十字花科蔬菜制品在成為商品之前,加熱操作是不可避免的。而在蔬菜中硫苷發生熱降解之時,其降解還有可能包括酶降解和熱介質浸提兩種,然而在研究十字花科植物熱降解的很多情況下,并未將這兩種因素分開考慮[19]。
MacLeod等[20]首次證明異硫氰酸鹽可以由非酶解反應得到,他們采用控制氣相色譜柱溫的方式,發現當柱溫達到200℃時,2-丙烯基硫苷會分解成為44%腈,32%異硫氰酸鹽,而苯甲基硫苷則降解為63%腈和13%的異硫氰酸鹽。亦有研究發現(R)-2-羥基-3-烯基硫苷在100℃加熱3h會完全分解,吲哚硫苷也在熱降解條件下會大量分解,并產生異硫氰酸鹽和吲哚乙酸腈等風味產物[21]。Oerlemans等[22]首次將紅球甘藍中的黑芥子酶在實驗進行前滅活,以排除酶降解的干擾,專門研究熱降解對其中硫苷的影響,發現在110℃溫度以下,吲哚類硫苷的降解速率大于脂肪族硫苷,并且將熱燙、蒸煮、罐裝3種熱處理條件對硫苷的熱降解進行了模擬,研究表明,硫苷的熱降解程度在罐裝條件下最大,蒸煮次之,最小為熱燙。
2.3 化學降解
硫苷還能夠被酸、堿以及鹽溶液降解。在酸溶液中,硫苷的一般降解產物為相應羧酸、D-葡萄糖、羥胺離子和氫硫酸鹽,也可能產生新的硫苷[23]。而在堿溶液中,硫苷的降解產物則比較復雜,有硫氰酸鹽、硫酸鹽、硫化氫、腈、烯丙基腈、吲哚等。Fensick等[24]從2-丙烯基硫苷的NaOH水溶液中得到了硫代葡萄糖,Friis等[25]在2-丙烯基硫苷和苯甲基硫苷中加入2mol/L NaOH溶液,產生了相應氨基酸,并猜測其可能是由于Neber重排產生。不僅是在酸堿溶液,在鹽溶液如2-丙烯基硫苷在BaCl2中,會產生BaSO4、S、KCl和丙烯腈,而在AgNO3水溶液中,有H2S存在時,生成丙烯腈和AgS;有Na2S2O3存在時,則生成異硫氰丙烯和AgS。硫苷的化學降解涉及較少,且降解機制還未被研究,因此,在這方面存有較大的空間供以后深入探討。
2.4 硫苷降解影響因素
影響硫苷降解的因素有很多。首先是蔬菜的制作過程中工藝過程的影響。工藝過程可能會使黑芥子酶部分失活、使硫苷或硫苷降解產物發生熱降解、酶解硫苷或減少能增大酶活的物質(如抗壞血酸、Fe2+)等。當代研究者們的研究也主要集中在工藝因素對硫苷降解的作用上,主要的工藝因素有打碎、煮制、冷凍、脫水、發酵以及涼菜烹調工藝例如添加劑、調味料的添加等[26]。Volden等[27]研究了3種工藝過程:熱燙、煮沸和蒸氣加工對花椰菜硫苷含量和其他抗氧化參數的影響,得出蒸氣加工對硫苷的影響最小,只降解了19%,其次為熱燙,42%,影響最大的是煮沸,55%,且吲哚類比脂肪族硫苷耐熱性強。Wenngerg等[28]研究了熱燙以及乙酸處理對兩個不同品種白菜中碳水化合物和硫苷組成的影響,發現熱燙會使兩種白菜內的硫苷含量都降低,而乙酸只降低其中一種白菜中的硫苷含量,這項研究還對單個硫苷的含量進行了分析,使工藝因素的作用程度更加具體化。Martinez-Villaluenga等[29]研究了白菜發酵條件下硫苷的變化,發現硫苷含量急劇降低,而且在發酵7d后,僅有微量單個硫苷被檢測出。
