與灰葡萄孢致病性相關的真菌毒素及胞外酶
2010-07-31 01:22:50劉莎莎陳小龍
浙江農業科學 2010年4期
關鍵詞:植物
劉莎莎,陳小龍
(浙江工業大學生物與環境工程學院發酵工程研究所,浙江杭州 310032)
灰葡萄孢 (Botrytis cinerea)屬于半知菌亞門(Hyphomycetes)絲孢綱 (Deuteromycotina)葡萄孢屬,是一種植物病原真菌,宿主達200余種,其中包括重要的經濟作物葡萄、番茄、草莓等。在植物的生長期、開花期、結果期及運輸儲藏期灰葡萄孢都可以感染作物,所以由灰葡萄孢引起的灰霉病被認為是最嚴重的植物真菌病害之一。每年都有關于灰葡萄孢感染新的作物品種的相關報道,在環境適宜的條件下,特別是溫室培養,灰霉病會大面積爆發,帶來嚴重的經濟損失[1-3]。
目前抗灰霉病的方法主要集中在化學防治和生物防治2方面[4],我國防治灰霉病使用的藥劑主要包括苯丙咪唑類 (如多菌靈),二甲酰亞胺類 (如速克靈)和氨基甲酸酯類 (如乙霉威)殺菌劑,國外也有一些通過控制作物栽培氣候條件和培養抗灰葡萄孢植株來抵抗灰霉病的相關報道[5-7]。由于化學防治容易產生抗性,生物防治和其它舉措還沒有特別行之有效的方法[8-10],所以找到對灰霉病有特效的殺真菌劑是全世界范圍內急需解決的難題。
灰葡萄孢在進入植物組織之前先殺死細胞,然后吸取植物的營養,這個過程中灰葡萄孢會分泌多種有毒的代謝產物和胞外酶。目前國內外對灰葡萄孢致病機理研究的重點都集中在這些致病因子的合成途徑以及作用方式。
1 灰葡萄孢毒素
灰葡萄孢毒素成分種類很多,國外已分離出將近20種化合物,這些毒素中最為關鍵的是二環倍半萜烯botrydial和高取代內酯 botcinolide,其它鑒定出結構的毒素都為這2種物質的前體或衍生物,毒性相對較低。
botrydial最早在1974年被分離鑒定出來,botcinolide最早在1993年被分離鑒定出來,它們的結構式如圖1所示[11-12]。所有已經分離鑒定出的灰葡萄孢毒素中,botrydial的毒性最高,活體實驗結果表明,灰葡萄孢分泌botrydial越多,其對宿主細胞表現出的毒力就越高[13]。目前國內外的研究都集中在botrydial的致病機理以及生物合成途徑方面。
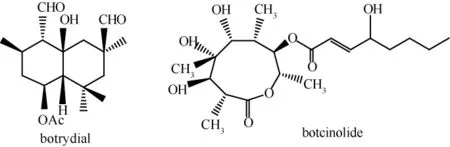
圖1 botrydial和botcinolide的化學結構式
1.1 botrydial和以灰霉二醛為骨架的化合物
很多絲狀真菌的毒力都是萜類化合物引起的,這些化合物幫助植物真菌突破宿主的細胞防御。botrydial是非特異性毒素,可以作用于不同作物和基因型,正是因為這樣灰霉病才能引發大面積的植物病害。Isidro等人發現,被灰葡萄孢侵染的宿主植物出現傷口后,植物組織細胞中就可以檢測到botrydial,但植物本身并不能分泌這種化合物。隨著植物葉片上萎黃程度增高,檢測到botrydial的量也增加,所以灰葡萄孢分泌的二環倍半萜類毒素被認為是灰霉病致病的關鍵因素,這些毒素可以引發宿主植物萎黃病和組織細胞破裂[14]。Colmenares等[15]發現,botrydial的活性具有光依賴性,光照條件下botrydial引發宿主產生病斑的能力比黑暗條件下高出1倍,但其機理還未闡明。