其次是蔬菜前處理方式的不同,Ildikó等[30]研究了微波熱處理對白芥末營養和膠狀化學性質的影響,其研究表明,處理過后的硫苷含量保持不變,而釋放出的異硫氰酸鹽的含量有較為顯著的降低,從這個角度說明了黑芥子酶的活性被抑制。van Eylen等[31]研究了壓力/溫度處理對花椰菜內源黑芥子酶的穩定性和活性以及細胞可滲透性的影響。表明壓力/溫度對黑芥子酶活性的影響有先增大后降低的趨勢,且在45℃、100MPa達到最大,因此用壓力/溫度處理食品可以先增大酶活使硫苷降解產生風味物質,再降低酶活抑制反應以保持食品的感官性質。這項研究也為溫度/壓力對于風味物質保存的應用提供了理論依據。
再次在反應條件的不同上,Uda等[32]從Tanaka種子中提取黑芥子酶,用此酶水解黑芥子糖苷,并研究了pH值和Fe2+兩因素對硫苷降解作用的影響,發現在pH4.5和5.5時,Fe2+能嚴重抑制異硫氰酸鹽的產生,這種抑制作用在pH6.5時降低,并與pH7.5時消失。又通過此酶水解金龜草種子中提取出的硫苷,驗證了Fe2+不影響酶解過程中葡萄糖的釋放,而只影響風味物質的生成。Pan等[33]也研究了金屬離子對辣根風味形成的影響,另外還研究了p H值、水浴溫度、時間、濕度、抗壞血酸、干燥方法等諸多因素對辣根風味成分中的烯丙基異硫氰酸酯和β-苯乙基異硫氰酸酯含量的影響,以及β-環糊精、變性淀粉和大豆蛋白對風味物質的保留作用。陳虹霞等[34]以辣根為原料,研究硫代葡萄糖苷水解為異硫氰酸酯的轉化率。結果表明,在溫度40℃、pH7、料液比1∶20(m/V)的條件下,水中加入VC 0.2%(以辣根粉計),水解2h,水蒸氣蒸餾法得到的異硫氰酸酯質量分數最高。Jia等[35]又發現完全氣調儲藏比部分氣調儲藏更能保持花椰菜內硫苷的含量,進而也更能保持蔬菜的風味成分。
由此可見影響硫苷降解產物因素的多樣化。除此之外,張清峰等[36]研究了硫苷降解產物異硫氰酸酯的水解穩定性,并對其水解機理做出推測,將硫苷的降解又延伸到了深一層次。由于很多研究都沒有對降解后產生的風味物質變化的原因作出闡述,因此就需要以后的研究進行拓展。
3 硫甙葡萄糖苷降解產物的分析方法
檢測硫苷降解產物的化學方法很多,張清峰等[37]在化學方法分析硫苷降解產物上做出了很多研究。嗎啉滴定法、哌啶滴定法、氯胺-T間接氧化法等都是很成功的檢測方法。但為獲得較為精確的風味組成,一般來說,主要利用的分析方法是用溶劑將其從植物組織中提取出來,再利用色譜技術進行分析,最常用的是GC-MS聯用。Jiang等[16]利用哌啶滴定法測辣根水解后的異硫氰酸鹽含量,再將辣根組織于65℃水解120min,水解液經過水蒸氣蒸餾后用乙醚提取,用GC-MS分析風味物質的具體組成。Blaevi等[38]則利用Clevenger-type裝置從辣根的葉、莖中提取硫苷降解產物和其他揮發性成分后,采用GC加GC-MS的方法進行性質分析。而若要獲得純度較高的降解產物,則需要色譜分離技術的應用。Vaughn等[39]研究了蔥芥中的風味成分,采用有機溶劑旋蒸提取可揮發性成分,過親脂性Sephadex LH-20柱進一步純化,得到的組分經GC-MS測定,檢測出烯丙基異硫氰酸酯、苯甲基異硫氰酸酯和2,3-環硫丙基腈。Vaughn等[40]創新的運用不同pH值的緩沖液,從
白芥末中提取出純度高于97%的硫苷水解物,再進行GC-MS分析,開創了不用色譜方法而直接用化學方法提取高純度硫苷水解物的先河。
應用色譜方法檢測十字花科中風味物質上,LC-MS也是較為常用的一種。Song等[41]利用氨水將異硫氰酸鹽類轉化為相應的脲硫衍生物,再用LC-MS/MS方法檢測異硫氰酸鹽的組分及含量。隨著新型檢測食品風味物質的方法——固相微萃取法(SPME)的興起,越來越多的風味研究用到這項技術,肖華志等[42]利用固相微萃取/氣相色譜-質譜(SPME/GC-MS)聯用技術檢測出芥末油中6種風味成分,青芥辣中12種風味成分,以及沖菜中的17種風味組成。Zhao等[43]也利用SPME/GC-MS對雪里蕻腌菜發酵過程中的揮發性組分進行了分析,檢測出87種組分,其中異硫氰酸鹽有5種,腈類有4種。用SPME/ GC-MS分析方法簡單、靈敏度高,且不用對分析物中的揮發性物質進行提取,進一步減少了實驗的耗時和結果誤差。
除以上較為常用的風味成分分析方法之外,還有Likens-Nickerson裝置從羅馬花椰菜中提取風味物質[44],或采用β-環糊精包埋風味物質再進行GC-MS檢測等方法[45],只限于提取方法的改善,分析組分時所用的儀器手段沒有太大變化。但Karcher等[46]利用毛細管電泳法測定硫苷降解產物,則是對風味成分進行分析的另辟蹊徑。
4 硫甙葡萄糖苷降解產物的功能作用
硫苷降解產物因其功能作用的復雜而受到研究者們的廣泛關注。硫苷降解產物一方面被認為能夠有效抑制癌細胞的產生。其主要機制一是作為阻遏因子抑制階段Ⅰ和誘導階段Ⅱ生物轉化酶的活性:其中階段Ⅰ酶不僅能代謝某些毒素,還能激活某些致癌物質,因而可能導致致癌程度的增加。而階段Ⅱ酶能催化階段Ⅰ酶生成的一系列水解物或氧化還原產物解毒,使這些產物發生結合反應進而生成較易分泌的分子;一些階段Ⅱ酶還能通過氧化還原反應破壞具有高度活化中央結構的致癌劑,進而抑制癌癥[47];機制二是硫苷能作為抑制因子誘導癌細胞程序化死亡,由于癌細胞都具有基因毒性,修補基因組損傷的難度較大,利用癌細胞具有高代謝速率而較易程序化死亡的特點,可以誘導癌細胞程序化死亡以消滅癌細胞[48]。
另一方面,硫苷降解產物還在高等動物中表現出一系列毒性和抗營養化效應,其中研究最透徹的是對甲狀腺的副作用。含有β-OH的硫苷能降解生成異硫氰酸鹽和唑烷-2-硫,這兩種降解產物都能導致甲狀腺腫大。其中異硫氰酸鹽會競爭碘離子,因此可在豬或鼠的飼料中增加碘離子濃度以減少甲狀腺腫大的現象;而5-乙烯基唑烷-2-硫則直接抑制甲狀腺合成。硫苷降解產物能通過不同的方式作用于動物或人體的甲狀腺造成危害。
硫苷降解產物還有不少其他功能。Brabban等[49]研究表明硫苷對微生物的生長無任何影響,而它的水解產物則對微生物的生長具有抑制作用;Depree等[50]指出山葵中的異硫氰酸鹽有較強的抗細菌、真菌作用,從山葵葉、葉柄、根等中提取的精油也能抑制血小板凝集作用;從洋蔥中提取的物質也被證明具有抗哮喘效應。諸多研究都表明硫苷水解產物有著很強的功能作用,若能廣泛開發,將極大程度的造福于人類。
隨著人們消費水平的提高和消費方式的改變,追求辛辣刺激又健康的飲食方式逐漸被人們所喜愛,很多以十字花科蔬菜為主要原料的具有強烈辛辣沖味的調味品如辣根醬、芥末油等在近年來越來越廣泛的進入人們的日常飲食[51]。硫苷降解產物,是這一辛辣沖味的主要來源,研究它的降解途徑、降解機理和降解產物,必將積極推動此項研究在工業生產當中的應用,并在很大程度上推進我國在辛辣沖味調味品上落后于他國、依賴進口的劣勢,滿足消費者的需求。
[1]錢驊, 趙伯濤, 黃曉穗. 西蘭花中β-硫甙葡萄糖苷總量的測定[J]. 中國野生植物資源, 2008, 27(2)∶ 52-53.