灰葡萄孢在液體培養時除了會產生botrydial外,發酵液中還發現了大量以灰霉二醛為骨架的二環倍半萜類毒素,這些含灰霉二醛骨架化合物的生物活性存在一定差異。通過比較后發現,這些化合物的生物活性與灰霉二醛骨架結構上C-10和C-15原子的氧化態以及C-9上的羥基有直接關系[16]。
1.2 botrydial的生物合成途徑和相關基因
了解botrydial及其相關化合物的合成途徑對灰霉病致病性的研究具有重大意義。最近,合成botrydial的基因簇及其編碼的蛋白序列已經鑒定出來,5個合成botrydial基因簇的開放閱讀框及功能如圖2所示[17]。BcBOT2負責編碼的蛋白是1種倍半萜合酶,將FPP(法呢基焦磷酸)環化生成關鍵的中間產物 PSP(presilphiperfolan-8βol),隨后PSP在3種單加氧酶細胞色素 P450(BcBOT3,BcBOT1,BcBOT4編碼)和乙酰轉移酶 (BcBOT5編碼)的作用下生成 botrydial。botrydial的生物合成途徑如圖 3 所示[18]。

圖2 合成botrydial基因簇的開放閱讀框及功能

圖3 botrydial的生物合成途徑
在5個基因簇中,最重要的是1.2 kb的BcBOT2基因,它負責編碼倍半萜合酶,此酶是botrydial合成途徑中的關鍵酶。敲除BcBOT2基因的灰葡萄孢菌株失去了合成botrydial和相關化合物的能力,菌株毒力基本喪失[18]。PSP轉化成botrydial的過程中涉及到多種以灰霉二醛為骨架的化合物,這些化合物在整個途徑中起到中間體的作用。
1.3 灰葡萄孢毒素的應用
灰葡萄孢產生的毒素會引發植物枯萎死亡,特別是大部分雙子葉植物。botrydial在1.0 μg·mL-1的濃度下就可以使植物葉片表現典型的癥狀,botcinolide在濃度為10-3mol·L-1時對某些單子葉和雙子葉植物就有很強的抑制或殺除作用,但這2種化合物及其相關化合物對小鼠的毒性都不高[13],所以灰葡萄孢毒素可以作為生物除草劑進行開發利用,國內外對灰葡萄孢毒素的研究也都集中在除草劑方向。Cutler在1995年申請了botcinolide應用為除草劑的專利,鄭蒙等[19]對灰葡萄孢發酵液中的除草活性組分進行純化和結構鑒定,得到有望開發為生物除草劑前體的10-順-二氫化灰霉二醛。
2 與致病性相關的胞外酶
灰葡萄孢侵染植物組織并導致植物死亡的過程一般從孢子開始,分生孢子成熟后脫落,借氣流、雨水傳播附著在植物表面,再萌發出芽管形成菌絲。灰葡萄孢突破植物體表面屏障入侵細胞,再將植物組織降解為可供真菌利用的能量,這一整個過程都要依賴胞外酶的協同作用[20]。
2.1 與破壞宿主表面屏障相關的酶
植物表面都有一層起保護作用的表皮,這層表皮由角質和脂肪酸構成,此外許多植物表面還有一層由脂肪醇組成的疏水蠟質層。角質的主要成分是單、雙及三羥基脂肪酸,這些羥基脂肪酸通過酯鍵相互連接。表皮細胞壁的角質化以及角質層和蠟質層的存在使整個植物表面形成堅固的疏水層抵御病原菌的侵害。
灰葡萄孢感染植物從植物表面開始進行,即使宿主表面不存在傷口,灰葡萄孢的孢子也能由植物表皮直接入侵宿主引起發病,但有創口或死亡的植物組織更容易感染灰霉病。這是因為灰葡萄孢入侵宿主表面時能分泌減小宿主表面疏水性并分解蠟質層的蛋白及代謝物,為其它致病因子提供了入口。這些分泌物起到表面活化劑的作用,也就是減小了宿主的表面張力,在此過程中起作用的胞外酶包括了角質酶、絲氨酸酯酶、脂酶和其它非特異性酯酶。表1涵蓋了一些協助灰葡萄孢破壞宿主表面屏障的酶和這些酶起到的作用[21-25]。

表1 與灰葡萄孢破壞宿主表面相關的胞外酶及功能
2.2 與利用植物能量相關的酶
灰葡萄孢的孢子穿過植物表面保護層后萌發為菌絲,分泌大量可分解植物組織的胞外酶,這些酶將植物細胞中的多糖降解為可供真菌利用的生物量。植物細胞壁由多種多糖組成,其中主要的組成是纖維素和半纖維素,胞間層則含有大量的果膠質,這個階段灰葡萄孢產生的與致病性相關的酶主要是纖維素酶、半纖維素酶等細胞壁降解酶和果膠酶。
2.2.1 纖維素酶和半纖維素酶
灰葡萄孢在纖維素的誘導下分泌纖維素水解酶,將植物細胞壁中的纖維素分解為纖維二糖和葡萄糖。體外實驗證明,當培養基中加入果膠時纖維素酶的產量會增加,而葡萄糖和纖維二糖則會抑制纖維素酶分泌[26]。灰葡萄孢分泌的纖維素水解酶包括 β-葡萄糖苷酶,1,4-β-D-葡聚糖內切酶和1,4-β-D-葡聚糖外切酶,其中β-葡萄糖苷酶既有胞內酶也有胞外酶,但在灰葡萄孢侵染植物的過程中,只有胞外酶作用于植物細胞,胞內β-葡萄糖苷酶與灰葡萄孢的致病性沒有關聯[27]。灰葡萄孢的孢子無論是在萌發前還是萌發后都沒有檢測到纖維素酶的活性,說明纖維素酶只在灰葡萄孢進入宿主后起作用。
植物細胞壁中的半纖維素是由木聚糖和阿拉伯糖組成的,有報道稱灰葡萄孢侵染植物后分泌出3種木聚糖酶和1種阿拉伯糖酶[28],其它關于灰葡萄孢分泌半纖維素酶的研究幾乎沒有。
2.2.2 果膠酶
植物的細胞壁和胞間層中都含有大量的果膠質,在灰葡萄孢侵染植物的過程中會產生多種可以降解果膠的酶,它們在降解果膠的過程中起著不同的作用,協助灰葡萄孢分解果膠的酶及其功能如表2 所示[29-33]。

表2 灰葡萄孢分泌的果膠酶及功能
Movahedi等在經灰葡萄孢侵染后壞死的植物細胞中發現了多外聚半乳糖醛酸酶,果膠裂合酶和果膠酸裂解酶。果膠酸裂解酶在孢子中就存在,但在灰葡萄孢侵染宿主的起始階段不發揮作用,而多外聚半乳糖醛酸酶在灰葡萄孢侵染宿主的早期就開始發揮作用,黃瓜葉片接種灰葡萄孢9 h后就能檢測到多外聚半乳糖醛酸酶。此外,在大豆下胚軸和南瓜果實中都檢測到果膠裂合酶和果膠酸裂解酶,而蘋果組織中卻沒有檢測它們的酶活。體外實驗發現這兩種酶在酸性pH下會失活,所以這2種酶在酸性的蘋果組織內失活,此外果膠裂合酶還需要Ca2+離子起協同作用[31-33]。
3 展望
灰葡萄孢引起的灰霉病是世界性的重要病害,給農業生產帶來了極大危害。幾十年來大面積使用化學藥劑防治灰霉病帶來了嚴重的后果,抗藥性的菌株大量出現,土地污染和水污染都日趨嚴重。發展靶標性的抗灰霉病藥物成為目前研究的熱點,靶標性的抗菌劑只作用于目標微生物,而不作用于其它生物,對于保持生態系統的平衡十分有利。
設計靶標性的抗菌劑需要了解病原真菌代謝產物的合成途徑以及對植物的作用機理,灰葡萄孢的代謝產物包括酶類、多糖、植物激素、毒素和脂肪酸酯類等,其中酶類、毒素和植物激素是重要的致病因子。致病因子作用于植物是一個復雜的過程,近年來更多的研究由致病因子的作用機制轉向編碼相關致病因子的基因與合成途徑,對這些基因及調控機理的研究必將極大地促進新型抗灰霉病藥物的研制和發展。
[1]Garibaldi A.First report of botrytis blight caused by Botrytis cinerea on Chamelaucium uncinatum in Italy [J]. Plant Disease,2009,93(9):968-968.