[2]BUROW M, ZHANG Z Y, OBER J A, et al. ESP and ESM1 mediate indol-3-acetonitrile production from indol-3-ylmethyl glucosinolate in Arabidopsis[J]. Phytochemistry, 2008, 69(3)∶ 663-664.
[3]ELFAKIR C, DREUX M. Simultaneous analysis of intact and desulfated glucosinoaltes with a porous graphitized carbon column[J]. Journal of Chromatography A, 1996, 727∶ 71-72.
[4]BONES A M, ROSSITER J T. The myrosinase-glucosinolate system, its organization and biochemistry[J]. Physiologia Plantarum, 1996, 97∶204-205.
[5]KLIEBENSTEIN D J, KROYMANN J, MITCHELL-OLDS T. The glucosinolate-myrosinase system in an ecological and evolutionary context [J]. Current Opinion in Plant Biology, 2005, 8∶ 264-271.
[6]MITHEN R F, DEKKER M, VERKERK R, et al. The nutritional significance, biosynthesis and bioavailability of glucosinolates in human foods[J]. Journal of Science and Food Agricultural, 2000, 80(7)∶973-974.
[7]DEKKER M, VERKERK R, JONGEN W M F. Predictive modeling of health aspects in the food production chain∶ a case study on glucosinolates in cabbage[J]. Trends in Food Sci Tech, 2000, 11∶ 177-178.
[8]FRANCIS F, LOGNAY G, WATHELET J P, et al. Effects of allelochemicals from first (Brassicaceae) and second (Myzus persicae and Brevicoryne brassicae) trophic levels on Adalia bipunctata[J]. Journal of Chemical Ecology, 2001, 27(2)∶ 243-244.
[9]LüTHY B, MATILE P H. The mustard oil bomb∶ rectified analysis of the subcellular organization of the myrosinase system[J]. Biochem. Physiol Pflanz, 1984, 179∶ 5-12.
[10]GROB K, MATILE P H. Compartmentation of ascorbic acid in vacuoles of horseradish root cells. Note on vacular peroxidase[J]. Z Pflanzenphysiol, 1980, 98∶ 235-243.
[11]BONES A M, SLUPPHAUG G. Purification, characterization and par-
tial amino acid sequencing of β-thioglucosidase from Brassica napus L.[J]. Journal of Plant Physiol, 1989, 134∶ 722-729.
[12]FRANCIS F, LOGNAY G, WATHELET J P, et al. Characterisation of aphid myrosinase and degradation studies of glucosinolates[J]. Archives of Insect Biochemistry and Physiology, 2002, 50∶ 173-182.
[13]GETAHUN S M, CHUNG F L. Conversion of glucosinolates to isothiocyanates in humans after ingestion of cooked watercress[J]. Cancer Epi Bio Pre, 1999, 8∶ 447-451.
[14]SMITS J P, KNOL W, BOL J. Glucosinolate degradation by Aspergillus clavatus and Fusarium oxysporum in liquid and solid-state fermentation [J]. Applied Microbiology and Biotechnology, 1993, 38∶ 696-697.
[15]SHEN L Q, SU G Y, WANG X Y. Endogenous and exogenous enzymolysis of vegetable-sourced glucosinolates and in uencing factors [J]. Food Chemistry, 2009, 8(3)∶ 1-2.
[16]JIANG Zitao, LI Rong, YU J C. Pungent components from thioglucosides in Armoracia rusticana grown in China, obtained by enzymatic hydrolysis [J]. Food Technol Biotechnol, 2006, 44(1)∶ 41-45.
[17]RAKARIYATHAM N, SAKORN P. Biodegration of glucosinolates in brown mustard seed meal (Brassica juncea) by Aspergillus sp.NR-4201 in liquid and solid-state cultures[J]. Biodegradation, 2002, 13∶ 395-399.
[18]SHATTUCK V I. Glucosinolates and glucosinolate degradation in seeds from turnip mosaic virus-infected rapid cycle Brassica campestris L. plants[J]. Journal of Experimental Botany, 1993, 44(5)∶ 963-970.
[19]VOLDEN J, BORGE G I A, BENGTSSON G B. Effect of thermal treatment on glucosinolates and antioxidant-related parameters in red cabbage (Brassica oleracea L. ssp. capitata f. rubra)[J]. Food Chemistry, 2008, 109∶ 595-596.