[2]Zhang M,Tsukiboshi T,Okabe I.First report of Botrytis cinerea causing leaf spot of Aquilegia vulgaris in Japan [J].Plant Disease,2009,93(4):425-425.
[3]Oliveira M,Guerner M J,Mesquita M M,et al.Important phytopathogenic airborne fungal spores in a rural area:incidence of Botrytis Cinerea and Oidium Spp [J].Annals of Agricultural and Environmental Medicine,2009,16(2):197-204.
[4]Long C A,Yuan G.Kloeckera apiculata strain(34-9)to control Botrytiscinereaduringthepre- andpostharvest handling of strawberries[J].Annals of Microbiology,2009,59(1):77-81.
[5]Morandi M A B,Mattos L P V,Santos E R,et al.Influence of application time on the establishment,survival,and ability of Clonostachys rosea to suppress Botrytis cinerea sporulation on rose debris[J].Crop Protection,2008,27(1):77-83.
[6]Chen S C, Liu A R, Wang F H, etal. Combined overexpression of chitinase and defensin genesin transgenic tomato enhances resistance to Botrytis cinerea [J].African Journal of Biotechnology,2009,8(20):5182-5188.
[7]Jeon E H,Chung E S,Lee H Y,et al.Ectopic expression of wild rice OgGRP gene encoding a glycine rich cell wall protein confers resistance to Botrytis cinerea pathogen on Arabidopsis[J].Plant Pathology Journal,2009,25(2):193 -198.
[8]Mercier J,Kong M,Cook F.Prevalence of fungicide resistance in Botrytis cinerea isolates from strawberry fields in California[J].Phytopathology,2009,99(6):84-84.
[9]Rosslenbroich H J,Stuebler D.Botrytis cinerea - history of chemical control and novel fungicides for its management[J].Crop Protection,2000,19(8-10):557-561.
[10]Drobya S,Wisniewski M,Macarisinb D,et al.Twenty years of postharvest biocontrol research:Is it time for a new paradigm[J].Postharvest Biology and Technology,2009,52:137-145.
[11]Culter H G,Jacyno J M,Harwood J S,et al.Botcinolide:A Biologically active natural product from Botrytis cinerea [J].Bioscience,Biotechnology and Biochemist,1993,57(11):1980-1982.
[12]Jacyno J M,Harwood J S,Culter H G,et al.Structure and solution-state conformation of botcinolide,a new biologically active metabolite from the fungus Botrytis cinerea [J].Tetrahedron,1994,50(40):11585-11592.
[13]Rebordinos L,Cantoral J M,Prieto M V,et al.The phytotoxic activity of some metabolites of Botrytis cinerea [J].Phytochemistry,1996,42(2):383-387.
[14]Deighton N, MuckenschnabelI, ColmenaresA J, etal.Botrydial is produced in plant tissues infected by Botrytis cinerea[J].Phytochemistry,2001,57(5):689-692.
[15]Colmenares A J,Aleu J,Duran-Patron R,et al.The putative role ofbotrydialand related metabolites in the infection mechanism of Botrytis cinerea[J]. Journal of Chemical Ecology,2002,28(5):997-1005.
[16]Patron R D,Galan R H,Rebordinos L G,et al.Structure-Activity relationships of new phytotoxie metabolites with the botryane skeleton from Botrytis cinerea [J]. Tetrahedron,1999,55:2389-2400.
[17]Pinedo C,Wang C M,Pradier J M,et al.Sesquiterpene synthase from the botrydial biosynthetic gene cluster of the phytopathogen Botrytis cinerea[J ].Acs Chemical Biology,2008,3(12):791-801.
[18]Wang C M,Hopson R,Lin X,et al.Biosynthesis of the sesquiterpene botrydialin Botrytiscinerea mechanism and stereochemistry of the enzymatic formation of presilphiperfolan-8β - ol[J].Journal of the American Chemical Society,2009,131(24):8360-8361.
[19]鄭蒙,徐擴,董金皋.灰葡萄孢BC7-3菌株除草活性組分的純化與結構鑒定 [J].微生物學報,2008,48(10):1362-1366.
[20]Boulard T,Chave M,Fatnassi H,et al.Botrytis cinerea spore balance of a greenhouse rose crop[J].Agricultural and Forest Meteorology,2008,148(3):504-511.
[21]Commenil P, BelingheriL, Bauw G, etal. Molecular characterization of a lipase induced in Botrytis cinerea by components of grape berry cuticle [J]. Physiological and Molecular Plant Pathology,1999,55(1):37-43.
[22]Reis H, Pfiffi S, Hahn M. Molecular and functional characterization of a secreted lipase from Botrytis cinerea[J].Molecular Plant Pathology,2005,6(3):257-267.
[23]Gindro K,Pezet R.Purification and characterization of a 40.8-kDa cutinase in ungerminated conidia of Botrytis cinerea Pers.Fr[J].Fems Microbiology Letters,1999,171(2):239 -243.
[24]Vankan J A L,Vantklooster J W,Wagemakers C A M,et al.Cutinase A of Botrytis cinerea is expressed,but not essential,during penetration of gerbera and tomato[J].Molecular Plant-Microbe Interactions,1997,10(1):30-38.
[25]Vankan J.License to kill:The role of phytotoxic proteins and pectinases in pathogenicity of Botrytis cinerea [J].Phytopathology,2005,95(6):136.
[26]Touzan A,DonecheB. Production and propertiesofthe cellulase complex of Botrytis cinerea[J].Canadian Journal of Botany-Revue Canadienne De Botanique,1996,74(3):486-491.
[27]Sasaki I,Nagayama,H. Beta - Glucosidase from Botrytis cinerea its relation to the pathogenicity of this fungus[J].Bioscience Biotechnology and Biochemistry,1994,58(4):616-620.
[28]Urbanek H,Zalewska S J.Multiplicity of cell wall degrading glycosidic hydrolases produced by apple infecting Botrytis cinerea[J].Phytopathologische Zeitschrift,1984,110(3):261 -271.
[29]Kars I,McCalman M, WagemakersL, etal. Functional analysis of Botrytis cinerea pectin methylesterase genes by PCR-based targeted mutagenesis:Bcpme1 and Bcpme 2 are dispensable for virulence of strain B05.10[J].Molecular Plant Pathology,2005,6(6):641-652.
[30]Poinssot B, Vandelle E, Bentejac M, et al. The endopolygalacturonase 1 from Botrytis cinerea activates grapevine defense reactions unrelated to its enzymatic activity [J].Molecular Plant Microbe Interactions,2003,6(16):553-564.
[31]Kapat A,Zimand G,Elad Y.Biosynthesis of pathogenicity hydrolytic enzymes by Botrytis cinerea during infection of bean leaves and in vitro[J].Mycological Research,1998,102:1017-1024
[32]Rha E, Park H J, Kim M O, etal. Expression of exopolygalacturonases in Botrytis cinerea [J]. FEMS Microbiology Letters,2001,201:105-109.
[33]Movahedi S,Heale J B.The Roles of aspartic proteinase and endo-pectin lyase enzymes in the primary stages of Infection and pathogenesis of various host tissues by different isolates of Botrytis cinerea Pers Ex Pers[J].Physiological and Molecular Plant Pathology,1990,36(4):303-324.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