[20]MACLEOD A J, ROSSITER J T. Non-enzymic degradation of 2-hydroxybut-3-enylglucosinolate (progoitrin)[J].Phytochemistry, 1986, 25∶ 855-858.
[21]BONES A M, ROSSITER J T. The enzymic and chemically induced decomposition of glucosinolates[J]. Phytochemistry, 2006, 67∶ 1053-1067.
[22]OERLEMANS K, BARRETT D M. Thermal degradation of glucosinolates in red cabbage[J]. Food Chemistry, 2006, 95∶ 19-29.
[23]OLSEN O, SφRENSEN H S. Recent advances in the analysis of glucosinolates[J]. Journal of the American Oil Chemists Society, 1981, 9∶ 858-860.
[24]FENSICK G R, HEANEY R K, GEMLIN R, et al. Glucosinalbin in Brassica napus∶ a re-evaluation[J]. Zeitschrift fuer Pflanzenzuechtung, 1981, 87∶ 254-259.
[25]FRIIS P, LARSEN P O, OLSEN C E. Base-catalyzed neber-type rearrangement of glucosinolates [1-(β-D-glucosylthio)-N-(sulfonatooxy) alkylideneamines][J]. Journal of the Chemical Society∶ Perkin Transactions 1, 1977(6)∶ 661-665.
[26]de VOS R H, BLIJLEVEN W G H. The effect of processing conditions on glucosinolates in cruciferous vegetables[J]. Ubersichtsbericht, 1988, 187∶ 526-529.
[27]VOLDEN J, BORGE G I A, HANSEN M, et al. Processing (blanching, boiling, steaming) effects on the content of glucosinolates and antioxidant-related parameters in cauliflower (Brassica oleracea L. ssp. botrytis) [J]. Food Science and Technology, 2009, 42∶ 63-73.
[28]WENNGERG M, EKVALL J, OLSSON K, et al. Changes in carbohydrate and glucosinolate composition in white cabbage (Brassica oleracea var. capitata) during blanching and treatment with acetic acid[J]. Food Chemistry, 2006, 95∶ 226-236.
[29]MARTINEZ-VILLALUENGA C, PENAS E, FRIAS J, et al. Influence of fermentation conditions on glucosinolates, ascorbigen, and ascorbic acid content in white cabbage (Brassica oleracea var. capitata cv. Taler) cultivated in different seasons[J]. Food Chemistry, 2009, 74(1)∶ 62-67.
[30]ILDIKO S G, KLARA K A, MARIANNA T M, et al. The effect of radio frequency heat treatment on nutritional and colloid-chemical properties of different white mustard (Sinapis alba L.) varieties[J]. Innovative Food Science Emerging Technologies, 2006, 7∶ 74-79.
[31]van EYLEN D, OEY I, HENDRICKX M, et al. Effects of pressure/ temperature treatments on stability and activity of endogenous broccoli (Brassica oleracea L.cv. Italica) myrosinase and on cell permeability[J]. Journal of Food Engineering, 2008, 89∶ 178-186.
[32]UDA Y, KURATA T, ARAKAWA N. Effects of pH and ferrous ion on the degradation of glucosinolates by myrosinase[J]. Agric Biol Chem, 1986, 50(11)∶ 2735-2740.
[33]PAN S Y, HYDAMAKA A, KUO A. Horseradish flavor formation optimization[J]. Transactions of the CSAE, 2003, 19(5)∶ 141-145.
[34]陳虹霞, 王成章, 葉建中, 等. 辣根硫代葡萄糖苷水解工藝比較及其水解產物的結構鑒定[J]. 林產化學與工業, 2009, 29(2)∶ 100-104.
[35]JIA Chengguo, XU Chaojiong, WEI Jia, et al. Effect of modied atmosphere packaging on visual quality and glucosinolates of broccoli orets [J]. Food Chemistry, 2009, 114(1)∶ 28-37.
[36]張清峰, 姜子濤, 張云華. 辣根油中異硫氰酸酯的水解穩定性研究[J]. 食品科學, 2005, 26(11)∶ 125-127.
[37]張清峰, 姜子濤, 董峰光, 等. 二乙胺滴定法測定辣根及芥末制品中異硫氰酸酯含量的研究[J]. 中國調味品, 2005(2)∶ 48-51.
[38]BLAZEVIC I, MASTELIC J. Glucosinolates degradation products and other bound and free volatiles in the leaves and roots of radish(Raphanus sativus L.)[J]. Food Chemistry, 2009, 113∶ 96-102.
[39]VAUGHN S F, BERHOW M A. Allelochemicals isolated from tissues of the invasive weed garlic mustard (Alliaria petiolata)[J]. Journal of Chemical Ecology, 1999, 25 (11)∶ 2495-2502.
[40]VAUGHN S F, BERHOW M A. Glucosinolate hydrolysis products from various plant sources∶ pH effects, isolation, and purification[J]. Industrial Crops and Products, 2005, 21∶ 193-202.
[41]SONG L J, MORRISON J J, BOTTING N P. Analysis of glucosinolates, isothiocyanates, and amine degradation products in vegetable extracts and blood plasma by LC-MS/MS[J]. Analytical Biochemistry, 2005, 347∶ 234-243.
[42]肖華志, 牛麗影, 廖小軍, 等. 芥末油、青芥辣、沖菜的揮發性風味成分的SPME-GC-MS測定[J]. 中國調味品, 2004(6)∶ 44-45.
[43]ZHAO Dayun, TANG Jiang, DING Xiaolin. Analysis of volatile components during potherb mustard (Brassica juncea, Coss.) pickle fermentation using SPME-GC-MS[J]. LWT, 2007, 40∶ 439-447.
[44]VALETTE L, FERNANDEZ X, POULAIN S, et al. Volatile constituents from Romanesco cauliflower[J]. Food Chemistry, 2003, 80∶ 353-358.
[45]YU J C, JIANG Zitao, LIU Hoyan, et al. β-Cyclodextrin epichlorohydrin copolymer as a solid-phase extraction adsorbent for aromatic compounds in water samples[J]. Analytica Chemica Acta, 2003, 477(1)∶ 93-101.
[46]KARCHER A, RASSI Z E. Capillary electrophoresis of glucosinolates and their degradation products[J]. Electrophresis, 1999, 20∶ 3181-3189.
[47]修麗麗, 鈕昆亮. 十字花科植物中的硫甙葡萄糖苷及其降解產物[J].浙江科技學院學報, 2004, 16(3)∶ 189-190.
[48]汪俏梅, ABEL S. 異硫代氰酸鹽的抗癌機理及其相關研究[J]. 細胞生物學雜志, 2002, 24(3)∶ 171-173.
[49]BRABBAN A D, EDWARDS C. The effects of glucosinolates and their hydrolysis products on microbial growth[J]. Journal of Applied Bacteriology, 1995, 79∶ 171-177.
[50]DEPREE J A, HOWARD T M, SAVAGE G P. Flavour and pharmaceutical properties of the volatile sulphur compounds of Wasabi (Wasabia japonica)[J]. Food Research International, 1999, 31(5)∶ 329-337.
[51]牛麗影,肖華志, 胡小松, 等. 沖菜辛辣風味物質的頂空固相微萃取-氣質聯用法測定[J]. 中國食品學報, 2005, 5(3)∶ 122-123.
Research Progress in the Degradation of Glucosinolates
LI Chen,XUE Feng,MIAO Wen-hua,PAN Si-yi*
(College of Food Science and Technology, Huazhong Agricultural University, Wuhan 430070, China)
Biosynthesis, degradation pathways, degradation product analysis, degradation product functions of glucosinolates play important roles in the secondary metabolites in Cruciferae plants, which has been reviewed in this paper. The biosynthesis of glucosinolates was completed using amino acids as the substrates through three steps. The degradation pathways of glucosinolates were classified as enzymatic degradation, thermal degradation and chemical degradation. Chemical and chromatographic analyses were the major strategies for the analysis of glucosinolate degradation products. The degradation products of glucosinolates exhibited a positive role in anti-cancer as well as a negative role in anti-nutrient.
glucosinolates;degradation pathway;degradation product analysis;degradation product function
TS255.1
A
1002-6630(2010)09-0313-05
2009-10-23
李晨(1987—),女,碩士研究生,研究方向為食品科學。E-mail:lichen@webmail.hzau.edu.cn
*通信作者:潘思軼(1964—),男,教授,博士,研究方向為食品生物技術。E-mail:pansiyi@mail.hzau.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
